Vitamin D is a potent immunonutrient that through its main metabolite calcitriol, regulates the immunomodulation of macrophages, dendritic cells, T and B lymphocytes, which express the vitamin D receptor (VDR), and they produce and respond to calcitriol. Genetic association studies have shown that up to 65% of vitamin D serum variance may be explained due to genetic background.
- vitamin D
- polymorphism
- calcidiol
- calcitriol
- VDBP
- GC
- CYP2R1
- CYP27B1
- VDR
1. Introduction
The etiology and progression of autoimmune diseases (AIDs) are multifactorial and complex[1]. Genetic and environmental factors such as nutrients have been proposed to partially explain the pathophysiology progression of autoimmunity[2]. Notably, vitamin D regulates the growth and differentiation of various cells of the immune system such as macrophages, dendritic cells, T cells, and B cells, which are able to express the vitamin D receptor (VDR), produce and respond to the active form of vitamin D, calcitriol (1α,25(OH)
2
3)[3].
In autoimmune diseases, epidemiological studies have reported a high prevalence of vitamin D deficiency by the quantification of calcidiol; this deficiency has been associated with worse disease clinical activity and progression of systemic lupus erythematosus (SLE), rheumatoid arthritis (RA), and multiple sclerosis (MS) diseases[4]. Likewise, several clinical trials and murine studies using cholecalciferol supplementation have described vitamin D′s immunomodulatory properties in autoimmune diseases[5]. Lower calcidiol serum levels are a strong predictor for worse MS clinical manifestations [6] as well as RA patients show lower calcidiol levels than control subjects (CS), and they have shown a negative association of calcidiol serum levels with clinical disease activity[7]. Regarding SLE patients, a higher prevalence of calcidiol serum deficiency than the general population has also been reported[8], and the lower serum calcidiol in SLE patients is associated with high clinical disease activity[9].
Calcidiol serum deficiency in patients with autoimmune diseases and the general population could be attributed to several factors, including lack of exposure to sunlight, skin pigmentation, sunscreen use, nutrient intake deficiencies, age, use of glucocorticoids, and the genetic background of the populations[10]. The potential roles of 35 genes that could modulate the vitamin D serum levels status have been reported in previous studies, highlighting that multiple single nucleotide polymorphisms (SNPs) in these genes are associated with lower calcidiol serum levels[11], such as the SNPs described in the vitamin D binding protein (VDBP; rs2282679
GC
CYP2R1
CYP27B1
FokI
Bsml
Apal
Taql
VDR
GC
CYP2R1
CYP27B1
VDR
2. Vitamin D Status and Genetic Evidence in the Populations
Regarding vitamin D deficiency, several factors have been described, mainly the lack of exposure to sunlight, latitude, the season of the year, skin pigmentation, and use of sunscreen; other factors involved in vitamin D deficiency are diet, age, pharmacotherapy administered (antiepileptic and glucocorticoids), and particularly, several studies have described that genetic differences between individuals and populations such as genetic polymorphisms could influence the vitamin D deficiencies presented in all populations around the world[10].
GC
CYP2R1
CYP27B1
VDR genes are associated with lower calcidiol serum levels[11]. Moreover, vitamin D serum deficiencies are present in a high frequency in a healthy population, which could be related to the SNPs’ presence in these genes that may modify the response to supplementation of vitamin D in health and disease. A study conducted in healthy Iranian adolescents described the differential effect of the
CYP2R1
p = 0.002)[12]. This is evidence of the role of polymorphisms in genes related to vitamin D metabolism in the variation of the response to vitamin D supplementation, even in healthy conditions.
3. Polymorphisms in the Main Key Genes Related to Vitamin D Metabolism
The skin produces around 80% of vitamin D in the form of cholecalciferol when exposed to ultraviolet B (UVB) light at a wavelength of 290–320 nm, and the remaining 20% is obtained from the diet as ergocalciferol from mushrooms or as cholecalciferol from fortified dairy products, fish, and eggs, mainly [13].
GC
Figure 1a). The
GC (rs2282679) SNP was been associated with modulation of vitamin D serum levels in several populations[14][15][16]. In these studies, the main vitamin D circulating metabolite in blood, calcidiol, was used to evaluate vitamin D deficiencies [17].
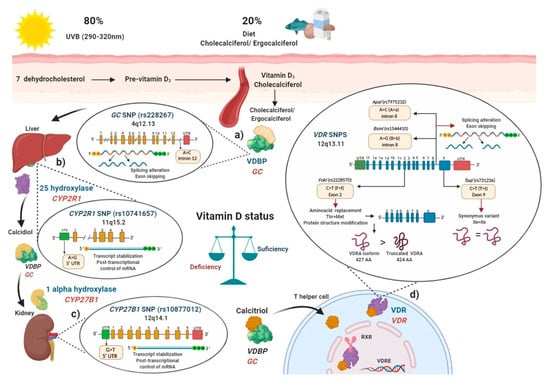
Figure 12.
a
GC
GC
b
CYP2R1
CYP2R1
c
CYP27B1
CYP27B1
d
VDR
IL-17A
FOXP3.
VDR
FokI
FokI
BsmI
VDR
VDR
ApaI
VDR
VDR
TaqI
VDR, presents a change of C > T (also called T > t) and generates a synonymous change of the isoleucine amino acid in the coding sequence, therefore it does not change the encoded protein, but it could influence the stability of the mRNA. All these SNPs are related to modulating de vitamin D serum status in health and disease. Ile: isoleucine; Thr: threonine; Met: methionine VDBP: vitamin D binding protein; VDR: vitamin D receptor; RXR: retinoid X receptor; VDRE: vitamin D response elements; UTR: untranslated region; THEM4: thioesterase superfamily member 4; Th: T helper lymphocyte; VDRA: wild type full-length VDR A isoform.
CYP2R1
Figure 1b), the (rs10741657) SNP described in
CYP2R1 had also been related to modulate calcidiol serum status in several studies[12][14][15][18]. Then, after the generation of calcidiol in the liver, it binds again to VDBP and subsequently interacts with the enzyme 1-α hydroxylase, encoded by the
CYP27B1
Figure 1c)[13]. Polymorphisms in
CYP27B1, particularly the rs10877012 SNP, was associated with lower calcidiol serum levels[19].
VDR
After the calcitriol translocation to the nucleus, this complex forms a heterodimer with the retinoid x receptor (RXR), and thus regulates vitamin D target genes through its binding to specific DNA sequences called vitamin D response elements (VDREs)[20][21]. Since VDR is expressed in different immune cells, such as neutrophils, macrophages, dendritic cells (CDs), T and B lymphocytes, calcitriol may regulate the immune system[8].
Calcitriol can be produced by monocytes and macrophages and generate a shift from pro-inflammatory to tolerogenic immune status[22]. Calcitriol promotes M1 phenotype switching to M2 via the nVDR-PPARγ pathway and via the upregulation of the expression of IL-10[23]. Particularly, calcitriol promotes a shift from Th1 and Th17 to Th2 immune profile via suppression of expression of cytokines of the Th1 (IL-2, IFN-γ, TNF-α) and Th17 (IL-17, IL-21) profiles, and induction of the expression of cytokines of Th2 profile (IL-4, IL-5, IL-9, IL-13), in order to limit inflammatory processes and autoimmune reactions [8][22].
VDR
FokI
BsmI
ApaI
TaqI
Figure 1d). Therefore, SNPs′ presence in key enzymes related to vitamin D metabolism may modulate calcidiol levels and calcitriol function; they may also modify the disease activity in MS, RA, and SLE through the vitamin D deficiency, and contribute to genetic susceptibility to autoimmunity. In the following sections, we will describe how polymorphisms in these genes can modulate their expressions.
3.1. Vitamin D Binding Protein (VDBP) (SNP rs2282679
3.1. Vitamin D Binding Protein (VDBP) (SNP rs2282679
GC)
Vitamin D binding protein (VDBP) is the main carrier protein for vitamin D, which binds 85 to 90% of the total circulating calcidiol [24]. This protein is encoded by the
)
GC gene located in chromosome 4, position 13.3 (4q13.3), and has 55,136 bp size [25]. The
GC gene consists of 13 exons and the rs2282679 SNP was described located at the position 71,742, presenting an A ˃ C change (ancestral allele: A) in the intron 12, near to the actin III subdomain[26] and the endonuclease enzyme commonly used to identify this
GC
FokI
Flavobacterium okeanokoites) [27]. According to rs2282679 SNP intron location, its hypothetical functional effect may alter genetic mRNA expression due to modification of the splicing process (44). A complete GWAS showed that
GC (rs2282679) C allele was associated with lower serum calcidiol and VDBP levels in a study carried out in approximately 30,000 subjects with European ancestry included in 15 cohorts[28]. Additionally, various studies have shown the association of the
GC (rs2282679) SNP with genetic susceptibility and disease modulation in MS, RA, and SLE[29][30][31][32] (
Figure 1a).
3.2. Vitamin D 25-Hydroxylase (SNP rs10741657
3.2. Vitamin D 25-Hydroxylase (SNP rs10741657
CYP2R1)
The enzyme vitamin D 25-hydroxylase is a protein of around 500 amino acids with a molecular weight of 50–55 kilo-daltons (kDa). The liver is the main site where it is synthesized and performs its function of 25-hydroxylation enzyme activity, besides this enzyme activity was also described in the kidney and intestines[33], but cholecalciferol and ergocalciferol are metabolized mainly by the vitamin D 25-hydroxylase to calcidiol in the liver[34].
The enzyme vitamin D 25-hydroxylase is encoded by the
)
CYP2R1
CYP2R1 is positioned at 14,893,332 pb in the gene, it displays an A ˃ G change in the 5′ UTR region (ancestral allele: A)[18].
In order to recognize the presence of this SNP, the endonuclease restriction enzyme
MnlI
Moraxella nonliquefaciens) is commonly used[35]. According to the SNP location in 5′ UTR, the rs10741657 SNP may regulate gene expression by modifying translation initialization and transcript stabilization of mRNA, and therefore, modulate 25-hydroxylase expression and enzymatic activity rate[36].
A meta-analysis that included 16 articles with 52,417 participants showed that the
CYP2R1 (rs10741657) GG genotype shows trends of low calcidiol serum levels compared to the AA genotype in the Caucasian and Asian population[18] and when healthy persons are supplemented with cholecalciferol, carriers of the AA genotype showed a 2.5-fold increase in calcidiol levels compared to GG genotype carriers [12], which highlights the role of
CYP2R1
Figure 1b).
3.3. Vitamin D 1-α Hydroxylase (SNP rs10877012
3.3. Vitamin D 1-α Hydroxylase (SNP rs10877012
CYP27B1)
The enzyme vitamin D 1-α hydroxylase corresponds to a P450 protein of 507 amino acids of around 55 kDa[37]. This enzyme metabolizes calcidiol to calcitriol, the active form of vitamin D[34], and it is encoded by the
)
CYP27B1
CYP27B1 is located at the non-coding region -1260 and is characterized by a change of G ˃ T (ancestral allele: G)[38][39]. The restriction endonuclease enzyme
HinfI
Haemophilus influenza) is commonly used for its detection [27][35]. Due to its location in the 3′ UTR region, this SNP may alter transcript stabilization regulation and its localization in the cytoplasm [36].
A study carried out in 253 German patients with differentiated thyroid carcinoma showed that patients with the presence of the GG genotype (Referred as CC in this study by its position in the negative DNA strand) was associated with lower calcitriol serum levels than patients carrying the TT genotype (Referred as AA in this study by its position in negative DNA strand) (60 pmol/mL vs. 72 pmol/mL, respectively)[38].
In another study carried out in Caucasian German patients with gestational diabetes, the GG (CC) genotype was also associated with lower calcidiol levels[19]. Likewise, another study carried out in a healthy Caucasian British population demonstrated an association between the
CYP27B1 (rs10877012) G (C) allele and lower serum calcidiol levels[40].
Besides, in type 1 diabetes in Caucasian patients from Germany, those carrying the GG (CC) genotype had a reduced amount of mRNA from
CYP27B1
p = 0.0220)[19]. Therefore, the SNP rs10877012
CYP27B1
Figure 1c).
3.4. Polymorphisms in Vitamin D Receptor (VDR)
VDR
Functionally VDR is a transcription factor regulated by ligand binding and possibly by phosphorylation events[41]. It is a soluble 427 amino acid protein located mainly in the nucleus, cell cytoplasm, and cellular membrane, from where it translocates to the nucleus through the microtubule system after interaction with its ligand, calcitriol [42].
The most common is the VDRA isoform of 427 amino acids and 48 kDa, with a start site in exon 2. The second is a long VDRB1 isoform of 477 amino acids and 54 kDa, this isoform presents 50 amino acids more in the N-terminal domain by an ATG start site in the exon 1d, described in the human kidney as well as in intestinal and renal epithelial cell lines[42][43].
FokI in the exon 2[42].
VDR
FokI
BsmI
ApaI
TaqI (rs731236) [41][44]. These were related to modulating the vitamin D status independently (in alleles and risk genotypes) as well as in haplotypes and haplogenotypes, both in original articles and meta-analyses (
Figure 1d).
3.4.1. FokI (rs2228570) VDR SNP
FokI
VDR
FokI
(Flavobacterium okeanokoites) restriction enzyme in a restriction fragment length polymorphism test (RFLP) [45].
FokI (rs2228570) is located in exon 2 and is considered a non-synonymous polymorphism, because the change of C > T (ancestral allele T), also referred as F > f change, which generates a non-synonymous change of threonine to methionine and dictates two potential translation initiation sites[44].
The presence of the restriction site
FokI
FokI restriction enzyme), this C allele generates a new start codon (ATG) 9 bp after the common starting site, which translates to a shorter VDRA protein of 424 amino acids instead of the wild type full-length VDRA isoform of 427 amino acids[45].
FokI VDR SNP (rs2228570) was found to be functional, and the short 424 amino acid VDRA isoform is somewhat more active than the long VDRA isoform of 427 amino acid, in terms of its transactivation capacity as a transcription factor[46][47]. In the absence of the restriction site
FokI, the T allele (also called f allele), translation begins at the first original site at the exon 2, and the VDRA of 427 amino acids is expressed, which is 1.7-fold less active in its transactivation capacity, and presents less stability[48][49][50][51].
3.4.2. BsmI (rs1544410) and ApaI (rs7975232) VDR SNPs
BsmI
ApaI
VDR
BsmI
Bacillus stearothermophilus
ApaI
Acetobacter pasteurianus)
BsmI
VDR SNP, located in the intron 8, presents a change of A > G (also called B > b), and the ancestral allele is the G allele[49]. Regarding its functional effect, it could generate an alteration in the splice sites for mRNA transcription or a change in the intron regulatory elements of
VDR
ApaI (rs7975232) VDR SNP, also is located in the intron 8, presents a change of A > C, (also called A > a), and the ancestral allele is the C allele [21]. Both SNPs are located at the 3′ end of the
VDR
3.4.3. TaqI (rs731236) VDR SNP
3.4.3. TaqI (rs731236) VDR SNP
TaqI
VDR
TaqI
Thermus aquaticus
TaqI generates a synonym change of the coding sequence; therefore, it does not produce an amino acid change of the encoded protein, but it could influence the stability of the mRNA [49].
TaqI
VDR
ApaI
VDR SNP, its functional effect is the possible modification of one of the zinc fingers of the nuclear signaling heterodimer that binds to the VDREs located in the target genes[52].
VDR
BsmI
ApaI
TaqI
VDR SNPs were described with a strong LD[53][54], which infers that the alleles of these three polymorphisms could segregate into haplotypes from one generation of persons to another. In the case of
FokI
VDR
FokI
VDR SNPs [53][54]. However, because the genetic recombination points vary between populations,
FokI
BsmI
ApaI
TaqI