Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Andrey Sergeevich Erst and Version 2 by Sirius Huang.
Despite the limited geographic range of Eranthis plants, it is possible to search for active substances, develop methods for biological and chemical synthesis of the isolated substances, and create a finished therapeutic substance based on them. Seven out of ~14 species found in Asia and parts of Europe have been studied to various degrees. Here, data are presented on the diversity of sets of chromones, furochromones, triterpene saponins, coumarins, and other classes of secondary metabolites of Eranthis species according to the literature. For new compounds—isolated from Eranthis for the first time—structural formulas are also provided.
- Eranthis
- chromone
- furochromone
- triterpene saponin
- coumarin
- biological activity
1. Introduction
According to molecular and morphological data, the tribe Cimicifugeae Torrey & Gray belongs to the family Ranunculaceae Juss. and includes four recognized genera and ~49 species: Actaea L. (32 species), Anemonopsis Siebold et Zucc. (one species), Beesia Balf. f. et W. W. Sm. (two species), and Eranthis Salisb. (14 species) [1][2][3][1,2,3]. Most of these species occur mainly in the northern hemisphere and are perennial herbs [4]. The taxonomic position of the genera Eranthis and Beesia has been a matter of systematic uncertainty within the tribal rank in the Ranunculaceae family. According to morphological information, Beesia has been assigned to three different tribes (Helleboreae DC., Actaeeae Spach, and Trollieae Schröd.) by intuitive taxonomic techniques but has seldom been included in cladistic analyses [5][6][7][5,6,7]. By contrast, the Eranthis genus has consistently been assigned to the Helleboreae tribe or as the only genus to the tribe Eranthideae T. Duncan & Keener in morphological classifications but always has been a sister taxon to plants of the Actaeeae tribe in cladistic analyses [8]. The genus Eranthis consists of 8–14 species growing in southern Europe and temperate Asia [9][10][11][9,10,11]. Traditionally, the genus has been subdivided into two sections: Eranthis sect. Eranthis and E. sect. Shibateranthis (Nakai) Tamura [12]. The type section Eranthis is characterized by plants with tubers, yellow sepals, and emarginate or slightly bilobate upper petal margins without pseudonectaries (Figure 1) [6][11][6,11]. The section Eranthis in Europe includes E. hyemalis (L.) Salisb. and E. bulgarica (Stef.) Stef., whereas in Southwest and West Asia, it includes E. cilicica Schott et Kotschy, E. kurdica Rukšāns, E. longistipitata Regel, and E. iranica Rukšāns et Zetterl. [13][14][15][16][13,14,15,16]. The section Shibateranthis has long-lived tubers, white sepals, and bilobate or forked petal margins with pseudonectaries (Figure 1) [6][17][6,17]. Representatives of this section have a natural geographic range in temperate North and East Asia (E. albiflora Franch., E. byunsanensis B.Y. Sun, E. lobulata W.T.Wang, E. pinnatifida Maxim., E. pungdoensis B.U. Oh, E. sibirica DC., E. stellata Maxim., and E. tanhoensis Erst) [10][11][10,11].

Figure 1. Species of the genus Eranthis. (A) E. longistipitata, (B) E. cilicica, (C) E. hyemalis, (D) E. sibirica, (E) E. tanhoensis, and (F) E. stellata.
Plants of the tribe Cimicifugeae are some of the richest sources of various active ingredients and of therapeutic and health-promoting substances. The value of the constituents has been confirmed by many years of use in East Asian countries in folk medicine. Thus, it is important to integrate new technologies into research on Cimicifugeae, both for the sustainable use of pharmaceutical resources from Cimicifugeae and for a search for new compounds with potential clinical efficacy and fewer adverse effects [18][19][20][18,19,20]. In the tribe Cimicifuga, representatives of the genus Actaea are the most frequently studied plants in the world of science. Nonetheless, little is known about the chemical profile and biological activity of other representatives of Cimicifugeae: Beesia and Anemonopsis. In recent decades, new information has been obtained about the chemical profiles of (and biological effects of extracts and individual compounds from) Eranthis species, which are early flowering geophytes with a limited geographic range.
2. Phytocomponents Identified in Eranthis Plants and Their Chemotaxonomic Significance
2.1. Chromones
Since the 1960s, from some Eranthis species, a series of substances has been isolated that represents an important class of oxygen-containing heterocyclic compounds that are derivatives of benzo-γ-pyrone: chromones. Their isolation has been performed by various chromatographic methods, and the structures of individual compounds have been investigated by 1-dimensional (H-NMR) and 2-dimensional nuclear magnetic resonance (C-NMR) spectroscopy. In structure, chromones are similar to flavonoids and coumarins but are substantially less common in the wild. Chromones can give rise to hydroxy- and methoxy-derivatives and can attach a sugar moiety, whereas after condensation with benzene, pyran, or furan rings, they can be transformed into a variety of benzo-, pyrano-, or furochromones, respectively. Compounds from the class “simple furochromones and chromones” are most often found in Eranthis species; chromones have been detected in underground parts, whereas furochromones have been found in underground and aboveground parts (Table 1).Table 1.
Chemical constituents of the genus
Eranthis
(all classes of metabolites identified to date: vertical subdivisions in the table).
| ID No. | Compound | Source | Eranthis | Species | Reference |
|---|---|---|---|---|---|
| Chromones | |||||
| 1 | 8,11-dihydro-5-hydroxy-2,9-dihydroxymethyl-4H-pyrano [2,3-g][1] benzoxepin-4-one |
E. cilicica (T *) | [21] | ||
| 2 | Eranthin (5-hydroxy-9-hydroxymethyl-2-methyl-8,11-dihydro-4H-pyrano[2,3-g][1]benzoxepin-4-one) |
E. hyemalis (R) | [22] | ||
| 3 | Eranthin-β-D-glucoside (9-{[(β-D-glucopyranosvl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) |
E. hyemalis (R, T) | [22][23][22,23] | ||
| 4 | Eranthin 9-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) E. hyemalis (T) |
[21][21[23],23] | ||
| 5 | Eranthin β-D-gentiobioside (9-{[(β-D-gentiobiosyl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) |
E. hyemalis (T) | [23] | ||
| 6 | 2-C-Hydroxyeranthin β-D-glucopyranoside (9-{[(β-D-glucopyranosyl)oxy]methyl}-8,11-dihydro-5-hydroxy-2-(hydroxymethyl-4H-pyrano[2,3-g][1]benzoxepin-4-one) |
E. hyemalis (T) | [23] | ||
| 7 | 9-[(O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl)oxy]methyl-8,11-dihydro-5,9-dihydroxy-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one | E. cilicica (T) | [21] | ||
| 8 | 8,11-dihydro-5,9-dihydroxy-9-hydroxymethyl-2-methyl-4H-pyrano[2,3-g][1]benzoxepin-4-one | E. cilicica (T) | [21] | ||
| 9 | 5,7-dihydroxy-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-2-methyl-4H-1-benzopyran-4-one | E. cilicica (T) | [21] | ||
| 10 | 5,7-dihydroxy-2-hydroxymethyl-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-4H-1-benzopyran-4-one | E. cilicica (T) | [21] | ||
| 11 | 7-[(β-D-glucopyranosyl)oxy]-5-hydroxy-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-2-methyl-4H-1-benzopyran-4-one | E. cilicica (T) | [21] | ||
| 12 | 7-[(β-D-glucopyranosyl)oxy]-5-hydroxy-2-hydroxymethyl-8-[(2E)-4-hydroxy-3-methylbut-2-enyl]-4H-1-benzopyran-4-one | E. cilicica (T) | [21] | ||
| 13 | 7,8-Secoeranthin β-D-glucoside (8-{(2E)-4-[(β-D-glucopyranosyl)oxy]-3-methylbut-2-enyl}-5,7-dihydroxy-2-methyl-4H-1-benzopyran-4-one) |
E. hyemalis (T) | [23] | ||
| 14 | 2-C-Hydroxy-7,8-secoeranthin β-D-glucoside (8-{(2E)-4-[(β-D-glucopyranosyl)oxy]-3-methylbut-2-enyl}-5,7-dihydroxy-2-(hydroxymethyl)-4H-1-benzopyran-4-one) |
E. hyemalis (T) | [23] | ||
| Furochromones | |||||
| 15 | Visnagin (4-methoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) |
E. hyemalis E. longistipitata (L) |
[24][25][24,25] | ||
| 16 | Khellin (4,9-dimethoxy-7-methyl-5H-furo[3,2-g]chromen-5-one) |
E. hyemalis E. longistipitata (L) |
[24][25][24,25] | ||
| 17 | Khellol (7-(hydroxymethyl)-4-methoxyfuro[3,2-g]chromen-5-one) |
E. pinnatifida (L, St) | [26] | ||
| 18 | Khellol glucoside (khellinin; 7-hydroxymethyl-4-methoxy-5H-furo [3,2-g]][1]benzopyran-5-one glucoside) |
E. hyemalis (L, F) | [27] | ||
| 19 | Norkhellol (4-hydroxy-7-(hydroxymethyl)-5H-furo[3,2-g][1]benzopyran-5-one) |
E. pinnatifida (L, St) | [26] | ||
| 20 | Norammiol (4-hydroxy-7(hydroxymethyl)-9-methoxy-5H-furo[3,2-g][1]-benzopyran-5-one) |
E. pinnatifida (L, St) | [26] | ||
| 21 | 7-[(O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranosyl)oxy]methyl-4-hydroxy-5H-furo[3,2-g][1]benzopyran-5-one | E. cilicica (T) | [21] | ||
| 22 | Methoxsalen (9-methoxyfuro[3,2-g]chromen-7-one) |
E. longistipitata (L) | [25] | ||
| 23 | Cimifugin (2S)-7-(hydroxymethyl)-2-(2-hydroxypropan-2-yl)-4-methoxy-2,3-dihydrofuro[3,2g]chromen-5-one) |
E. pinnatifida (L, St) E. cilicica (T) E. longistipitata (L) |
[21][25][26][21,25,26] | ||
| 24 | Norcimifugin (2S)-4-hydroxy-7-(hydroxymethyl)-2-(2-hydroxypropan-2-yl)-2,3-dihydrofuro[3,2-g]-chromen-5-one) |
E. pinnatifida (L, St) | [26] | ||
| 25 | Cimifugin β-D-glucopyranoside (7-{[(β-D-glucopyranosy1)oxy]methyl}-2,3-dihydro-2-(l-hydroxy-1-methylethyl)-4-methoxy-5H-furo[3,2-g][1]benzopyran-5-one) |
E. hyemalis (T) | [23] | ||
| 26 | 5-O-Methylvisammioside (4-O-β-D-glucosyl-5-O-methylvisamminol) |
E. longistipitata (L) | [25] | ||
| 27 | Visamminol-3′-O-glucoside (4-hydroxy-2-(2-hydroxypropan-2- yl)-methyl-2,3-dihydrofuro[3,2-g] chromen-5-one) |
E. longistipitata (L) | [25] | ||
| Triterpene saponins | |||||
| 28 | Eranthisaponin A | E. cilicica (T) | [28] | ||
| (3β-[(O-β-D-allopyranosyl-(1→3)-O-α-L-rhamnopyranosyl-(1→2)-O-[β-D-glucopyranosyl-(1→4)]-α-L-arabinopyranosyl)oxy]-23-hydroxyolean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl- (1→6)-β-D-glucopyranoside) |
|||||
| 29 | Eranthisaponin B (3β-[(O-β-D-glucopyranosyl-(1→4)-O-[α-L-rhamnopyranosyl-(1→2)]-α-L-arabinopyranosyl) oxy]-23-hydroxyolean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-O-β-D-glucopyranosyl-(1→4)-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl- (1→6)-β-D-glucopyranoside) |
E. cilicica (T) | [28] | ||
| 30 | 3β-[(O-β-D-glucopyranosyl-(1→4)-O-[α-L-rhamnopyranosyl-(1→2)]-α-L-arabinopyranosyl) oxy]-23-hydroxyolean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside |
E. cilicica (T) | [28] | ||
| 31 | 23-Hydroxy-3β-[(O-α-L-rhamnopyranosyl-(1→2)-α-L-arabinopyranosyl)oxy]olean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [28] | ||
| 32 | 3β-[(O-β-D-glucopyranosyl-(1→4)-α-L-arabinopyranosyl)oxy]-23-hydroxyolean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [28] | ||
| 33 | 3β-[(O-β-D-glucopyranosyl-(1→2)-O-[β-D-glucopyranosyl-(1→4)]-α-L-arabinopyranosyl)oxy]-23-hydroxyolean-12-en-28-oic acid 28-O-α-L-rhamnopyranosyl-(1→4)-O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [28] | ||
| 34 | 3β-[(O-β-D-glucopyranosyl-(1→4)-O-[α-L-rhamnopyranosyl-(1→2)]-α-L-arabinopyranosyl) oxy]-23-hydroxyolean-12-en-28-oic acid |
E. cilicica (T) | [29] | ||
| 35 | 3β-[(O-β-D-galactopyranosyl-(1→3)-O-α-L-rhamnopyranosyl-(1→2)-O-[β-D-glucopyranosyl-(1→4)]-α-L-arabinopyranosyl) oxy]-23-hydroxyolean-12-en-28-oic acid | E. cilicica (T) | [29] | ||
| 36 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cycloartan-3β-yl β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 37 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-9,19-cycloartane-3β,28-diol | E. cilicica (T) | [29] | ||
| 38 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cylcoartan-3β-yl O-β-D-glucopyranosyl-(1→4)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 39 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→3)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 40 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 41 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cylcoartan-3β-yl O-β-D-glucopyranosyl-(1→4)-O-[β-D-glucopyranosyl (1→6)]-β-D-glucopyranoside |
E. cilicica (T) | [29] | ||
| 42 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-28-oxo-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→4)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 43 | (23R,24R,25R)-16β,23:23,26:24,25-triepoxy-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 44 | (23S,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→4)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 45 | (23S,24R,25R)-16β,23:23,26:24,25-triepoxy-9,19-cycloartan-3β,28-diol | E. cilicica (T) | [29] | ||
| 46 | (23S,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cylcoartan-3β-yl O-β-D-glucopyranosyl-(1→3)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 47 | (23S,24R,25R)-16β,23:23,26:24,25-triepoxy-28-hydroxy-9,19-cylcoartan-3β-yl O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| 48 | (23S,24R,25R)-16β,23:23,26:24,25-triepoxy-9,19-cycloartan-3β-yl O-β-D-glucopyranosyl-(1→6)-β-D-glucopyranoside | E. cilicica (T) | [29] | ||
| Alkaloids | |||||
| 49 | Corytuberine (2,10-dimethoxy-6α-aporphine-1,11-diol) |
E. hyemalis (T; Ap) | [30] | ||
| Coumarins | |||||
| 50 | 5,7-Dihydroxy-4-methylcoumarin | E. longistipitata (L) | [25] | ||
| 51 | Scoparone (6,7-dimethoxycoumarin) |
E. longistipitata (L) | [25] | ||
| 52 | Fraxetin (7,8-dihydroxy-6-methoxycoumarin) |
E. longistipitata (L) | [25] | ||
| 53 | Luvangetin (10-methoxy-2,2-dimethylpyrano[3,2-g]chromen-8-one) |
E. longistipitata (L) | [25] | ||
| Flavonoids | |||||
| 54 | Quercetin | E. longistipitata (L) E. stellata (L) E. tanhoensis (L) |
[25][31][32][25,31,32] | ||
| 55 | Isoquercitrin (quercetin-3-O-β-D-glucoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 56 | Hyperoside (quercetin 3-O-β-D-galactoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 57 | Reynoutrin (quercetin-3-O-β-D-xylopyranoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 58 | Quercetin-6-O-β-D-xylopyranosyl-β-D- glucopyranoside |
E. longistipitata (L) | [25][32][25,32] | ||
| 59 | Quercetin-3-sambubioside (quercetin-3-O-[β-D-xylosyl-(1→2)-β-D-glucoside]) |
E. longistipitata (L) | [25][32][25,32] | ||
| 60 | Peltatoside (quercetin-3-(6-O-α-L-arabinopyranosyl)-β-D- glucopyranoside)) |
E. longistipitata (L) | [25][32][25,32] | ||
| 61 | Rutin (quercetin 3-O-β-D-rutinoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 62 | Kaempferol | E. longistipitata (L) E. stellata (L) E. tanhoensis (L) |
[25][31][32][25,31,32] | ||
| 63 | Juglalin (kaempferol 3-O-α-L-arabinopyranoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 64 | Trifolin (kaempferol-3-O-β-D-galactoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 65 | Aromadendrin [(+)-dihydrokaempferol] |
E. longistipitata (L) | [25][32][25,32] | ||
| 66 | Vitexin (apigenin 8-C-glucoside) |
E. sibirica (L) | [31] | ||
| 67 | Orientin (luteolin-8-C-glucoside) |
E. sibirica (L) | [31] | ||
| E. stellata (L) | |||||
| 68 | Carlinoside (luteolin 6-C-β-D-glucopyranoside-8-C-α-L- |
E. longistipitata (L) | [25][32][25,32] | ||
| arabinopyranoside) | |||||
| 69 | Cianidanol [(+)-catechin] |
E. longistipitata (L) | [25][32][25,32] | ||
| 70 | Auriculoside (7,3,5′-trihydroxy-4′-methoxyflavan-3′-glucoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 71 | 6-Methoxytaxifolin | E. longistipitata (L) | [25][32][25,32] | ||
| 72 | Aspalathin | E. longistipitata (L) | [25][32][25,32] | ||
| 73 | Phloridzin (phloretin-2′-O-β-glucoside) |
E. longistipitata (L) | [25][32][25,32] | ||
| 74 | Phloretin (dihydroxy naringenin) |
E. longistipitata (L) | [25][32][25,32] | ||
| Cinnamic acids | |||||
| 75 | Chlorogenic acid (3-O-caffeoylquinic acid) |
E. sibirica (L) E. stellata (L) E. tanhoensis (L) |
[31] | ||
| 76 | Caffeic acid (3,4-dihydroxycinnamic acid) |
E. sibirica (L) E. stellata (L) |
[31] | ||
| Phenolic acids | |||||
| 77 | Salicylic acid (3-tert-2-butyl-2-hydroxy-6-methylbenzoic acid) |
E. sibirica (L) E. tanhoensis (L) |
[31] | ||
| 78 | Gentisic acid (2,5-dihydroxybenzoic acid) |
E. stellata (L) | [31] | ||
| Fatty acids and their derivatives | |||||
| 79 | Myristic acid (14:0) | E. hyemalis (S) | [33] | ||
| 80 | Pentadecylic acid (15:0) | E. hyemalis (S) | [33] | ||
| 81 | Palmitic acid (16:0) | E. hyemalis (S) | [33] | ||
| 82 | 16-Hydroxyhexadecanoic acid | E. longistipitata (L) | [25] | ||
| 83 | cis-5-Hexadecenoic acid (16:1 Δ5cis) | E. hyemalis (S) | [33] | ||
| 84 | Palmitoleic acid (16:1 Δ9cis) | E. longistipitata (L) | [25] | ||
| 85 | cis-9-Octadecanoic acid (18:0 Δ9cis) | E. hyemalis (S) | [33] | ||
| 86 | cis-Vaccenic acid (18:1 Δ11cis) | E. hyemalis (S) | [33] | ||
| 87 | Linoleic acid (18:2 Δ9cis, 12cis) | E. hyemalis (S) | [33] | ||
| 88 | 9-oxo-ODA (9-Oxo-trans-10, trans-12-octadecadienoic acid) |
E. longistipitata (L) | [25] | ||
| 89 | (+/−)13-HODE | E. longistipitata (L) | [25] | ||
| (13-hydroxyoctadecadienoic acid) | |||||
| 90 | Corchorifatty acid F (9,12,13-trihydroxy-10(E),15(Z)-octadecadienoic acid) | E. longistipitata (L) | [25] | ||
| 91 | α-Linolenic acid (18:3 Δ9cis, 12cis, 15cis) | E. hyemalis (S) E. longistipitata (L) |
[25][25[33],33] | ||
| 92 | Linolenic acid ethyl ester | E. longistipitata (L) | [25] | ||
| 93 | α-Eleostearic acid (18:3 Δ9cis, 11trans, 13trans) | E. longistipitata (L) | [25] | ||
| 94 | Pinolenic acid (18:3 Δ5cis, 9cis, 12cis) | E. longistipitata (L) | [25] | ||
| 95 | 13(S)-HOTrE (13-OH-cis-9, trans-11, cis-15-octadecatrienoic acid) |
E. longistipitata (L) | [25] | ||
| 96 | (15Z)-9,12,13-Trihydroxy-15-octadecenoic acid | E. longistipitata (L) | [25] | ||
| 97 | 12-Oxo-phytodienoic acid | E. longistipitata (L) | [25] | ||
| 98 | 9S,13R-12-Oxo-phytodienoic acid | E. longistipitata (L) | [25] | ||
| 99 | Arachidic acid (20:0) | E. hyemalis (S) | [33] | ||
| 100 | cis-5-Eicosenoic acid (20:1 Δ5cis) | E. hyemalis (S) | [33] | ||
| 101 | Gondoic acid (20:1 Δ11cis) | E. hyemalis (S) | [33] | ||
| 102 | Keteleeronic acid (20:2 Δ5cis, 11cis) | E. hyemalis (S) | [33] | ||
| 103 | cis-11,14-Eicosadienoic acid (20:2 Δ11cis, 14cis) | E. hyemalis (S) | [33] | ||
| 104 | 15-OxoEDE (15-Oxo-cis-11,trans-13-eicosadienoic acid) |
E. longistipitata (L) | [25] | ||
| 105 | cis-5,11,14-Eicosatrienoic acid (20:3 Δ5cis, 11cis, 14cis) | E. hyemalis (S) | [33] | ||
| 106 | Behenic acid (22:0) | E. hyemalis (S) | [33] | ||
| 107 | Erucic acid (22:1 Δ13cis) | E. hyemalis (S) | [33] | ||
| 108 | cis-5,13-Docosadienoic acid (22:2 Δ5cis, 13cis) | E. hyemalis (S) | [33] | ||
| 109 | cis-13,16-Docosadienoic acid (22:2 Δ13cis, 16cis) | E. hyemalis (S) | [33] | ||
| 110 | cis-5,13,16-Docosatrienoic acid (22:3 Δ5cis, 13cis, 16cis) | E. hyemalis (S) | [33] | ||
| 111 | cis-10,13,16-Docosatrienoic acid (22:3 Δ10cis, 13cis, 16cis) | E. hyemalis (S) | [33] | ||
| Amino acids | |||||
| 112 | D-(+)-Pyroglutamic acid | E. longistipitata (L) | [25] | ||
| 113 | D-(+)-Tryptophan | E. longistipitata (L) | [25] | ||
| 114 | Isoleucine | E. longistipitata (L) | [25] | ||
| 115 | L-Phenylalanine | E. longistipitata (L) | [25] | ||
| 116 | L-Tyrosine | E. longistipitata (L) | [25] | ||
| 117 | D-(−)-Glutamine | E. longistipitata (L) | [25] | ||
| Organic acids | |||||
| 118 | Citric acid | E. longistipitata (L) | [25] | ||
| 119 | D-α-Hydroxyglutaric acid | E. longistipitata (L) | [25] | ||
| 120 | Gluconic acid | E. longistipitata (L) | [25] | ||
| Sugars | |||||
| 121 | α-Lactose | E. longistipitata (L) | [25] | ||
| 122 | D-(+)-Galactose | E. longistipitata (L) | [25] | ||
| 123 | α.α-Trehalose | E. longistipitata (L) | [25] | ||
| Alcohols | |||||
| 124 | D-(−)-Mannitol | E. longistipitata (L) | [25] | ||
| Phenylpropanoids | |||||
| 125 | 6-Gingerol | E. longistipitata (L) | [25] | ||
| Lectins | |||||
| 126 | EHL | E. hyemalis (R) | [34][35][34,35] | ||
* Ap, aerial part; F, flowers; L, leaves; R, rhizome; S, seeds; St, stems; T, tubers.
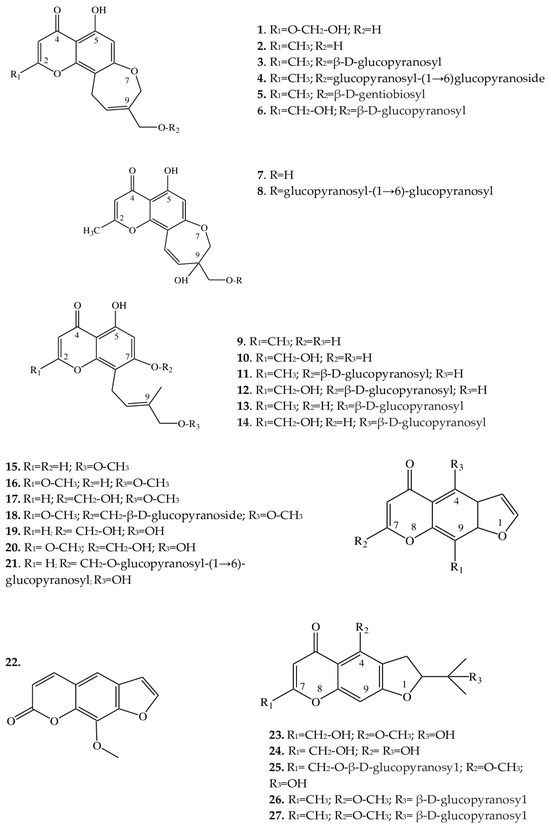
Figure 2.
Structures of furochromones and chromones from
Eranthis
species.
2.2. Furochromones
These compounds of Eranthis species are formed by the condensation of a simple chromone with a furan ring at positions C-6 and C-7 and, in contrast to the aforementioned chromones, are relatively common in the plant kingdom. For instance, the first representative of this subclass of compounds called khellin (16) has long been used in folk medicine to relieve ureteral pain during colic. For the first time, khellin was found in a seed extract of Ammi visnaga (L.) Lam. and was isolated as far back as the end of the 19th century [36]. Currently, khellin’s ability to act directly on smooth-muscle fibers is widely used in clinical practice [37]. Khellin, aside from species of the genus Ammi, has been found in other representatives of the family Apiaceae Lindl., for example, in Dioscorea L. sp. and Pimpinella L. sp. [38][39][40][38,39,40], and among Eranthis species, in E. hyemalis and E. longistipitata Regel [24][25][24,25]. The diversity of the structures in the furochromone subclass, which includes khellin, is mostly determined by the presence of substituents at the C-4, C-7, and C-9 positions. At the C-4 position, methoxy or hydroxyl groups can serve as a substituent; at position C-7, methoxy groups and glucose; and at position C-9, a methoxy group, or—as in 15, 17, 19, and 21—the substituent may be absent. Khellol (17) represents an aglycone of khellol glucoside (18), in which the sugar moiety is attached at position C-7. In the genus Eranthis, most research on furochromones of this subclass has been conducted on samples of the aerial parts (leaves, stems, and flowers) of E. pinnatifida Maxim., E. hyemalis, and E. longistipitata [24][25][26][27][24,25,26,27], and only compound 21 has been detected in an underground part (tubers) of E. cilicica [21]. Recently, new compounds not previously found in Eranthis species were discovered in samples of E. longistipitata from Central Asia (Kyrgyzstan): methoxsalen (22), 5-O-methylvisammioside (26), and visamminol-3′-O-glucoside (27) [25]. Methoxsalen (22) is often seen in the plant extracts of such families as Apiaceae, Rutaceae Juss., Fabaceae Lindl., and Brassicaceae Burnett [41][42][41,42], whereas the last two compounds of this subclass (26 and 27) have been registered only in extracts from an underground part of Saposhnikovia divaricata (Turcz.) Shischk. (Apiaceae) [43][44][45][43,44,45]. Because Eranthis species synthesize chromones during normal physiological processes, preliminary conclusions have been made that the genus Eranthis is closest to the genera Cimicifuga and Actaea (in whose extracts, chromones have also been found), and not Helleborus L. (for example), whose species do not synthesize chromones but are distinguished by the accumulation of cardenolides and bufadienolides [23][46][23,46].2.3. Triterpene Saponins
A phytochemical analysis of a methanol extract from tubers of E. cilicica has revealed two new bisdesmosidic triterpenes, named eranthisaponins A (28) and B (29) [28]. The new saponins are based on the structural backbone of hederagenin, which is a triterpenoid first isolated from seeds and leaves of Hedera helix L. [47]. A distinctive feature of eranthisaponin A (28) is a branched tetraglycoside attached at the C-3 position of the aglycone, whereas a feature of eranthisaponin B (29) is a linear hexaglycoside attached at the C-28 position of the aglycone. Such sugar forms in triterpene saponins have not been described previously. In addition, as one of the substituents, eranthisaponin A (28) contains D-allopyranose: a monosaccharide that is extremely rare in plant saponins [28]. Furthermore, in Eranthis plants, a number of known triterpene saponins (30–35) have been discovered that are (just as eranthisaponins B and A) based on the backbone of hederagenin with substituents at positions C-3 and C-28; extremely rarely (only in 31), the substituent (a hydroxyl group) is located at the C-23 position. Other substituents include di- and triglycosides composed of glucose, arabinose, and rhamnose residues. The research continued by K. Watanabe with coauthors [29] has allowed to subsequently isolate a new oleanane glycoside (34) from the tubers of E. cilicica. Another oleanane glycoside (35) had been discovered earlier in the underground part of Anemone coronaria L. (Ranunculaceae) [48]. These substances (34 and 35) contain triglycosides only at position C-3 of the carbohydrate part of the molecule. All of the above triterpene saponins belong to the oleanan type. In the same study [29], when fractionating a methanol extract from the tubers of E. cilicica, Watanabe et al. isolated several cycloartane-type compounds (36–48). There were 13 such triterpene saponins, all of which had not been characterized before. Compounds 37 and 45 are aglycones of 36 and 44, respectively. The new compounds can be categorized into two very similar subclasses: 36–43 and 44–48. In terms of their structure, rings A–D are similar, and differences lie in rings E and F (Figure 3).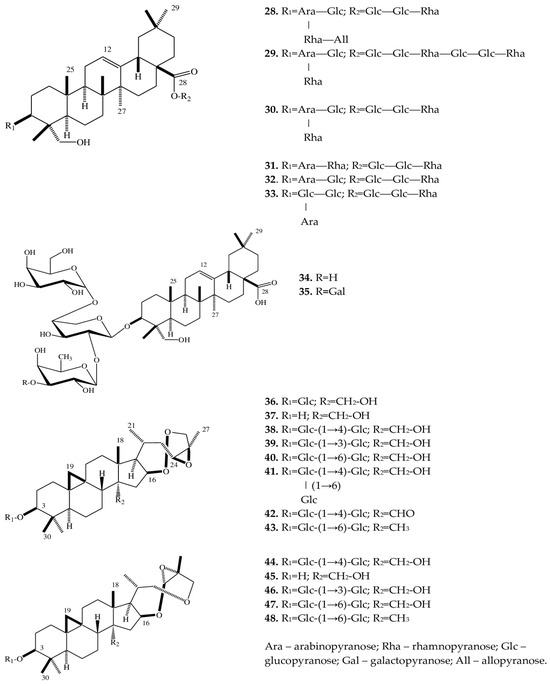
Figure 3.
Structures of triterpene saponins of
Eranthis
species.