Ovarian cancer is an aggressive gynaecological cancer with extremely poor prognosis, due to late diagnosis as well as the development of chemoresistance after first-line therapy.
- ovarian cancer
- cancer stem cells
- immune surveillance
1. Introduction
In 1992, ovarian cancer was termed ‘the most lethal gynaecologic malignancy’ [1], with the overall five-year survival rate reported at 30%. Although the past three decades have seen a significant improvement in diagnostic advances, therapeutic strategies and overall care in ovarian cancer, prognosis continues to remain poor. The current five-year survival rate of 48.6% is the lowest among all gynaecological cancers [2], meriting the dismal title of ovarian cancer being the deadliest gynaecological cancer. Over 90% of all ovarian cancers are of epithelial origin and can be broadly divided further into Type I (including low- grade serous, endometrioid, clear-cell or mucinous carcinomas) and Type II (including high-grade serous or undifferentiated carcinomas).
Population-based cancer incidence and mortality data is compiled by various organisations across the world. For Europe, the European Cancer Information System estimates an age standardised incidence rate of ovarian cancer at 16.1 per 100,000 and an associated mortality rate of 10.4 per 100,000 (
) [3]. This high mortality-to-incidence ratio is attributable to a combination of late detection and resistance to therapy. The improbability of early diagnosis is a direct consequence of the lack of specific symptoms during the early stages of the disease, as well as the absence of reliable screening strategies. Owing to the success of cervical and breast cancer screening, as well as the rather modest increase in survival from improved treatment, there have been fervent efforts to boost ovarian cancer survival via screening using CA125, an epitope of MUC16, a large glycoprotein marker. However, the accuracy of this biomarker is still questionable, although more effective screening strategies with CA125 are being developed [4]. As outlined before, therapeutic advances have led to only a small increase in ovarian cancer survival rate over the years. Standard treatment for ovarian cancer is cytoreductive surgery along with combination taxane–platinum-based chemotherapy. More recently, the two most promising novel therapeutic approaches are using monoclonal antibodies such as bevacizumab, targeting tumour microenvironmental pathways such as angiogenesis, and inhibitors of the poly (ADP-ribose) polymerase (PARP) enzyme which is involved in critical cellular functions such as DNA repair. Both have been approved by the FDA and show promising outcomes as combinatorial and maintenance drugs in ovarian cancer [5].
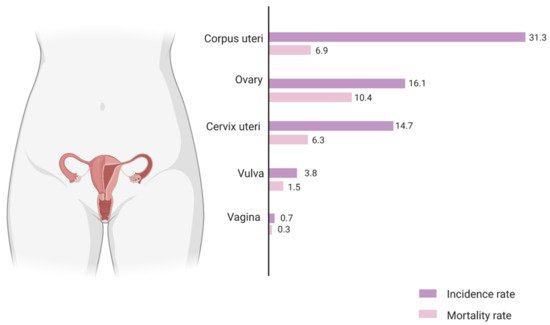
Figure 1.
Although first-line therapy has an initial remission rate of 70–80%, the majority of patients relapse, develop chemoresistance and proceed to respond only very modestly to second-line chemotherapy. The high recurrence rate and chemoresistance associated with ovarian cancer is thought to be due to intra-tumoral heterogeneity, microenvironmental interactions as well as the presence of dynamic cancer stem cell sub-populations. There are three main models proposed to explain the heterogeneity of intra-tumoral cell populations. The two conventional models are the clonal evolution or stochastic model and the stem cell or hierarchical model. It is now understood that the two ideas are not mutually exclusive, and a third model termed the plasticity model conceptualises a more dynamic, flexible understanding of the tumoral niche (
). Stem cell-like subpopulations existing in the tumoral hemisphere in solid tumours such as ovarian cancer have been found to dynamically interact with the immediate cellular microenvironment so as to induce tumorigenesis, survival and metastases as well as self-renewal leading to an intrinsically generated and maintained tumour niche capable of immunosuppression and therapeutic evasion. Hence, it is vital to study these interactions and devise methods that effectively target these stem cell niches to make substantial strides in the therapeutic targeting and management of aggressive ovarian tumours. This review aims to summarize the current understanding of the ovarian cancer stem cell niche and its interactions with the host immune system and to highlight implications for the development of novel ovarian cancer therapies.
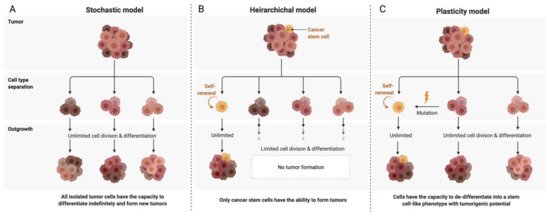
Figure 2.
A
B
C
2. Ovarian Cancer Stem Cells (OCSCs): Signaling Pathways and Markers
Like many solid tumours, ovarian cancer has been shown to reflect significant tumoral phenotypic diversity [6]. Key evidence suggests that the high relapse rate inevitably seen in ovarian cancer is linked to chemoresistant stem cell-like subpopulations which persist through therapy and have tumorigenic properties [7]. In 2013, Virant-Klun et al., first discovered very small embryonic-like stem cells identifying stage-specific embryonic antigen-4- (SSEA-4; a marker of human embryonic stem cells) positive cells from cultures of human ovarian cancers and validated their discovery in women with borderline ovarian cancer (a less aggressive form of epithelial ovarian cancer) versus healthy women. The cells from the test group were proliferative and formed tumour-like structures in vitro as well as in vivo [8][9].
2.1. Signaling Pathways
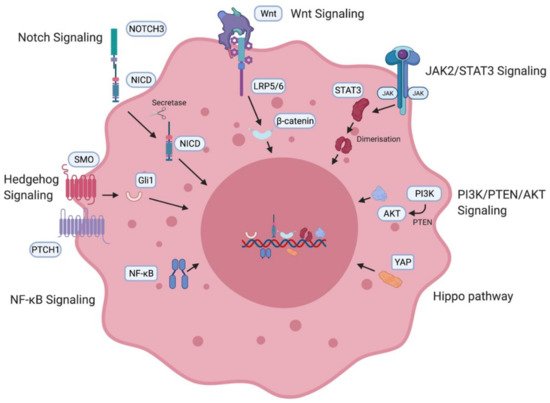
Figure 3. Ovarian cancer stem cell (OCSC)-associated signaling pathways. OCSC signaling pathways involved in the generation and maintenance of OCSCs including the Notch pathway [10][11], Wnt signaling pathway [10][11], JAK2/STAT3 pathway [12][13][14], PI3K/PTEN/AKT pathway [15], Hippo pathway [16], NF-κB [17][18] and the Hedgehog pathway [19]. NICD—intracellular domain of Notch protein; LRP—low-density lipoprotein-related protein; JAK—Janus kinase, STAT—signal transducer and activator of transcription proteins; PI3K—phosphatidylinositol 3-kinase, PTEN—phosphatidylinositol 3,4,5—triphosphate 3-phosphatase, AKT/PKB—protein kinase B; YAP—Yes-associated protein; NF-κB—nuclear factor kappa B.
2.2. Cancer Stem Cell Markers
Table 1.
| Marker | Characteristic | Function in Ovarian Cancer | Evidence |
|---|---|---|---|
| CD133 | Transmembrane glycoprotein | Identified by several groups to be expressed in tumour-initiating cells; promotes adhesion to metastatic cancer niche. | Ferrandina et al. [32], Roy et al. [33] |
| ALDH | Aldehyde dehydrogenase enzyme | Correlates with tumourigenicity and spheroid formation; increased expression significantly associated with poor outcomes in patients with serous ovarian cancer. | Ma et al. [34], Ishiguro et al. [35], Deng et al. [36] |
| CD44 | Transmembrane glycoprotein | Positively associated with ovarian cancer migration and metastatic spread; high expression correlates to recurrence and drug resistance. | Bourguignon et al. [37], Carpenter et al. [38], Sacks et al. [39] |
| CD24 | Glycophosphatidylinositol-anchored membrane glycoprotein | Positive marker; cell lines and tumour samples displayed stemness genes, tumourigenicity, spheroid formation. | Burgos-Ojeda, D. et al. [40], Gao, M.Q. et al. [41] |
| CD117 | Receptor tyrosine kinase | Surface marker binding to stem cell factor; consistently formed tumours in mice models | Mazzoldi et al. [42], Luo et al. [43] |
3. The Ovarian Cancer Stem Cell Niche
3.1. Epithelial–Mesenchymal Transition
The process by which an epithelial phenotype undergoes transition first by increasing in dimension and subsequently by acquiring a mesenchymal phenotype is called EMT [44]. One of the very first studies identifying stem-cell like subpopulations in the ovarian epithelium by Virant-Klun et al. found strong evidence that the stem cell niche induced EMT [8]. This transition is a dynamic process occurring in conjunction with persisting surrounding epithelial cells, as well as a wide spectrum of stromal cells (fibroblasts, immune cells) and endothelial cells, and enabling invasive and migratory properties within cancer cell populations [45]. Specific transcription factors are associated with the transitional process and can be mainly categorized into three families—TWIST, Snail and ZEB [46]. They suppress epithelial state-inducing genes like E-cadherin and stimulate mesenchymal state-inducing genes like N-cadherin [46]. These transcription factors have also been associated with expression of stemness-enhancing genes [47][48]. In the ovarian cancer stem cell niche, TGF-β signaling plays a significant role in promoting EMT via regulation of tissue transglutaminase 2 (TTGM2) [49]. A dynamic EMT state leads to increased stemness and enables chemoresistance. OCSCs exist in an intermediate epithelial–mesenchymal state, expressing both kinds of markers and equipping them with unique potential for adhesion and migration, respectively [50]. This dials into the plasticity model for cancer stem cells by proving that stemness is a dynamic interconvertible state [51].
3.2. Hypoxia
While hypoxia has been implicated as a driver in the maintenance of most cancer stem cell niches, it is of particular interest in ovarian cancer due to the presence of ascites which serve as metastasis hotspots for invasive spheroid formation. Ascites contain half the soluble oxygen as blood [52]. This hypoxic condition stimulates the hypoxia-inducible transcription factor-1 alpha (HIF-1α) to initiate hypoxia-responsive downstream signaling of various target genes which allows cells to adapt to environmental insults. Hypoxia drives stemness [53] and induces chemoresistance potential by maintaining OCSCs in a quiescent state, shielding them from drugs intended to target proliferative cells [54]. HIF-1,2 are involved in stimulating fibroblasts to secrete CXCL12, which is believed to initiate the cancerous phenotype in ovarian cancer. [55] These cells are also able to respond to stress [56], whilst also being invasive and migratory, and can promote increased angiogenic potential [52]. Reactive oxygen species (ROS) are produced by cancer cells and can stimulate oncogenes and facilitate new mutations. A recent study verified that ROS levels were eight times higher in tumours from 34 Stage III/IV HGSOC patients than in non-cancerous ovaries [57]. Elevated level of ROS in cancer stem cells has been found to promote cancer metastasis by inducing EMT via the TGF-β pathway [58][59].
3.3. Neovascularisation and Angiogenesis
3.4. Inflammation
4. Ovarian Cancer Stem Cell Niche and Inflammatory Networks
Although there has been a long apparent association between inflammation and cancer, it was only introduced as one of the ‘Hallmarks of Cancer’ in Hanahan and Weinberg’s second, revised magnum opus [64]. Chronic inflammation has been established as a cause of several cancers [65], and the phenotypes, processes and pathways associated with various immune cells and interactions contribute to the dynamic maintenance of the tumours at a microenvironmental level [66]. These correlations have been verified in vitro, in zebrafish [67] and mouse models [68] as well as in patient prognostic data [69]. Specifically, cancer stem cells can use immune surveillance evasion to enhance their survival and invasive properties. Growing evidence suggests that cancer stem cells are able to not only circumvent key immune checkpoints, but also manipulate inflammatory networks to promote self-sustenance, tumorigenesis and cellular invasion [68]. Ovarian cancer, in particular, is a classic example of a stem cell-driven cancer. It metastasises via a trans-coelomic route spreading to the peritoneal organs in the form of persistent spherical multicellular aggregates. The primary tumour is capable of metastasising very early due to the ability to form spheroids from ascites, which proliferate and persist even in the absence of organ adhesion, and displays key stemness attributes [70]. These cells invade the extracellular matrix where they interact with the cellular microenvironment consisting broadly of immune (cytokines, macrophages, lymphocytes) and non-immune (adipocytes, fibroblasts, endothelial) cell components (
).
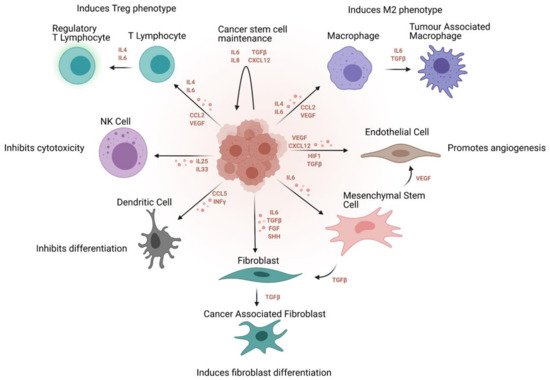
Figure 4.
4.1. Cytokine Signaling
Not only have cytokines been identified in ovarian cancer patient ascites and cysts [71], they have also been found in the tumour stroma and epithelium [72]. This indicates that active cytokine-mediated signaling is part of the microenvironmental interactions in the ovarian tumour niche. Non-tumoral cells like adipocytes in the omentum and endothelial cells of the vasculature also trigger the release of cytokine signaling. Adipocyte-mediated cytokine signaling induces a change in lipid metabolism and allows cancer cells to use fatty acids as fuel for proliferation [73]. In ovarian cancer, adipocytes express IL-6, increasing the expression of BCLxl that provides the ability to cancer stem cells to become resistant to drug therapy [74]. Endothelial cells on the other hand, enhance inflammation and angiogenetic potential along with cell migration in the tumoral niche via the release of TNF-α, VEGF and interleukins (IL) [75]. IL-17 was one of the first cytokines identified in the ovarian cancer niche which was found to promote self-renewal of OCSCs [76][77]. Upon further investigation, it was found that OCSCs expressed the IL-17 receptor which promotes self-sustenance and growth via the NF-κB and MAPK pathways [76]. The NF-κB pathway has also been implicated via the release of IL-23 [78] and CCL5 [79] by OCSCs, which further enriches the angiogenic potential of tumour cells within the niche.
4.2. Tumour-Associated Macrophages (TAMs)
TAMs constitute the highest percentage of immune cells in the tumour niche. JAK2/STAT3 activation within TAMs promotes increased tumorigenicity, chemoresistance and stemness within tumours [80]. Subsequently, anti-tumour CD8+ responses from chemotherapeutic targeting are blocked by the cancer stem cell niche and the polarisation of the TAMs towards an anti-inflammatory M2 phenotype [80]. M2 macrophages in general have been seen to have a notable positive impact on the progression of tumours in different cancers [81]. In particular, among patients with high-grade ovarian cancer, M1 macrophages were significantly associated with better outcomes, while the M2 phenotype was associated with worse outcomes [82]. A co-culture study proved that OCSCs are capable of polarising the macrophage phenotype towards an M2 state via COX-2 overexpression and cytokine production, involving the JAK2/STAT3 pathway [83]. Furthermore, the M2 phenotype stimulates cancer stem cell self-renewal and growth via various signaling pathways e.g., EGF, TGF-β, IL-6 and IL-10, that lead to STAT3 activation [84]. NF-κB signaling pathways are activated, causing subsequent recruitment of M2 macrophages and also contributing to supplementary production of cytokines, and hence feedback into the self-sustaining cycle of the cancer stem cell niche [84]. Furthermore, an immunosuppressive microenvironment may originate as an outcome of the responses of CD4+ Treg T cells that are stimulated by M2 macrophages [85]. Additionally, macrophages make the tumour microenvironment amicable for cancer stem cell seeding as well as migration [86][87].
4.3. Tumour-Infiltrating Lymphocytes (TILs)
4.4. Natural Killer Cells (NK)
Similar to T cells, NK cells are capable of acquiring memory functional phenotypes once target cells are encountered, thereby bridging the gap between adaptive and innate immune systems [93]. Cancer stem cells can be killed in a major histocompatibility complex (MHC)-unrestricted manner by NK cells [94] via the release of TNF family members [95]. Immunoglobulin Fc, inflammatory cytokines and endogenous ligands activate these NK cells [96]. A range of activating and inhibitory receptors modulate NK cell function. These receptors sense changes, such as loss of MHC in tumour cells, and subsequently allow NK cells to respond accordingly [93]. It was found that OCSCs downregulate NK cell function. Ascites of ovarian cancer patients have been found to have increased levels of NK cells. However, due to the immunosuppressive effects of the ovarian cancer stem cell niche and the dysregulation of natural and cell-mediated cytotoxicity, these cells are functionally impaired [97].