Tight junctions (TJ) are cell–cell adhesive structures that define the permeability of barrier-forming epithelia and endothelia. In contrast to this seemingly static function, TJs display a surprisingly high molecular complexity and unexpected dynamic regulation, which allows the TJs to maintain a barrier in the presence of physiological forces and in response to perturbations. Cell–cell adhesion receptors play key roles during the dynamic regulation of TJs. They connect individual cells within cellular sheets and link sites of cell–cell contacts to the underlying actin cytoskeleton.
- cell–cell adhesion
- JAM-A
- tight junction
- integral membrane proteins
- CRB3
- claudins
- TAMPs
- IgSF proteins
1. Introduction
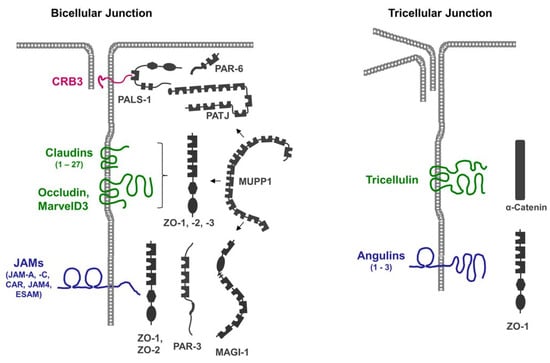
2. CRB3
CRB3 is a member of the vertebrate Crumbs family of proteins, which are homologous to Drosophila Crumbs. CRB3 and has a very short extracellular region that consists of 33 amino acids (AA), a single transmembrane region, and a short cytoplasmic region of 40 AA [20][26]. In addition to its localization at TJs, CRB3 is also localized at the apical membrane domain of epithelial cells [21][27]. As opposed to Drosophila Crumbs, the vertebrate CRB3 isoform localized at the TJs is most likely not involved in homophilic or heterophilic interactions [22][28].3. Claudins
Claudins are a family of tetraspan transmembrane proteins comprising 27 members [23][24][29,30]. Notably, claudins support cell aggregation when expressed in fibroblasts, indicating that their adhesive activity not only regulates their localization at homotypic cell–cell contacts but also contributes to the physical cell–cell adhesion at the TJs [25][31]. A central property of claudins is their ability to multimerize by interacting both in cis and in trans with either the same or a different claudin family member to form strands that are visible via freeze-fracture electron microscopy (EM) [26][27][32,33]. Claudin-based strands can act as occluding barriers for water and small solutes, as well as anion- or cation-selective paracellular channels, and are the principal paracellular permeability regulators of epithelial and endothelial barriers [28][34].4. TAMPs
Members of the TAMP family include occludin, tricellulin/MarvelD2, and MarvelD3 [29][35]. Similar to claudins, TAMPs are tetraspan transmembrane proteins, but based on their sequence homologies, they are not related to claudins. Their characteristic feature is a conserved four-transmembrane “MAL and related proteins for vesicle trafficking and membrane link” (Marvel) domain [30][36]. Of the three TAMPs, tricellulin is unique as it is enriched at sites of contact between three cells (tricellular TJs, tTJs) [31][37]. Heterotypic interactions between tricellulin and MarvelD3, as well as between occludin and MarvelD3, have been described via co-immunoprecipitation (CoIP) and via Förster resonance energy transfer (FRET) experiments. These interactions most likely occur in cis [29][32][35,38]. A homophilic interaction in trans has been found for occludin but not for tricellulin or MarvelD3 [32][33][38,39]. While several studies have suggested that TAMPs per se are not essential for the development of an epithelial barrier function [34][35][36][40,41,42], recent findings indicate that tricellulin is required for the establishment of the barrier function in mammary gland-derived epithelial cells [37][43].5. IgSF Proteins
IgSF members localized at the TJs include the junctional adhesion molecule (JAM) family members JAM-A and JAM-C [38][44], and the JAM-related adhesion molecules Coxsackie- and Adenovirus-Receptor (CAR), JAM4, CAR-like membrane protein (CLMP), and Endothelial Cell-Selective Adhesion Molecule (ESAM, in endothelial cells) [39][40][41][42][45,46,47,48]. All these IgSF proteins can undergo trans-homophilic interactions which stabilize their localization at cell–cell contacts. In addition, the trans-homophilic activities of CAR, JAM4, CLMP, and ESAM support cell aggregation after transfection in cells [39][40][41][43][45,46,47,49], suggesting that their adhesive activities contribute to the strength of the physical interaction at the TJs. Angulins (Angulin-1, -2, -3) are IgSF proteins with a single N-terminal Ig-like domain [44][50]. Among all other IgSF proteins localized at the TJs, angulins are unique in that they are enriched at the tTJs [45][51]. A main function of all three angulins is to recruit tricellulin to the tTJs [45][46][51,52]. The ectopic expression of angulin-1 in L cell fibroblasts resulted in angulin-1 enrichment at cell–cell contacts, suggesting that angulin-1 is engaged in homophilic or heterophilic trans interactions [46][52]. Strikingly, many of the integral membrane proteins localized at the TJs contain a C-terminal PDZ domain-binding motif (PBM), through which they can interact with the PDZ domains present in many TJ-localized scaffolding proteins (Table 1).| Integral Membrane Protein | Size (AA) | Cytoplasmic Region (AA) | COOH-Terminal Residues (PBM) |
|---|---|---|---|
| CRB3 | 120 | 40 | - EERLI (+) |
| JAM-A | 299 | 40 | - SSFLV (+) |
| JAM-C | 310 | 48 | - SSFVI (+) |
| CAR | 365 | 107 | - DGSIV (+) |
| JAM4 | 407 | 122 | - NTTVV (+) |
| ESAM | 390 | 121 | - AGSLV (+) |
| Angulin-1/LSR | 649 | 369 | - ESLVV (+) |
| Angulin-2/ILDR1 | 546 | 358 | - RSVVI (+) |
| Angulin-3/ILDR2 | 639 | 432 | - MSLVV (+) |
| Claudins (1–26) | 207–305 | 27–66 | - XXXYF (+) |
| Occludin | 522 | 257 | - DRQKT (−) |
| Tricellulin | 558 | 196 | - VQGYS (−) |
| MarvelD3 | 401 | 20 | - EMFEF (+) |
The presence of multiple PDZ domains in many scaffolding proteins localized at the TJs (Figure 1), combined with the promiscuity of PDZ domains in ligand binding, allows for the incorporation of several integral membrane proteins in a single scaffolding protein-organized complex. Vice versa, individual membrane proteins can recruit and assemble distinct protein complexes at the TJ. These biochemical properties may, in part, explain the high complexity and dynamics of protein complexes at the TJs [21][27].