Drug resistance remains one of the main causes of poor outcome in cancer therapy. It is also becoming evident that drug resistance to both chemotherapy and to antibiotics is driven by more than one mechanism. So far, there are at least eight recognized mechanisms behind such resistance. In normal tissues, ATP-binding cassette (ABC) transporters protect the cells from the toxic effects of xenobiotics, whereas in tumor cells, they reduce the intracellular concentrations of anticancer drugs, which ultimately leads to the emergence of multidrug resistance (MDR). A deeper understanding of the structures and the biology of these proteins is central to current efforts to circumvent resistance to both chemotherapy, targeted therapy, and antibiotics.
- mass spectrometry multidrug efflux pumps
- (ABC) transporter proteins
- multidrug resistance
- ion mobility
1. Introduction
1.1. Developments Which Enhanced the Role of MS in Proteins Characterization
1.1.1. Hydrogen–Deuterium Exchange Mass Spectrometry(H/DX-MS)
In recent years, hydrogen–deuterium exchange combined with mass spectrometry (HDX-MS) [31][23] has emerged as a key player in studying the conformational dynamics and interaction of proteins in solution [16]. In this technique, a protein is diluted in a deuterated buffer, enabling H/D exchange of labile backbone amides. This process of isotopic exchange is strongly dependent on the protein secondary structure and solvent accessibility. In general, the exchange reaction can be quenched by dropping the pH and the temperature of the solution to 2.5 and 0 °C, respectively, conditions necessary to reduce the D-to-H back exchange. The quenched protein sample is then digested by an acid protease (e.g., pepsin), and the resulting peptides are separated by LC at low temperature and low pH, followed by MS and MS/MS analyses to determine the extent of deuterium incorporation (Figure 1).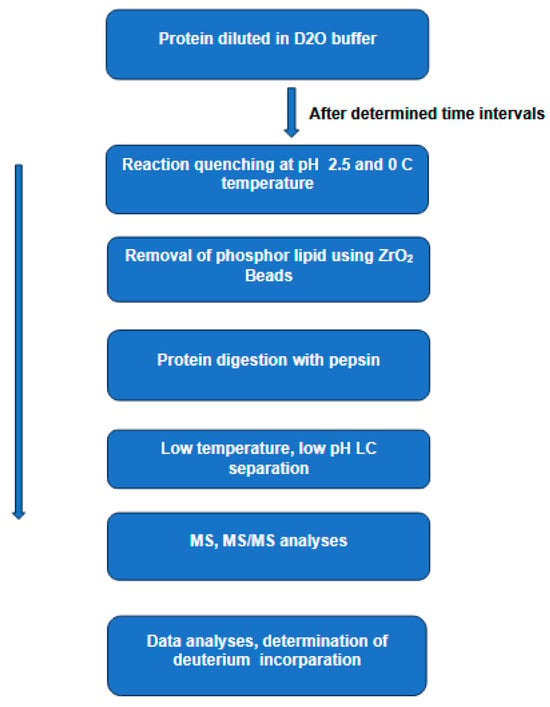
1.1.2. Ion Mobility–Mass Spectrometry
1.1.3. More Efficient Activation Methods of Macromolecules
For over 40 years, tandem mass spectrometry (MS/MS) has been performed by accelerating a mass selected precursor ion into a collision cell containing a neutral gas (Ar, N2) maintained at pressure approximately 10−4 to 10−2 torr and a collision energy of 10–100 eV. Such collision conditions are commonly used in triple quadrupoles [65][42], ion traps, and hybrid time-of-flight (Q-TOF). Over the last twenty years, softer and more efficient fragmentation methods have been introduced as alternatives or as complementary techniques to collision-induced dissociation (CID). These methods include electron transfer dissociation (ETD), electron capture dissociation (ECD), and photodissociation.1.1.4. Electron Capture Dissociation (ECD)
Over the last twenty years, electron capture (ECD) and electron transfer dissociation (ETD) have emerged as two of the most useful methods for peptide/protein fragmentation. The diffused use of both fragmentation methods coincided with unprecedented advances in mass spectrometry and liquid chromatography. In the ECD method, low-energy (less than 1 eV) electrons are captured by multiply charged protein/peptide positive ions generated by ESI [66][43]. This exothermic reaction induces dissociation through a H atom transfer to the backbone carbonyl group, resulting in a relatively soft cleavage of the Cα–N bond in a peptide backbone. Such low-energy interaction allows for much better detection and localization of labile PTMs as well as more sequencing information compared to classical CID. The predominant bonds cleaved in such a reaction are the backbone N–Cα, yielding c and z ions, with the exception of N-terminal proline cleavage, due to the proline’s cyclic nature [67,68][44][45] Since its introduction, the application of ECD has been limited to Fourier transform ion cyclotron resonance instruments (FT-ICR) [69][46]. The main reason for such a limitation is that efficient ECD requires a dense population of thermal electrons to interact with the precursor ions for a given time window. Creating such conditions in instruments, which use radio frequency to trap the precursor ions (e.g., ion traps and hybrid Q-TOF), proved to be technically challenging.1.1.5. Electron Transfer Dissociation (ETD)
Electron transfer dissociation (ETD) [73][47] is the direct result of interaction between multiply charged positive ions generated by electrospray ionization and a negatively charged radical reagent. The chemical properties of the radical reagent and the charge density of the precursor ion are the two parameters which influence the reaction. The two reaction pathways are deprotonation of the multiply charged ion and electron transfer from the radical reagent to the multiply charged ion (peptide or protein). The reactive radical anions are generated from polycyclic aromatic hydrocarbon molecules, such as anthracene or fluoranthene. The latter is considered as one of the more favorable ETD reagents. Electron transfer from the negatively charged reagent ions to multiply charged positive precursor ions induces dissociation of Cα–N bonds in a similar fashion to that observed in ECD. Both ETD and ECD are low-energy reactions, which explains their capability to detect and localize labile post-translational modifications, which are difficult to detect in collision-induced dissociation. That said, obtaining rich sequencing information by electron-based methods is closely related to the charge density of the precursor ion. Such influence manifests itself clearly in low charge density ions, where backbone cleavage takes place, but the resulting fragment ions are held together by non-covalent interactions present in the more compact structures, a process known as non-dissociative electron transfer [74][48]. One way to overcome such shortcoming is to chemically modify the investigated peptides [75,76][49][50] or to conduct combined ETD and CID, where the first method is used for high charge states, while the second is used for lower mass lower charge states [77,78][51][52].1.1.6. Photodissociation Methods
Currently there are two main photon-based methods for the activation and subsequent fragmentation of macromolecular ions. The first method uses infrared multiphoton dissociation (IRMPD), which was first demonstrated almost 50 years ago [81][53]. The authors used the wavelength 10.6 μm from a tunable continuous wave CO2 laser to irradiate a proton-bound dimer of di-Et ether [(C2H5)2O]2H+. The measurements were conducted in Fourier transform ion cyclotron resonance (FTICR). For many years the use of IRMPD remained limited to fundamental studies of small molecules; however, more recently, it is increasingly being applied to the analyses of macromolecules. This increase in the use of IRMPD can be attributed to advances in the characteristics of IR lasers, particularly the IR optics. The extension of this photodissociation method to other mass spectrometers, including linear and quadrupole ion traps, orbitraps, and hybrid TOFs is another reason for the increasing use of this method in peptide/protein fragmentation This activation method combined with tandem mass spectrometry (MS/MS) can provide fairly precise information on the type and sites of various PTMs of individual amino acids as well as on amino acids within peptides and proteins. Some of the modifications examined by this method include phosphorylation, sulfation, acetylation, methylation, glycosylation, and disulfide bond formation [82,83][54][55]. The photodissociation of the precursor molecular ions containing one or more of these PTMs is induced by the presence of one or more reactant group(s) (e.g., -OH, -C-H, -S-H, -S-CH3) responsible for the absorption of the irradiating photons. The second method uses ultraviolet photodissociation (UVPD) at different wavelengths. For instance, 193 nm was first applied in the 1980s [84,85][56][57]; however, such an application was limited to specific peptides performed on Fourier transform ion cyclotron resonance (FTICR) mass spectrometers. Over the last ten years and due to advances in UV lasers and in MS instrumentation, the technique has witnessed a resurgence, represented in its use in a wide range of instruments, including TOF [86][58], linear ion traps [87][59], and more recently in the orbitrap [88][60]. UVPD mainly uses two wave lengths, 157 and 193 nm; however, the wavelength 213 nm has been used to sequence intact proteoforms in top-down proteomics [89][61]. In UVPD, due to the absorption of a single photon having 5–7 eV energy by the precursor ion, fragmentation proceeds either through electronic dissociative excitation or through the interconversion of the electronic excitation to vibrational excitation, thus leading to bond rapture, which is the major fragmentation pathway [90,91][62][63].2. Analysis of Some ATP-Binding Cassette (ABC) Transporters
The introduction of two soft ionization techniques, namely electrospray ionization (ESI) [23][64] and matrix-assisted laser desorption/ionization (MALDI) [24][65], was a major step towards a diffused use of mass spectrometry (MS) in biological and biochemical worlds. Over the last two decades, advances in MS instrumentation together with more refined labelling protocols, the combination of ion mobility with MS, and more frequent use of electron-based and photon-based fragmentation methods paved the way for what is known today as structural mass spectrometry. With these innovative developments, mass spectrometry can be applied for the structural characterization of a wide range of macromolecules and in some cases their respective assemblies. It is relevant to point out that continuous progress in sample preparations, including the use of a new class of detergents, extended the application of MS to membrane proteins, which are not water soluble.
2.1. Monitoring the Conformation of P-glycoprotein
P-glycoprotein (also known as ABCB1 and MDR1) is the first identified mammalian ABC transporter protein, discovered almost half a century ago [93][66]. It has a ~170 kDa molecular weight and is comprised of two homologous halves, each containing a nucleotide-binding domain (NBD) and a transmembrane domain (TMD) [94][67]. The transmembrane domain contains a hydrophobic cavity, accessed by portals in the membrane, that binds cytotoxic compounds as well as lipids. This protein is expressed in multiple key organs, such as the small intestine, blood–brain barrier, kidney, and liver. Therefore, P-glycoprotein mediated drug–drug interactions can occur in various organs and in tissues. The overexpression of P-glycoprotein has been linked to chemotherapy failure in various cancers, including kidney, colon, and liver. A hallmark characteristic of this transporter protein is its ability to bind and transport a wide range of structurally different molecules in the molecular mass range, 100 to 4000 Da, a range which covers most if not all anticancer and antimicrobial drugs currently in use [95][68]. The transportation of these molecules across the membrane was found to coincide with changes in the size and shape of a large multi-specific drug-binding pocket [94][67]. Although its physiological function(s) are yet to be fully understood, the well-recognized role of this protein in mediating multidrug resistance in many types of cancers has made it an attractive therapeutic target [96][69]. In recent years it has been demonstrated that hydrogen–deuterium exchange mass spectrometry (HDX-MS) is ideal for studying protein dynamics as they occur in solution. Proteins are known to assume a wide range of conformations in solution, which means that every amide proton will eventually exchange with deuterium. Measuring this rate of exchange as a function of time would result in a highly informative picture of the regional dynamics. Information provided by this technique are fundamental for the understanding of the functioning mechanism of ABC exporters, including P-gp, and complement the snapshots of the catalytic cycle provided by high-resolution 3D structures. In a recent article, HDX-MS was used to investigate the conformational states as well as the dynamics and mechanism of transportation of P-glycoprotein. The authors detected three distinct conformational states and obtained information on transporter dynamics with a sequence coverage over 85% [100][70]. This high sequence coverage and the use of bio-layer interferometry to measure nucleotide affinity furnished further information on the states of conformation, dynamics, and mechanism of transportation by this protein. Since its discovery almost 50 years ago, P-glycoprotein has been structurally characterized by X-ray, cryo-electron spectroscopy, and, more recently, by HDX-MS. This structural information, together with data furnished by analyses of clinical samples provided by patients undergoing chemotherapy and molecularly targeted therapy, gives a clearer picture of the role of this protein in MDR. The initial enthusiasm that such new information could lead to the discovery of P-glycoprotein inhibitors was quickly dampened.2.2. Breast Cancer Resistance Protein (ABCG2)
The human ATP-binding cassette transporter ABCG2, also known as breast cancer resistance protein (BCRP), is a key player in anticancer resistance and in physiological detoxification across tissue barriers [107][71]. Despite numerous investigations, the molecular mechanism of substrate transport by this protein remains to be fully clarified. The activities of this protein are known to affect the pharmacokinetics of commonly used drugs as well as interfering in the delivery of various therapeutics into tumor cells, thus contributing to multidrug resistance [108][72]. Unlike most of the other ABC transporters, which usually have two nucleotide-binding domains and two transmembrane domains, ABCG2 consists of only one nucleotide-binding domain followed by one transmembrane domain. Thus, ABCG2 has been thought to be a half-transporter that may function as a homodimer (molecular weight approximately 144 kDa). The first high-resolution structure of human ABCG2 determined by cryo-electron microscopy was reported over five years ago [109][73]. The structure shows two cholesterol molecules bound in the multidrug-binding pocket that is located in a central, hydrophobic, inward-facing translocation pathway between the transmembrane domains. Within the same year, another research group used the X-ray crystal structure of ABCG5/G8 to generate a model of ABCG2 [109][73]. To validate structural and mechanistic predictions of their model, the authors used extensive molecular–genetic analyses. The ABCG2 structure contains two apparent cavities. The architecture of the central cavity includes the intracellular loop1, the elbow helix, and residues facing the cavity from transmembrane helices and the NBD dimer. The central cavity is part of the transmission interface, which is essential for ABCG2 drug transport. The smaller upper cavity is part of what is called the extracellular polar roof. These two cavities were found to be separated by two leucine residues, facing their equivalent residues in the core of the symmetric ABCG2 dimer [108,109,110,111][72][73][74][75]. The link between ABCG2 and resistance to anticancer drugs has been discussed in a number of studies conducted on cancer cell lines. In one of these studies from over 25 years ago, overexpression of ABCG2 in model cancer cell lines (MCF-7 breast cancer cells) was shown to cause resistance to a number of anticancer drugs, including doxorubicin and daunorubicin [114][76]. Over the last ten years, a new class of targeted anticancer drugs has emerged. One of these drugs, sonidegib (trade name odomzo), was approved by the FDA in 2015 for the treatment of adults with advanced basal cell carcinoma. This drug was tested as an inhibitor of the transport functions of both ABCB1 and ABCG2 [115][77]. The authors reported that in accumulation studies, the transport functions of both proteins were effectively inhibited by sonidegib.43. Commenting on Inhibitors of P-glycoprotein
Four generations of potential inhibitors of P-Glycoprotein have been developed and tested over a period of 40 years [104][78]. During this period, many potential inhibitors were extensively investigated, but none of them obtained the approval of either the FDA (Food and Drug Administration) or the EMA (European Medicines Agency). Most experts in the field argue that the failure of these inhibitors to restore sensitivity to chemotherapy reside in their poor selectivity, low potency, inherent toxicity, and/or adverse pharmacokinetic interaction with anticancer drugs. However, it has to be said that despite the poor results of years of research, a number of studies during the same period indicated that under specific conditions the combination of potential inhibitors of MDR1 and ABCG2 with certain chemotherapeutics resulted in increased drug accumulation and drug resistance reversal. - In vitro analyses: Cell lines are often used in research in place of primary cells. These in vitro analyses offer the advantages of low costs, ease of use, providing an unlimited supply of materials, and bypassing ethical concerns associated with the use of animal and human samples [101][79]. That said, the use of cell lines in cancer research has some drawbacks. A major drawback of in vitro models is their failure to capture the full reality of in vivo systems. For example, such models may not account for likely interactions between cells and other biological and biochemical processes. Cancer cell lines are often derived from a single subtype of tumor, which very often does not reflect the heterogeneity of tumors in patients. - The multifactorial nature of drug resistance is another factor to be considered as being partly responsible for the poor results in the search for specific inhibitors of P-gp. There is strong evidence that high expression of this protein remains one of the main reasons for the poor response to chemotherapy in many cancer types. That said, limiting the search for inhibitors to a single drug resistance mechanism is unlikely to restore chemotherapeutic sensitivity. As mentioned earlier, this protein is part of a superfamily containing at least 48 members. The existing literature shows that only three members have been studied in some detail. - Ligand–protein interaction. Over the last 40 years, chemical, biochemical, and biological studies have provided a wealth of information on the mechanisms of interaction between a limited number of ABC transporter proteins and various ligands. The complexity of such interaction renders such information insufficient to identify the precise mechanism(s) of such interactions. Identification of the basic physical and chemical characteristics, which mediate ligand–protein interaction are highly relevant for inhibitor(s) selection. - Level of expression. Accurate detection and quantification of ATP-binding cassette (ABC) transporters are highly relevant to the assessment of their role in chemoresistance. Such a parameter becomes more critical when expression is measured in vivo. Most studies conducted on cell lines indicate that the level of expression of one or more of these proteins varies from one tumor to another. The same literature shows that the detection and quantification of these levels have been mainly performed by reverse transcription–polymerase chain reaction (RT-PCR) for mRNA expression and by immunohistochemistry (IHC) for protein expression. - The good and the bad. ATP-binding cassette (ABC) transporters are expressed in both healthy and cancerous cells. Within the healthy cells, some of these proteins play a crucial physiological role in protecting tissues from toxic xenobiotics and endogenous metabolites, and also affect the uptake and distribution of many clinically important drugs. The same proteins form a major component of the blood–brain barrier and restrict the uptake of drugs from the intestine. In cancerous cells, on the other hand, the overexpression of these proteins has been strongly linked to MDR. Although MDR is multifactorial in origin, it is strongly associated with the overexpression of ATP-binding cassette (ABC) transporters.54. Conclusions
Mass spectrometry is becoming an increasingly important tool in the field of structural biology. In recent years, the role of this technique has been enhanced by more frequent use of ion mobility-MS, HDX-MS, and electron/photon-based ion activation methods. These techniques, together with X-ray crystallography and cryo-electron microscopy, are furnishing much-needed information on the structure of membrane proteins, including ABC transporters. Information provided by these techniques together with those acquired through advanced molecular dynamics simulation, molecular docking studies, and protein–ligand investigations will no doubt contribute to the ongoing scientific research to fill the gaps, which for over 40 years have hindered the identification of specific and effective clinical inhibitors of ABC transporters.References
- Almén, M.S.; Nordström, K.J.; Fredriksson, R.; Schiöth, H.B. Mapping the human membrane proteome: A majority of the human membrane proteins can be classified according to function and evolutionary origin. BMC Biol. 2009, 7, 50.
- Wallin, E.; von Heijne, G. Genome-wide analysis of integral membrane proteins from eubacterial, archaean, and eukaryotic organisms. Protein Sci. 1998, 7, 1029–1038.
- Arinaminpathy, Y.; Khurana, E.; Engelman, D.M.; Gerstein, M.B. Computational analysis of membrane proteins: The largest class of drug targets. Drug Discov. Today 2009, 14, 1130–1135.
- Overington, J.P.; Al-Lazikani, B.; Hopkins, A.L. Opinion—How many drug targets are there? Nat. Rev. Drug Discov. 2006, 5, 993–996.
- Billings, E.; Lundquist, K.; Overly, C.; Srinivasan, K.; Noinaj, N. Structure Determination of Membrane Proteins Using X-ray Crystallography. Methods Mol. Biol. 2021, 2302, 101–136.
- Hernando-Amado, S.; Blanco, P.; Alcalde-Rico, M.; Corona, F.; Reales-Calderón, J.A.; Sánchez, M.B.; Martínez, J.L. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resist. Update 2016, 28, 13–27.
- Sun, J.; Deng, Z.; Yan, A. Bacterial multidrug efflux pumps: Mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 2014, 453, 254–267.
- Levin, B.R.; Rozen, D.E. Non-inherited antibiotic resistance. Nat. Rev. Microbiol. 2006, 4, 556–562.
- Corona, F.; Martinez, J.L. Phenotypic Resistance to Antibiotics. Antibiotics 2013, 2, 237–255.
- Gottesman, M.M.; Fojo, T.; Bates, S.E. Multidrug resistance in cancer: Role of ATP–dependent transporters. Nat. Rev. Cancer 2002, 2, 48–58.
- Ambudkar, S.V.; Dey, S.; Hrycyna, C.A.; Ramachandra, M.; Pastan, I.; Gottesman, M.M. Biochemical, cellular, and pharmacological aspects of the multidrug transporter. Annu. Rev. Pharmacol. Toxicol. 1999, 39, 361–398.
- Choi, C.-H. ABC transporters as multidrug resistance mechanisms and the development of chemosensitizers for their reversal. Cancer Cell Int. 2005, 5, 30.
- Thomas, H.; Coley, H.M. Overcoming multidrug resistance in cancer: An update on the clinical strategy of inhibiting p-glycoprotein. Cancer Control 2003, 10, 159–165.
- Kühlbrandt, W. The Resolution Revolution. Science 2014, 343, 1443–1444.
- Thompson, R.F.; Walker, M.; Siebert, C.A.; Muench, S.P.; Ranson, N.A. An introduction to sample preparation and imaging by cryo-electron microscopy for structural biology. Methods 2016, 100, 3–15.
- Konermann, L.; Pan, J.; Liu, Y.H. Hydrogen exchange mass spectrometry for studying protein structure and dynamics. Chem. Soc. Rev. 2011, 40, 1224–1234.
- Calabrese, A.N.; Radford, S.E. Mass spectrometry-enabled structural biology of membrane proteins. Methods 2018, 147, 187–205.
- Laganowsky, A.; Reading, E.; Hopper, J.T.S.; Robinson, C.V. Mass spectrometry of intact membrane protein complexes. Nat. Protoc. 2013, 8, 639–651.
- Lanucara, F.; Holman, S.W.; Gray, C.J.; Eyers, C.E. The power of ion mobility-mass spectrometry for structural characterization and the study of conformational dynamics. Nat. Chem. 2014, 6, 281–294.
- Konijnenberg, A.; Butterer, A.; Sobott, F. Native ion mobility-mass spectrometry and related methods in structural biology. BBA—Proteins Proteom. 2013, 1834, 1239–1256.
- Zhong, Y.; Hyung, S.-J.; Ruotolo, B.T. Ion mobility–mass spectrometry for structural proteomics. Expert Rev. Proteom. 2012, 9, 47–58.
- Engen, J.R.; Wales, T.E. Analytical Aspects of Hydrogen Exchange Mass Spectrometry. Annu. Rev. Anal. Chem. 2015, 8, 127–148.
- Katta, W.; Chait, B.T. Hydrogen/Deuterium Exchange Electrospray Ionization Mass Spectrometry: A Method for Probing Protein Conformational Changes in Solution. J. Am. Chem. Soc. 1993, 115, 6317–6321.
- Hammerschmid, D.; Calvaresi, V.; Bailey, C.; Russell, B.; Argyris Politis, L.; Morris, M.; Denbigh, L.; Anderson, M.; Reading, E. Chromatographic Phospholipid Trapping for Automated H/D Exchange Mass Spectrometry of Membrane Protein–Lipid assemblies. Anal. Chem. 2023, 95, 3002–3011.
- Calvaresi, V.; Redsted, A.; Norais, N.; Rand, K.D. Hydrogen–Deuterium exchange mass spectrometry with integrated size-exclusion chromatography for analysis of complex protein samples. Anal. Chem. 2021, 93, 11406–11414.
- Donnarumma, D.; Maestri, C.; Giammarinaro, P.I.; Capriotti, L.; Bartolini, E.; Veggi, D.; Petracca, R.; Scarselli, M.; Norais, N. Native state organization of outer membrane porins unraveled by HDx-MS. J. Proteome Res. 2018, 17, 1794–1800.
- Mysling, S.; Salbo, R.; Ploug, M.; Jørgensen, T.J.D. Electrochemical reduction of disulfide-containing proteins for hydrogen/deuterium exchange monitored by mass spectrometry. Anal. Chem. 2014, 86, 340–345.
- Bobst, C.E.; Kaltashov, I.A. Enhancing the Quality of H/D Exchange measurements with mass spectrometry detection in disulfide-rich proteins using electron capture dissociation. Anal. Chem. 2014, 86, 5225–5231.
- Burns, J.A.; Butler, J.C.; Moran, J.; Whitesides, G.M. Selective reduction of disulfides by tris(2-carboxyethyl)phosphine. J. Org. Chem. 1991, 56, 2648–2650.
- Larsen, M.R.; Højrup, P.; Roepstorff, P. Characterization of Gel-separated Glycoproteins Using Two-step Proteolytic Digestion Combined with Sequential Microcolumns and Mass Spectrometry. Mol. Cell. Proteom. 2005, 4, 107–119.
- Darula, Z.; Medzihradszky, K.F. Glycan side reaction may compromise ETD-based glycopeptide identification. J. Am. Soc. Mass Spectrom. 2014, 25, 977–987.
- Houel, S.; Hilliard, M.; Yu, Y.Q.; McLoughlin, N.; Martin, S.M.; Rudd, P.M.; Williams, J.P.; Chen, W. N- and O-glycosylation analysis of etanercept using liquid chromatography and quadrupole time-of-flight mass spectrometry equipped with electron-transfer dissociation functionality. Anal. Chem. 2014, 86, 576–584.
- Rand, K.D. Pinpointing changes in higher-order protein structure by hydrogen/deuterium exchange coupled to electron transfer dissociation mass spectrometry. Int. J. Mass Spectrom. 2013, 338, 2–10.
- Javed, W.; Griffiths, D.; Politis, A. Hydrogen/deuterium exchange-mass spectrometry of integral membrane proteins in native-like environments: Current scenario and the way forward. Essays Biochem. 2023, 67, 187–200.
- Majeed, S.; Ahmad, A.B.; Sehar, U.; Georgieva, E.R. Lipid Membrane Mimetics in Functional and Structural Studies of Integral Membrane Proteins. Membranes 2021, 11, 685.
- Karasek, F.W. Drift-mass spectrometer. Res. Dev. 1970, 21, 25–27.
- Alge, E.; Villinger, H.; Lindinger, W. Drift tube investigations on the reactions of O2 + with CH4 and of CO2 + with NO in various buffer gases. Plasma Chem. Plasma Process. 1981, 1, 65–71.
- Smith, D.P.; Giles, K.; Bateman, R.H.; Radford, S.E.; Ashcroft, A.E. Monitoring Copopulated Conformational States during Protein Folding Events Using Electrospray Ionization-Ion Mobility Spectrometry-Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2007, 18, 2180–2190.
- May, J.C.; Morris, C.B.; McLean, J.A. Ion mobility collision cross section compendium. Anal. Chem. 2017, 89, 1032–1044.
- Wyttenbach, T.; Bowers, M.T. Structural stability from solution to the gas phase: Native solution structure of ubiquitin survives analysis in a solvent-free ion mobility–mass spectrometry environment. J. Phys. Chem. B 2011, 115, 12266–12275.
- Ruotolo, B.T.; Robinson, C.V. Aspects of native proteins are retained in vacuum. Curr. Opin. Chem. Biol. 2006, 10, 402–408.
- Yost, R.A.; Enke, C.G. Selected ion fragmentation with a tandem quadrupole mass spectrometer. J. Am. Chem. Soc. 1978, 100, 2274–2275.
- Zubarev, R.A.; Horn, D.M.; Fridriksson, E.K.; Kelleher, N.L.; Kruger, N.A.; Lewis, M.A.; Carpenter, B.K.; McLafferty, F.W. Electron capture dissociation for structural characterization of multiply charged protein cations. Anal. Chem. 2000, 72, 563–573.
- Syrstad, E.A.; Turecček, F. Toward a general mechanism of electron capture dissociation. J. Am. Soc. Mass Spectrom. 2005, 16, 208–224.
- Zubarev, R.A.; Kelleher, N.L.; McLafferty, F.W. Electron capture dissociation of multiply charged protein cations. A nonergodic process. J. Am. Chem. Soc. 1998, 120, 3265–3266.
- Qi, Y.; Volmer, D.A. Electron-based fragmentation methods in mass spectrometry: An overview. Mass Spectrom. Rev. 2017, 36, 4–15.
- Syka, J.E.P.; Coon, J.J.; Schroeder, M.J.; Shabanowitz, J.; Hunt, D.F. Peptide and Protein sequence analysis by electron transfer dissociation mass spectrometry. Proc. Natl. Acad. Sci. USA 2004, 101, 9528–9533.
- Pitteri, S.J.; Chrisman, P.A.; McLuckey, S.A. Electron-Transfer Ion/Ion Reactions of Doubly Protonated Peptides: Effect of Elevated Bath Gas Temperature. Anal. Chem. 2005, 77, 5662–5669.
- Ko, B.J.; Brodbelt, J.S. Enhanced electron transfer dissociation of peptides modified at C-terminus with fixed charges. J. Am. Soc. Mass Spectrum. 2012, 23, 1991–2000.
- Frey, B.L.; Ladror, D.T.; Sondalle, S.B.; Krusemark, C.J.; Jue, A.L.; Coon, J.J.; Smith, L.M. Chemical derivatization of peptide carboxyl groups for highly efficient electron transfer dissociation. J. Am. Soc. Mass Spectrom. 2013, 24, 1710–1721.
- Swaney, D.L.; McAlister, G.C.; Coon, J.J. Decision tree–driven tandem mass spectrometry for shotgun proteomics. Nat. Methods 2008, 5, 959–964.
- Frese, C.K.; Altelaar, A.F.M.; Hennrich, M.L.; Nolting, D.; Zeller, M.; Griep-Raming, J.; Heck, A.J.R.; Mohammed, S. Improved peptide identification by targeted fragmentation using CID, HCD and ETD on an LTQ-orbitrap velos. J. Proteome Res. 2011, 10, 2377–2388.
- Woodin, R.L.; Bomse, D.S.; Beauchamp, J.L. Multiphoton dissociation of molecules with low power continuous wave infrared laser radiation. J. Am. Chem. Soc. 1978, 100, 3248–3250.
- Maitre, P.; Scuderi, D.; Corinti, D.; Chiavarino, B.; Crestoni, M.E.; Fornarini, S. Applications of Infrared Multiple Photon Dissociation (IRMPD) to the Detection of Posttranslational Modifications. Chem. Rev. 2020, 120, 3261–3295.
- Greisch, J.-F.; van der Laarse, S.A.; Heck, A.J. Enhancing Top-Down Analysis Using Chromophore-Assisted Infrared Multiphoton Dissociation from (Phospho)peptides to Protein Assemblies. Anal. Chem. 2020, 92, 15506–15516.
- Bowers, W.D.; Delbert, S.S.; Hunter, R.L.; Mciver, R.T. Fragmentation of oligopeptide ions using ultraviolet laser radiation and fourier transform mass spectrometry. J. Am. Chem. Soc. 1984, 106, 7288–7289.
- Hunt, D.F.; Shabanowitz, J.; Yates, J.R. Peptide sequence analysis by laser photodissociation Fourier transform mass spectrometry. J. Chem. Soc. Chem. Commun. 1987, 548–550.
- Thompson, M.S.; Cui, W.; Reilly, J.P. Fragmentation of Singly Charged Peptide Ions by Photodissociation at λ = 157 nm. Angew. Chem. Int. Ed. 2004, 43, 4791–4794.
- Cannon, J.R.; Cammarata, M.B.; Robotham, S.A.; Cotham, V.C.; Shaw, J.B.; Fellers, R.T.; Early, B.P.; Thomas, P.M.; Kelleher, N.L.; Brodbelt, J.S. Ultraviolet photodissociation for characterization of whole proteins on a chromatographic time scale. Anal. Chem. 2014, 86, 2185–2192.
- Smyrnakis, A.; Levin, N.; Kosmopoulou, M.; Jha, A.; Fort, K.; Makarov, A.; Papanastasiou, D.; Mohammed, S. Characterization of an Omnitrap-Orbitrap Platform Equipped with Infrared Multiphoton Dissociation, Ultraviolet Photodissociation, and Electron Capture Dissociation for the Analysis of Peptides and Proteins. Anal. Chem. 2023, 95, 12039–12046.
- Fornelli, L.; Srzentić, K.; Toby, T.K.; Doubleday, P.F.; Huguet, R.; Mullen, C.; Melani, R.D.; dos Santos Seckler, H.; DeHart, C.J.; Weisbrod, C.R.; et al. Thorough Performance Evaluation of 213 nm Ultraviolet Photodissociation for Top-down Proteomics. Mol. Cell. Proteom. 2020, 19, 405–420.
- Julian, R.R.; Amster, I.J.; Kong, X.; Brodbelt, J.S.; Jørgensen, T.J.D.; Wysocki, V.H.; Hendrickson, C.L.; Santos, I.; Shaw, J.B.; Boyarkin, O.V.; et al. The mechanism behind top-down uvpd experiments: Making sense of apparent contradictions. J. Am. Soc. Mass Spectrom. 2017, 28, 1823–1826.
- Zabuga, A.V.; Kamrath, M.Z.; Boyarkin, O.V.; Rizzo, T.R. Fragmentation mechanism of UV-excited peptides in the gas phase. J. Chem. Phys. 2014, 141, 154309.
- Yamashita, M.; Fenn, J.B. Electrospray ion source. Another variation on the free-jet theme. J. Phys. Chem. 1984, 88, 4451–4459.
- Karas, M.; Bachmann, D.; Hillenkamp, F. Influence of the Wavelength in High-Irradiance Ultraviolet Laser Desorption Mass Spectrometry of Organic Molecules. Anal. Chem. 1985, 57, 2935–2939.
- Juliano, R.; Ling, V. A surface glycoprotein modulating drug permeability in Chinese hamster ovary cell mutants. BBA—Biomembr. 1976, 455, 152–162.
- Ward, A.B.; Szewczyk, P.; Grimard, V.; Lee, C.-W.; Martinez, L.; Doshi, R.; Caya, A.; Villaluz, M.; Pardon, E.; Cregger, C.; et al. Structures of P-glycoprotein reveal its conformational flexibility and an epitope on the nucleotide-binding domain. Proc. Natl. Acad. Sci. USA 2013, 110, 13386–13391.
- Schinkel, A.H.; Jonker, J.W. Mammalian drug efflux transporters of the ATP binding cassette (ABC) family: An overview. Adv. Drug Deliv. Rev. 2003, 55, 3–29.
- Marquez, B.; Van Bambeke, F. ABC multidrug transporters: Target for modulation of drug pharmacokinetics and drug-drug interactions. Curr. Drug Targets 2011, 12, 600–620.
- Kopcho, N.; Geoffrey Chang, G.; Komives, E.A. Dynamics of ABC transporter P-glycoprotein in three conformational states. Sci. Rep. 2019, 9, 15092.
- Van Herwaarden, A.E.; Schinkel, A.H. The function of breast cancer resistance protein in epithelial barriers, stem cells and milk secretion of drugs and xenotoxins. Trends Pharmacol. Sci. 2006, 27, 10–16.
- Taylor, N.M.I.; Manolaridis, I.; Jackson, S.M.; Kowal, J.; Stahlberg, H.; Locher, K.P. Structure of the human multidrug transporter ABCG2. Nature 2017, 546, 504–509.
- Khunweeraphong, N.; Stockner, T.; Kuchler, K. The structure of the human ABC transporter ABCG2 reveals a novel mechanism for drug extrusion. Sci. Rep. 2017, 7, 13767.
- Jackson, S.M.; Manolaridis, I.; Kowal, J.; Zechner, M.; Taylor, N.M.I.; Bause, M.; Bauer, S.; Bartholomaeus, R.; Bernhardt, G.; Koenig, B.; et al. Structural basis of small-molecule inhibition of human multidrug transporter ABCG2. Nat. Struct. Mol. Biol. 2018, 25, 333–340.
- Manolaridis, I.; Jackson, S.M.; Taylor, N.M.I.; Kowal, J.; Stahlberg, H.; Locher, K.P. Cryo-EM structures of a human ABCG2 mutant trapped in ATP-bound and substrate-bound states. Nature 2018, 563, 426–430.
- Doyle, L.A.; Yang, W.; Abruzzo, L.V.; Krogmann, T.; Gao, Y.; Rishi, A.K.; Ross, D.D. A multidrug resistance transporter from human MCF-7 breast cancer cells. Proc. Natl. Acad. Sci. 1998, 95, 15665–15670.
- Zhang, Y.; Vagiannis, D.; Budagaga, Y.; Sabet, Z.; Hanke, I.; Rozkoš, T.; Hofman, J. Sonidegib potentiates the cancer cells’ sensitivity to cytostatic agents by functional inhibition of ABCB1 and ABCG2 in vitro and ex vivo. Biochem. Pharmacol. 2022, 199, 115009.
- Modok, S.; Mellor, H.R.; Callaghan, R. Modulation of multidrug resistance efflux pump activity to overcome chemoresistance in cancer. Curr. Opin. Pharmacol. 2006, 6, 350–354.
- Crowley, E.; McDevitt, C.A.; Callaghan, R. Generating Inhibitors of P-Glycoprotein: Where to, now? Multi-Drug Resist. Cancer 2010, 596, 405–432.