The world is currently facing global energy crises and escalating environmental pollution, which are caused by the extensive exploitation of conventional energy sources. The limited availability of conventional energy sources has opened the door to the search for alternative energy sources. In this regard, microalgae have emerged as a promising substitute for conventional energy sources due to their high photosynthetic rate, high carbohydrate and lipid content, efficient CO2 fixation capacity, and ability to thrive in adverse environments. The research and development of microalgal-based biofuel as a clean and sustainable alternative energy source has been ongoing for many years, but it has not yet been widely adopted commercially. However, it is currently gaining greater attention due to the integrated biorefinery concept. This study provides an in-depth review of recent advances in microalgae cultivation techniques and explores methods for increasing lipid production by manipulating environmental factors. Furthermore, our discussions have covered high lipid content microalgal species, harvesting methods, biorefinery concepts, process optimizing software tools, and the accumulation of triglycerides in lipid droplets. The study additionally explores the influence of abiotic stresses on the response of biosynthetic genes involved in lipid synthesis and metabolism. In conclusion, algae-based biofuels offer a viable alternative to traditional fuels for meeting the growing demand for energy.
- microalgae
- cultivation systems
- biomass
- environmental stresses
- biorefinery
- biofuels
1. Introduction
2. Microalgal Species and Biorefinery for Sustainable Biofuel Feedstock
Microalgae are photoautotrophs with the capability to survive in freshwater and marine ecosystems. Currently, it is estimated that approximately 200,000 to 800,000 microalgal species are present on the earth, of which only 50,000 have been identified and characterized [29]. The lipid content of microalgae is an important factor in the selection and screening of microalgae for biofuel production [30]. Microalgae consume CO2 and fix it into carbohydrates, proteins, and lipids under different environmental conditions, playing an essential role in the production of a variety of renewable fuels [31][32]. Previous research has indicated that numerous algal species, including Chlorella vulgaris, Chlorella emersonii, Chlamydomonas reinhardtii, Nannochloropsis salina, Skeletonema sp., and Parachlorella kessleri, have significant lipid content [33][34][35][36][37]. However, some species, such as Chlorella pyrenoidosa, Chlorella zofingiensis, Chaetoceros muelleri, Coelastrella sp., and Chlorococcum pamirum, have relatively modest lipid content [38][39][40][41][42]. When considering the lipid content in various algal groups, a general trend can be observed: green algae > yellow-green algae > red algae > blue-green algae. Similarly, when examining lipid productivity, the trend follows: green algae > red algae [43]. Figure 1 shows different microalgal species with high lipid content that hold promise for potential biofuel production.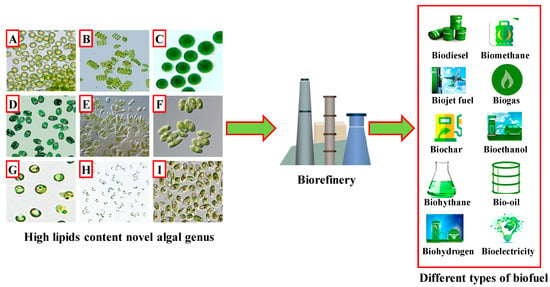
3. Microalgae Cultivation Systems
| Cultivation Mode | Carbon Source | Energy Supply | Light Availability | Advantages | Disadvantages |
|---|---|---|---|---|---|
| Autotrophic | Inorganic carbon | Light |
| Technique with Image | Principle | Advantages | Disadvantages | ||||
|---|---|---|---|---|---|---|---|
| Obligatory | Low cost, low energy consumption, high pigments production. | Low growth rate and biomass, specific photobioreactor required. | |||||
Flocculation |
Aggregation of cells is achieved by enlarging their size through the addition of a flocculant, which can be in the form of chemicals (such as ferric chloride, ferric sulfate, and ammonium sulfate) or microbes (bacteria). | Fast and easy technique, used for large scale, less cell damage, applied to a wide variety of species, less energy requirements. | Chemicals may be expensive, high pH required, separating the coagulant from harvested biomass is difficult, limited culture medium recycling, increased microbial contamination. |
||||
| Heterotrophic | Organic carbon | Organic | No requirement | High biomass productivity and lipid accumulation due to high growth rate, process of scaling up is simplified, organic substrates can be used to alter biomass composition. | Higher cost, easy to be contaminated by other microorganisms, only a few microalgal species that can grow in a heterotrophic environment, inability to synthesize metabolites triggered by light. |
||
Filtration |
Large cells (size > 70 µm) can be filtered under pressure or suction whereas smaller cells (size < 30 µm require ultrafilters to be harvested. | High recovery efficiency, cost effective, no chemical required, low energy consumption, low shear stress. | Slow hence requires pressure or vacuum, not effective for small algae, membrane fouling/clogging and replacement increases operational and maintenance costs. |
Mixotrophic | Inorganic and organic carbon |
Light and organic carbon |
No obligatory |
| Flotation | Increased growth rate, biomass, density and lipid accumulation, extended phase of exponential growth, stopping the photoinhibition effect and reducing biomass loss due to respiration during the dark hours, switch between photoautotroph and heterotroph regimens at any time. |  |
Trapping algal cells by bubbling air. | High cost, contamination problems, limited microalgae species will grow. | |||
| Well-suited for large-scale applications, economically efficient with minimal space demands, short operation time. | Depends on bubble distribution into the suspension, needs surfactants. | Algal turf scrubber | |||||
| Centrifugation | Organic and inorganic carbon |
Light and organic carbon |
Obligatory | Improved nutrient status, pollutant removal, high biomass productivity rate, easy harvesting and low maintaince, decreased the overall production cost | Requirement of sufficient space and infrastructure |
4. Microalgae Harvesting Techniques
Microalgae harvesting is the process of collecting microalgae from the liquid medium in which they are grown. The choice of microalgae harvesting method depends on a number of factors, including the properties of the microalgae, the desired properties of the end product, and the feasibility of recycling the growth medium. Microalgae cells can be harvested using biological, chemical, mechanical, or electrical methods. Microalgae harvesting is a time-consuming and challenging process that often requires the use of advanced chemical or mechanical techniques. Flocculation, filtration, flotation, sonication, centrifugation, and precipitation are some of the methods that are used to harvest microalgal biomass (Table 2). When choosing a harvesting method, it is important to consider the downstream processing requirements, as microalgal biomass will need to be processed further. Therefore, these processes must be carefully operated to avoid damaging or contaminating the microalgal biomass. It is also ideal if the chosen harvesting method permits the culture medium to be recycled [50]. Often, a combination of two or more harvesting methods is used to achieve greater separation efficiency and minimize costs | |||
| Sedimentation based on the velocity, cell size and density. | |||
| Fast and effective technique, high recovery efficiency (>90), applicable to all microalgae. | Expensive technique with high energy requirement, high operation and maintenance costs, risk of cell destruction. | ||
Precipitation |
Certain algae undergo self-precipitation, they settle at the bottom when circulation is halted. | No energy or chemicals are needed, it occurs naturally. | Species-specific, time periods vary depending on the species, not every species is self-precipitated. |
5. Approaches to Stimulate Lipid Production through Abiotic Stresses
5.1. Effect of Carbon Dioxide
Microalgae can mitigate CO2 emissions by CO2 biofixing and can grow well in high CO2 concentrations [20]. The growth rate and lipid content of microalgae can be affected by the concentration of carbon dioxide. Many studies have investigated the effects of CO2 on the growth and fatty acid composition of microalgal strains. The mechanism involves the uptake of CO2 by algae and converting the captured carbon into lipids. The carbon from microalgae is then converted into biodiesel through transesterification, as shown in Figure 2.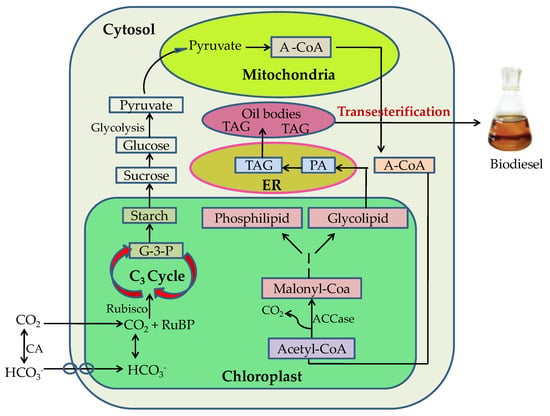
5.2. Effect of Nitrogen Starvation
5.3. Effect of Light Intensity and Wavelength
Light is one of the most important environmental factors for microalgal growth. Previous studies have shown that high light intensity can significantly increase lipid accumulation in microalgae. The TAG content increased and the polar phospholipid composition decreased as the light intensity was increased in Cladophora sp. [64]. Nannochloropsis sp. cultivated under low light intensity (35 µmol m−2 s−1) had a lipid composition of 26% TAG and 40% galactolipid. A unicellular alga, Scenedesmus sp., accumulates a significant amount of lipid content when cultivated under light conditions of 250–400 µmol m−2 s−1 [65]. The total lipid content of Dunaliella salina, Isochrysis galbana, and Nannochloropsis oculata increases when the light intensity is increased to 150 µmol m−2 s−1 [66]. Microalgae grown under red light showed higher lipid accumulation than those grown under yellow or white light. Microalgae cultivated under green light had lower lipid accumulation than those grown under red light. For example, exposing a unicellular microalga Chlorella sp. to red light resulted in increased growth and lipid content [67]. Another technique for improving microalgal growth and lipid accumulation is to use chemical dyes to change the light spectrum [68]. For example, organic dyes such as rhodamine 101 and 9,10-diphenylanthracene have been used to convert unused and/or harmful portions of incident sunlight into usable photons for microalgae cultivation, thereby increasing the productivity of biomass and lipids. The lipid yield of Chlorella vulgaris can be improved by cultivating it in a medium containing these chemical dyes [68]. Red paint produced the highest biomass output, while blue paint produced the maximum lipid content of 30% dry weight [69]. The lipid composition of microalgae is significantly affected by photoperiod. The effect of light intensity on the lipid composition of microalgae has been investigated in a variety of ways, and it has been found that PUFA levels decrease as light intensity increases [70]. Microalgae grown under high light intensity and long light periods had higher saturated fatty acids (SFAs) levels and lower MUFAs and PUFAs levels [71].5.4. Effect of Temperature
Temperature changes influence not only lipid synthesis but also lipid content in the microalgae. In most microalgae, a decrease in temperature resulted in an increase in polar lipid content, while an increase in temperature resulted in an accumulation of non-polar lipids [72]. Exposing Acutodesmus dimorphus to temperature stress at 35 °C led to a 22.7% increase in lipid yield and a 59.9% increase in neutral lipid accumulation [73]. A decrease in temperature from 30 to 25 °C in Chlorella vulgaris has been shown to increase lipid content by 2.5-fold, without any change in growth rate [74]. Low temperatures simultaneously stimulate lipid content and reduce growth in Chlorella sorokiniana [75]. Although most studies focus on total lipid content, few have investigated the impact of temperature on specific lipid classes. Microalgae cultivated at low temperatures have a high content of PUFAs, which are important for survival in harsh conditions. When the temperature was decreased from 25 to 10 °C, Isochrysis galbana showed a significant increase in PUFA content [76]. Phaeodactylum tricornutum also showed a similar increase in PUFA content when the temperature was decreased from 25 to 10 °C [77].5.5. Effect of Salinity
In response to salt stress, microalgae typically undergo biochemical changes that regulate lipid production. However, the salinity tolerance of different microalgae strains varies. Dunaliella sp. is an example of a microalga that can withstand high salt concentrations. Dunaliella salina is a well-studied microalga because it can use salt stress to increase both biomass productivity and lipid yield. Dunaliella tertiolecta exhibited an increase in both total lipid content and a notable percentage of TAGs when the amount of NaCl was increased [78]. Scenedesmus sp., when subjected to salinity stress of 400 mM during a biphasic cultivation process, resulted in a lipid content of 34.77% [79]. The lipid content of various microalgae, including Scenedesmus sp., Chlorella vulgaris, and Chlamydomonas mexicana, increases under salt stress conditions [19][79][80]. In most microalgae, the lipid content peaks at a certain salt concentration, and then declines significantly beyond this concentration. In Chlorella minutissima, salt stress affects both biomass production and lipid content [81].5.6. Effect of pH
The pH affects the photosynthetic process, the solubility of inorganic nutrients, the rate of lipid accumulation, and the activity of enzymes in the cell. For microalgal growth, a certain pH range is required, which is limited and strain-specific. Microalgae are able to accumulate higher lipid content within a pH range of 7 to 9.5. Chlorella sp. showed an increase in lipid content up to 23% at pH 8, and Tetraselmis suecica showed a similar trend at pH 7.5. A study of the impact of various pH values (6, 7, 8, 9, and 10) on the biomass and lipid content of Nannochloropsis salina found that N. salina exhibited optimal growth rates and lipid content at pH 8 and 9 [82]. The relationship between CO2 and pH for microalgal cultivation is intricately linked through a process known as carbon dioxide dissolution or carbonic acid equilibrium. pH is important for microalgal growth because it affects the solubility of CO2 in the culture medium. As the pH increases, the solubility of CO2 decreases. This means that there is less CO2 available for microalgae to use. The optimal pH range for microalgal growth varies depending on the species of microalgae. However, most microalgae grow best in a pH range of 6.5 to 8.5 [24]. At pH levels below 6.5, the solubility of CO2 is too low, and at pH levels above 8.5, the toxicity of CO2 increases. When CO2 dissolves in water, it forms carbonic acid (H2CO3). The relationship between CO2 and pH is complex. As microalgae grow, they take up CO2 from the medium, which causes the pH to increase. This can be counteracted by injecting CO2 into the culture, or by adding a buffer to the medium. The amount of CO2 that needs to be added or the amount of buffer that needs to be added depends on the pH of the medium, the species of microalgae, and the growth rate of the culture. As CO2 dissolves in the culture medium where microalgae are cultivated, it decreases the pH of the medium. This means that as more CO2 is added to the culture, the pH of the medium becomes more acidic due to the formation of carbonic acid. Conversely, when CO2 is removed from the medium (for instance, through microalgal photosynthesis), the pH tends to increase, becoming more alkaline. This pH fluctuation is essential to monitor and control in microalgal cultivation systems because microalgae have specific pH preferences for optimal growth.5.7. Effect of Metal
The lipid content of microalgae is affected by the presence of trace metals, and the amount of lipids produced depends on the concentration of these metals in the growth medium. Heavy metals, including copper, zinc, and cadmium has been shown to increase lipid yield in some microalgae. Iron stands out as particularly effective in enhancing photosynthetic enzyme activities. For example, the overall lipid content of Chlorella vulgaris increases by approximately 3 to 7 times when it is exposed to FeCl3 [83].5.8. Effect of Sulfur Starvation
Algae require sulfur in the form of sulfate as an essential nutrient for their growth. Sulfur limitation in the growth medium can induce lipid accumulation in algae. In response to sulfur limitation, algae may alter their metabolic pathways to prioritize lipid synthesis as a way to cope with the stress. For example, under sulfur starvation, Chromochloris zofngiensis and Scenedesmus acuminatus exhibit high lipid accumulation [84][85]. Sulfur stress can also change the fatty acid profile of algal lipids. This could lead to changes in the proportion of saturated to unsaturated fatty acids and the length of fatty acid chains [85]. Sulfur stress can alter the regulation of key enzymes involved in lipid biosynthesis pathways.5.9. Effect of Phytohormones
To stimulate lipid accumulation in microalgae, it is important to use both traditional abiotic stress conditions and innovative techniques. This is necessary because there is a reciprocal relationship between biomass and lipid accumulation under abiotic stress [86]. The effects of phytohormones on microalgal metabolism, especially in relation to lipid production, are still not fully clear. However, it has been noted that auxins, a type of phytohormone, can promote the growth of Scenedesmus sp. This growth regulator has been found to increase the content of TAGs and MUFAs, while simultaneously reducing the content of PUFAs [87]. The combination of indole-3-butyric acid (IBA) at a level of 10 mg/L and 6-benzylaminopurine (BAP) at a level of 5 mg/L exhibited a synergistic effect. The synergistic effect led to a 2.34 g/L increase in biomass production and a 42.43% increase in lipid content [88]. Similar findings were reported in Chlorella sp., where the application of IBA promoted maximum growth and lipid yield [89]. Fluvic acid, an additional phytohormone, has been used to enhance lipid yield in microalgae. This effect was achieved by modulating gene expression and the actions of principal enzymes such as phosphoenolpyruvate carboxylase and acetyl-CoA carboxylase [90]. The application of salicylic acid (SA) at a concentration of 10 ppm resulted in a substantial increase in lipid, reaching 475 mg/L in the early stationary growth phase. The application of salicylic acid (SA) was found to be crucial for the formation of omega-3 fatty acids, specifically eicosapentaenoic acid (EPA, C20:5) [91]. Higher concentrations of methyl jasmonate (MeJA) were found to promote the formation of MUFAs, especially oleic acid (C18:1). Most phytohormones have demonstrated the ability to enhance biomass production, and some have also shown promise in improving lipid yield and modifying lipid composition.6. Microalgal Biorefinery for Biofuels Production
A microalgal biorefinery is a specialized facility designed to harness the full potential of microalgae for the production of biofuels and various other valuable products. Microalgal biomass can be processed into an extensive array of products, including proteins, carbohydrates, lipids, pigments, polyunsaturated fatty acids, antioxidants, nutraceuticals, vitamins, biofertilizers, animal feed, biosurfactants, and bioenergy products (Figure 3). A microalgal biorefinery, in combination with other processes, has the potential to solve the current bioeconomic problem by producing multiple high-value products [92]. Microalgae are an efficient feedstock for generating various types of biofuels, depending on the chosen generation route (Figure 3). Microbial biomass conversion to biofuels is an alternative approach to the increasing demand for fossil fuels. Microalgal biorefineries hold promise as a sustainable and versatile solution for biofuel production, offering the potential to address energy needs while promoting environmental sustainability and economic diversification [93][94].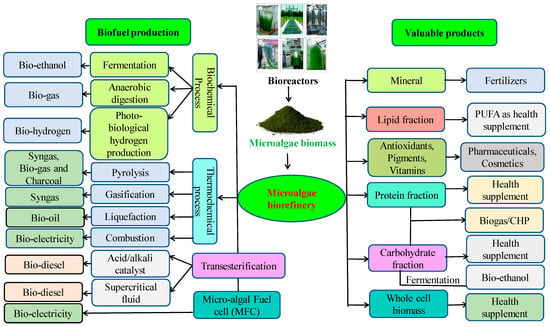
References
- Patil, P.D.; Gude, V.G.; Mannarswamy, A.; Cooke, P.; Nirmalakhandan, N.; Lammers, P.; Deng, S. Comparison of direct transesterification of algal biomass under supercritical methanol and microwave irradiation conditions. Fuel 2012, 97, 822–831.
- Ahamed, T.S.; Brindhadevi, K.; Krishnan, R.; Phuong, T.N.; Alharbi, S.A.; Chinnathambi, A.; Mathimani, T. In vivo detection of triacylglycerols through Nile red staining and quantification of fatty acids in hyper lipid producer Nannochloropsis sp. cultured under adequate nitrogen and deficient nitrogen condition. Fuel 2022, 322, 124179.
- Jeyakumar, N.; Hoang, A.T.; Nižetić, S.; Balasubramanian, D.; Kamaraj, S.; Pandian, P.L.; Sirohi, R.; Nguyen, P.Q.P.; Nguyen, X.P. Experimental investigation on simultaneous production of bioethanol and biodiesel from macro-algae. Fuel 2022, 329, 125362.
- Bhatia, S.K.; Ahuja, V.; Chandel, N.; Gurav, R.; Bhatia, R.K.; Govarthanan, M.; Tyagi, V.K.; Kumar, V.; Pugazendhi, A.; Banu, J.R.; et al. Advances in algal biomass pretreatment and its valorisation into biochemical and bioenergy by the microbial processes. Bioresour. Technol. 2022, 358, 127437.
- Singh, R.P.; Yadav, P.; Kumar, A.; Hashem, A.; Al-Arjani, A.B.F.; Abd_Allah, E.F.; Rodríguez Dorantes, A.; Gupta, R.K. Physiological and biochemical responses of bicarbonate supplementation on biomass and lipid content of green algae Scenedesmus sp. BHU1 isolated from wastewater for renewable biofuel feedstock. Front. Microbiol. 2022, 13, 839800.
- Tiwari, O.N.; Bhunia, B.; Bandyopadhyay, T.K.; Oinam, G. Strategies for improved induction of lipid in Leptolyngbya sp. BTA 287 for biodiesel production. Fuel 2019, 256, 115896.
- Ranganathan, P.; Pandey, A.K.; Sirohi, R.; Hoang, A.T.; Kim, S.H. Recent advances in computational fluid dynamics (CFD) modelling of photobioreactors: Design and applications. Bioresour. Technol. 2022, 350, 126920.
- Mehmood, M.A.; Shahid, A.; Malik, S.; Wang, N.; Javed, M.R.; Haider, M.N.; Verma, P.; Ashraf, M.U.F.; Habib, N.; Syafiuddin, A.; et al. Advances in developing metabolically engineered microbial platforms to produce fourth-generation biofuels and high-value biochemicals. Bioresour. Technol. 2021, 337, 125510.
- Anto, S.; Karpagam, R.; Renukadevi, P.; Jawaharraj, K.; Varalakshmi, P. Biomass enhancement and bioconversion of brown marine microalgal lipid using heterogeneous catalysts mediated transesterification from biowaste derived biochar and bio nanoparticle. Fuel 2019, 255, 115789.
- Japar, A.S.; Takriff, M.S.; Yasin, N.H.M. Microalgae acclimatization in industrial wastewater and its effect on growth and primary metabolite composition. Algal Res. 2021, 53, 102163.
- Bibi, F.; Ali, M.I.; Ahmad, M.; Bokhari, A.; Khoo, K.S.; Zafar, M.; Asif, S.; Mubashir, M.; Han, N.; Show, P.L. Production of lipids biosynthesis from Tetradesmus nygaardii microalgae as a feedstock for biodiesel production. Fuel 2022, 326, 124985.
- Shuba, E.S.; Kifle, D. Microalgae to biofuels: ‘Promising’alternative and renewable energy, review. Renew. Sustain. Energy Rev. 2018, 81, 743–755.
- Ali, S.S.; Mastropetros, S.G.; Schagerl, M.; Sakarika, M.; Elsamahy, T.; El-Sheekh, M.; Sun, J.; Kornaros, M. Recent advances in wastewater microalgae-based biofuels production: A state-of-the-art review. Energy Rep. 2022, 8, 13253–13280.
- Saengsawang, B.; Bhuyar, P.; Manmai, N.; Ponnusamy, V.K.; Ramaraj, R.; Unpaprom, Y. The optimization of oil extraction from macroalgae, Rhizoclonium sp. by chemical methods for efficient conversion into biodiesel. Fuel 2020, 274, 117841.
- Bhatia, S.K.; Mehariya, S.; Bhatia, R.K.; Kumar, M.; Pugazhendhi, A.; Awasthi, M.K.; Atabani, A.E.; Kumar, G.; Kim, W.; Seo, S.O.; et al. Wastewater based microalgal biorefinery for bioenergy production: Progress and challenges. Sci. Total Environ. 2021, 751, 141599.
- Patel, P.; Patel, B.; Vekaria, E.; Shah, M. Biophysical economics and management of biodiesel, a harbinger of clean and sustainable energy. Int. J. Energy Water Resour. 2020, 4, 411–423.
- Goswami, R.K.; Mehariya, S.; Obulisamy, P.K.; Verma, P. Advanced microalgae-based renewable biohydrogen production systems: A review. Bioresour. Technol. 2021, 320, 124301.
- Mehariya, S.; Goswami, R.K.; Verma, P.; Lavecchia, R.; Zuorro, A. Integrated approach for wastewater treatment and biofuel production in microalgae biorefineries. Energies 2021, 14, 2282.
- Salama, E.S.; Abou-Shanab, R.A.; Kim, J.R.; Lee, S.; Kim, S.H.; Oh, S.E.; Kim, H.C.; Roh, H.S.; Jeon, B.H. The effects of salinity on the growth and biochemical properties of Chlamydomonas mexicana GU732420 cultivated in municipal wastewater. Environ. Technol. Innov. 2014, 35, 1491–1498.
- Singh, S.P.; Singh, P. Effect of CO2 concentration on algal growth: A review. Renew. Sustain. Energy Rev. 2014, 38, 172–179.
- Liu, J.; Huang, J.; Sun, Z.; Zhong, Y.; Jiang, Y.; Chen, F. Differential lipid and fatty acid profiles of photoautotrophic and heterotrophic Chlorella zofingiensis: Assessment of algal oils for biodiesel production. Bioresour. Technol. 2011, 102, 106–110.
- Mathimani, T.; Uma, L.; Prabaharan, D. Formulation of low-cost seawater medium for high cell density and high lipid content of Chlorella vulgaris BDUG 91771 using central composite design in biodiesel perspective. J. Clean. Prod. 2018, 198, 575–586.
- Yadav, P.; Singh, R.P.; Alodaini, H.A.; Hatamleh, A.A.; Santoyo, G.; Kumar, A.; Gupta, R.K. Impact of dehydration on the physiochemical properties of Nostoc calcicola BOT1 and its untargeted metabolic profiling through UHPLC-HRMS. Front. Plant Sci. 2023, 14, 1147390.
- Cheng, D.; He, Q. Assessment of environmental stresses for enhanced microalgal biofuel production–an overview. Front. Energy Res. 2014, 2, 26.
- Miranda, A.M.; Hernandez-Tenorio, F.; Ocampo, D.; Vargas, G.J.; Sáez, A.A. Trends on CO2 capture with microalgae: A bibliometric analysis. Molecules 2022, 27, 4669.
- Park, S.; Nguyen, T.H.T.; Jin, E. Improving lipid production by strain development in microalgae: Strategies, challenges and perspectives. Bioresour. Technol. 2019, 292, 121953.
- Rastogi, R.P.; Pandey, A.; Larroche, C.; Madamwar, D. Algal Green Energy–R&D and technological perspectives for biodiesel production. Renew. Sustain. Energy Rev. 2018, 82, 2946–2969.
- Seo, J.Y.; Jeon, H.J.; Kim, J.W.; Lee, J.; Oh, Y.K.; Ahn, C.W.; Lee, J.W. Simulated-sunlight-driven cell lysis of magnetophoretically separated microalgae using ZnFe2O4 octahedrons. Ind. Eng. Chem. Res. 2018, 57, 1655–1661.
- Yin, Z.; Zhu, L.; Li, S.; Hu, T.; Chu, R.; Mo, F.; Hu, D.; Liu, C.; Li, B. A comprehensive review on cultivation and harvesting of microalgae for biodiesel production: Environmental pollution control and future directions. Bioresour. Technol. 2020, 301, 122804.
- Neofotis, P.; Huang, A.; Sury, K.; Chang, W.; Joseph, F.; Gabr, A.; Twary, S.; Qiu, W.; Holguin, O.; Polle, J.E. Characterization and classification of highly productive microalgae strains discovered for biofuel and bioproduct generation. Algal Res. 2016, 15, 164–178.
- Khandelwal, A.; Vijay, A.; Dixit, A.; Chhabra, M. Microbial fuel cell powered by lipid extracted algae: A promising system for algal lipids and power generation. Bioresour. Technol. 2018, 247, 520–527.
- Song, D.; Park, J.; Kim, K.; Lee, L.S.; Seo, J.Y.; Oh, Y.K.; Kim, Y.J.; Ryou, M.H.; Lee, Y.M.; Lee, K. Recycling oil-extracted microalgal biomass residues into nano/micro hierarchical Sn/C composite anode materials for lithium-ion batteries. Electrochim. Acta 2017, 250, 59–67.
- Beacham, T.A.; Macia, V.M.; Rooks, P.; White, D.A.; Ali, S.T. Altered lipid accumulation in Nannochloropsis salina CCAP849/3 following EMS and UV induced mutagenesis. Biotechnol. Rep. 2015, 7, 87–94.
- Deng, X.Y.; Gao, K.; Addy, M.; Li, D.; Zhang, R.C.; Lu, Q.; Ma, Y.W.; Cheng, Y.L.; Chen, P.; Liu, Y.H.; et al. Cultivation of Chlorella vulgaris on anaerobically digested swine manure with daily recycling of the post-harvest culture broth. Bioresour. Technol. 2018, 247, 716–723.
- Shin, Y.S.; Jeong, J.; Nguyen, T.H.T.; Kim, J.Y.H.; Jin, E.; Sim, S.J. Targeted knockout of phospholipase A2 to increase lipid productivity in Chlamydomonas reinhardtii for biodiesel production. Bioresour. Technol. 2019, 271, 368–374.
- Takeshita, T.; Ivanov, I.N.; Oshima, K.; Ishii, K.; Kawamoto, H.; Ota, S.; Yamazaki, T.; Hirata, A.; Kazama, Y.; Abe, T.; et al. Comparison of lipid productivity of Parachlorella kessleri heavy-ion beam irradiation mutant PK4 in laboratory and 150-L mass bioreactor, identification and characterization of its genetic variation. Algal Res. 2018, 35, 416–426.
- Yu, N.; Dieu, L.T.J.; Harvey, S.; Lee, D.Y. Optimization of process configuration and strain selection for microalgae-based biodiesel production. Bioresour. Technol. 2015, 193, 25–34.
- Feng, P.; Deng, Z.; Hu, Z.; Wang, Z.; Fan, L. Characterization of Chlorococcum pamirum as a potential biodiesel feedstock. Bioresour. Technol. 2014, 162, 115–122.
- Gao, Y.; Yang, M.; Wang, C. Nutrient deprivation enhances lipid content in marine microalgae. Bioresour. Technol. 2013, 147, 484–491.
- Karpagam, R.; Raj, K.J.; Ashokkumar, B.; Varalakshmi, P. Characterization and fatty acid profiling in two fresh water microalgae for biodiesel production: Lipid enhancement methods and media optimization using response surface methodology. Bioresour. Technol. 2015, 188, 177–184.
- Mao, X.; Wu, T.; Sun, D.; Zhang, Z.; Chen, F. Differential responses of the green microalga Chlorella zofingiensis to the starvation of various nutrients for oil and astaxanthin production. Bioresour. Technol. 2018, 249, 791–798.
- Wen, X.; Geng, Y.; Li, Y. Enhanced lipid production in Chlorella pyrenoidosa by continuous culture. Bioresour. Technol. 2014, 161, 297–303.
- Maity, J.P.; Bundschuh, J.; Chen, C.Y.; Bhattacharya, P. Microalgae for third generation biofuel production, mitigation of greenhouse gas emissions and wastewater treatment: Present and future perspectives—A mini review. Energy 2014, 78, 104–113.
- Wiencke, C.; Clayton, M.N.; Gómez, I.; Iken, K.; Lüder, U.H.; Amsler, C.D.; Karsten, U.; Hanelt, D.; Bischof, K.; Dunton, K. Life strategy, ecophysiology and ecology of seaweeds in polar waters. Rev. Environ. Sci. Biotechnol. 2007, 6, 95–126.
- Kuo, C.M.; Lin, T.H.; Yang, Y.C.; Zhang, W.X.; Lai, J.T.; Wu, H.T.; Chang, J.S.; Lin, C.S. Ability of an alkali-tolerant mutant strain of the microalga Chlorella sp. AT1 to capture carbon dioxide for increasing carbon dioxide utilization efficiency. Bioresour. Technol. 2017, 244, 243–251.
- Abid, A.; Saidane, F.; Hamdi, M. Feasibility of carbon dioxide sequestration by Spongiochloris sp. microalgae during petroleum wastewater treatment in airlift bioreactor. Bioresour. Technol. 2017, 234, 297–302.
- Choi, Y.Y.; Patel, A.K.; Hong, M.E.; Chang, W.S.; Sim, S.J. Microalgae Bioenergy with Carbon Capture and Storage (BECCS): An emerging sustainable bioprocess for reduced CO2 emission and biofuel production. Bioresour. Technol. Rep. 2019, 7, 100270.
- Araújo, R.; Vázquez Calderón, F.; Sánchez López, J.; Azevedo, I.C.; Bruhn, A.; Fluch, S.; Garcia Tasende, M.; Ghaderiardakani, F.; Ilmjärv, T.; Laurans, M.; et al. Current status of the algae production industry in Europe: An emerging sector of the blue bioeconomy. Front. Mar. Sci. 2021, 7, 626389.
- Zhan, J.; Rong, J.; Wang, Q. Mixotrophic cultivation, a preferable microalgae cultivation mode for biomass/bioenergy production, and bioremediation, advances and prospect. Int. J. Hydrogen Energy 2017, 42, 8505–8517.
- Uduman, N.; Qi, Y.; Danquah, M.K.; Forde, G.M.; Hoadley, A. Dewatering of microalgal cultures: A major bottleneck to algae-based fuels. J. Renew. Sustain. Energy 2010, 2, 012701.
- Barros, A.I.; Gonçalves, A.L.; Simões, M.; Pires, J.C. Harvesting techniques applied to microalgae: A review. Renew. Sustain. Energy Rev. 2015, 41, 1489–1500.
- Saad, M.G.; Dosoky, N.S.; Zoromba, M.S.; Shafik, H.M. Algal biofuels: Current status and key challenges. Energies 2019, 12, 1920.
- Raeesossadati, M.J.; Ahmadzadeh, H.; McHenry, M.P.; Moheimani, N.R. CO2 bioremediation by microalgae in photobioreactors: Impacts of biomass and CO2 concentrations, light, and temperature. Algal Res. 2014, 6, 78–85.
- Lv, J.M.; Cheng, L.H.; Xu, X.H.; Zhang, L.; Chen, H.L. Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresour. Technol. 2010, 101, 6797–6804.
- Heredia-Arroyo, T.; Wei, W.; Hu, B. Oil accumulation via heterotrophic/mixotrophic Chlorella protothecoides. Appl. Biochem. Biotechnol. 2010, 162, 1978–1995.
- Ferreira, G.F.; Pinto, L.R.; Maciel Filho, R.; Fregolente, L.V. A review on lipid production from microalgae: Association between cultivation using waste streams and fatty acid profiles. Renew. Sustain. Energy Rev. 2019, 109, 448–466.
- Nascimento, I.A.; Cabanelas, I.T.D.; dos Santos, J.N.; Nascimento, M.A.; Sousa, L.; Sansone, G. Biodiesel yields and fuel quality as criteria for algal-feedstock selection: Effects of CO2-supplementation and nutrient levels in cultures. Algal Res. 2015, 8, 53–60.
- Bibi, F.; Jamal, A.; Huang, Z.; Urynowicz, M.; Ali, M.I. Advancement and role of abiotic stresses in microalgae biorefinery with a focus on lipid production. Fuel 2022, 316, 123192.
- Sulochana, S.B.; Arumugam, M. Targeted metabolomic and biochemical changes during nitrogen stress mediated lipid accumulation in Scenedesmus quadricauda CASA CC202. Front. Bioeng. Biotechnol. 2020, 8, 585632.
- Chen, H.; Wang, Q. Regulatory mechanisms of lipid biosynthesis in microalgae. Biol. Rev. 2021, 96, 2373–2391.
- Shtaida, N.; Khozin, G.I.; Boussiba, S. The role of pyruvate hub enzymes in supplying carbon precursors for fatty acid synthesis in photosynthetic microalgae. Photosynth. Res. 2015, 125, 407–422.
- Nagappan, S.; Devendran, S.; Tsai, P.C.; Jayaraman, H.; Alagarsamy, V.; Pugazhendhi, A.; Ponnusamy, V.K. Metabolomics integrated with transcriptomics and proteomics: Evaluation of systems reaction to nitrogen deficiency stress in microalgae. Process Biochem. 2020, 91, 1–14.
- Liu, T.; Chen, Z.; Xiao, Y.; Yuan, M.; Zhou, C.; Liu, G.; Fang, J.; Yang, B. Biochemical and morphological changes triggered by nitrogen stress in the oleaginous microalga Chlorella vulgaris. Microorganisms 2022, 10, 566.
- Napolitano, G.E. The relationship of lipids with light and chlorophyll measurements in freshwater algae and periphyton 1. J. Phycol. 1994, 30, 943–950.
- Gris, B.; Morosinotto, T.; Giacometti, G.M.; Bertucco, A.; Sforza, E. Cultivation of Scenedesmus obliquus in photobioreactors: Effects of light intensities and light–dark cycles on growth, productivity, and biochemical composition. Appl. Biochem. Biotechnol. 2014, 172, 2377–2389.
- Gim, G.H.; Ryu, J.; Kim, M.J.; Kim, P.I.; Kim, S.W. Effects of carbon source and light intensity on the growth and total lipid production of three microalgae under different culture conditions. J. Ind. Microbiol. 2016, 43, 605–616.
- Severes, A.; Hegde, S.; D’Souza, L.; Hegde, S. Use of light emitting diodes (LEDs) for enhanced lipid production in micro-algae-based biofuels. J. Photochem. Photobiol. B Biol. 2017, 170, 235–240.
- Seo, Y.H.; Lee, Y.; Jeon, D.Y.; Han, J.I. Enhancing the light utilization efficiency of microalgae using organic dyes. Bioresour. Technol. 2015, 181, 355–359.
- Seo, Y.H.; Cho, C.; Lee, J.Y.; Han, J.I. Enhancement of growth and lipid production from microalgae using fluorescent paint under the solar radiation. Bioresour. Technol. 2014, 173, 193–197.
- Juneja, A.; Ceballos, R.M.; Murthy, G.S. Effects of environmental factors and nutrient availability on the biochemical composition of algae for biofuels production: A review. Energies 2013, 6, 4607–4638.
- Al-Qasmi, M.; Raut, N.; Talebi, S.; Al-Rajhi, S.; Al-Barwani, T. A review of effect of light on microalgae growth. In Proceedings of the World Congress on Engineering, London, UK, 4–6 July 2012; Volume 2, pp. 1–7.
- Paliwal, C.; Mitra, M.; Bhayani, K.; Bharadwaj, S.V.; Ghosh, T.; Dubey, S.; Mishra, S. Abiotic stresses as tools for metabolites in microalgae. Bioresour. Technol. 2017, 244, 1216–1226.
- Chokshi, K.; Pancha, I.; Trivedi, K.; George, B.; Maurya, R.; Ghosh, A.; Mishra, S. Biofuel potential of the newly isolated microalgae Acutodesmus dimorphus under temperature induced oxidative stress conditions. Bioresour. Technol. 2015, 180, 162–171.
- Converti, A.; Casazza, A.A.; Ortiz, E.Y.; Perego, P.; Del Borghi, M. Effect of temperature and nitrogen concentration on the growth and lipid content of Nannochloropsis oculata and Chlorella vulgaris for biodiesel production. Chem. Eng. Process. 2009, 48, 1146–1151.
- Wang, Y.; He, B.; Sun, Z.; Chen, Y.F. Chemically enhanced lipid production from microalgae under low sub-optimal temperature. Algal Res. 2016, 16, 20–27.
- Zhu, C.J.; Lee, Y.K.; Chao, T.M. Effects of temperature and growth phase on lipid and biochemical composition of Isochrysis galbana TK1. J. Appl. Phycol. 1997, 9, 451–457.
- Jiang, H.; Gao, K. Effects of lowering temperature during culture on the production of polyunsaturated fatty acids in the marine diatom Phaeodactylum tricornutum (bacillariophyceae) 1. J. Phycol. 2004, 40, 651–654.
- Takagi, M.; Yoshida, T. Effect of salt concentration on intracellular accumulation of lipids and triacylglyceride in marine microalgae Dunaliella cells. J. Biosci. Bioeng. 2006, 101, 223–226.
- Pancha, I.; Chokshi, K.; Ghosh, T.; Paliwal, C.; Maurya, R.; Mishra, S. Bicarbonate supplementation enhanced biofuel production potential as well as nutritional stress mitigation in the microalgae Scenedesmus sp. CCNM 1077. Bioresour. Technol. 2015, 193, 315–323.
- Duan, X.; Ren, G.Y.; Liu, L.L.; Zhu, W.X. Salt-induced osmotic stress for lipid overproduction in batch culture of Chlorella vulgaris. Afr. J. Biotechnol. 2012, 11, 7072–7078.
- Cao, J.; Yuan, H.; Li, B.; Yang, J. Significance evaluation of the effects of environmental factors on the lipid accumulation of Chlorella minutissima UTEX 2341 under low-nutrition heterotrophic condition. Bioresour. Technol. 2014, 152, 177–184.
- Sajjadi, B.; Chen, W.Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232.
- Liu, Z.Y.; Wang, G.C.; Zhou, B.C. Effect of iron on growth and lipid accumulation in Chlorella vulgaris. Bioresour. Technol. 2008, 99, 4717–4722.
- Mao, X.; Lao, Y.; Sun, H.; Li, X.; Yu, J.; Chen, F. Time-resolved transcriptome analysis during transitions of sulfur nutritional status provides insight into triacylglycerol (TAG) and astaxanthin accumulation in the green alga Chromochloris zofingiensis. Biotechnol. Biofuels. 2020, 13, 128.
- Wang, Q.; Zhang, Y.; Wu, H.; Xu, N.; Li, A. Effects of sulfur limitation on nitrogen and sulfur uptake and lipid accumulation in Scenedesmus acuminatus. J. Appl. Phycol. 2021, 33, 301–311.
- Xu, L.; Cheng, X.; Wang, Q. Enhanced lipid production in Chlamydomonas reinhardtii by co-culturing with Azotobacter chroococcum. Front. Plant Sci. 2018, 9, 741.
- Dao, G.H.; Wu, G.X.; Wang, X.X.; Zhuang, L.L.; Zhang, T.Y.; Hu, H.Y. Enhanced growth and fatty acid accumulation of microalgae Scenedesmus sp. LX1 by two types of auxin. Bioresour. Technol. 2018, 247, 561–567.
- Singh, J.; Jain, D.; Agarwal, P.; Singh, R.P. Auxin and cytokinin synergism augmenting biomass and lipid production in microalgae Desmodesmus sp. JS07. Process Biochem. 2020, 95, 223–234.
- Bhuyar, P.; Yusoff, M.M.; Rahim, M.H.A.; Sundararaju, S.; Maniam, G.P.; Govindan, N. Effect of plant hormones on the production of biomass and lipid extraction for biodiesel production from microalgae Chlorella sp. J. Microbiol. Biotechnol. Food Sci. 2021, 9, 671–674.
- Che, R.; Huang, L.; Xu, J.W.; Zhao, P.; Li, T.; Ma, H.; Yu, X. Effect of fulvic acid induction on the physiology, metabolism, and lipid biosynthesis-related gene transcription of Monoraphidium sp. FXY-10. Bioresour. Technol. 2017, 227, 324–334.
- Udayan, A.; Sabapathy, H.; Arumugam, M. Stress hormones mediated lipid accumulation and modulation of specific fatty acids in Nannochloropsis oceanica CASA CC201. Bioresour. Technol. 2020, 310, 123437.
- Zabochnicka-Świątek, M.; Kamizela, T.; Kowalczyk, M.; Kalaji, H.M.; Bąba, W. Inexpensive and universal growth media for biomass production of microalgae. Glob. Nest. J. 2019, 21, 82–89.
- Ananthi, V.; Brindhadevi, K.; Pugazhendhi, A.; Arun, A. Impact of abiotic factors on biodiesel production by microalgae. Fuel 2021, 284, 118962.
- Smachetti, M.E.S.; Coronel, C.D.; Salerno, G.L.; Curatti, L. Sucrose-to-ethanol microalgae-based platform using seawater. Algal Res. 2020, 45, 101733.
- Nagarajan, D.; Lee, D.J.; Chang, J.S. Circular bioeconomy: An introduction. In Biomass, Biofuels, Biochemicals; Elsevier: Amsterdam, The Netherlands, 2021; pp. 3–23.
- Behera, B.; Paramasivan, B. Research trends and market opportunities of microalgal biorefinery technologies from circular bioeconomy perspectives. Bioresour. Technol. 2022, 351, 127038.
- Choudhary, S.; Tripathi, S.; Poluri, K.M. Microalgal-based bioenergy: Strategies, prospects, and sustainability. Energy Fuel 2022, 36, 14584–14612.
- Sarangi, P.K.; Singh, A.K.; Srivastava, R.K.; Gupta, V.K. Recent Progress and Future Perspectives for Zero Agriculture Waste Technologies: Pineapple Waste as a Case Study. Sustainability 2023, 15, 3575.
- Kuppan, P.; Sudharsanam, A.; Venkateswarlu, K.; Megharaj, M. Solar technology—Closed loop synergy facilitates low-carbon circular bioeconomy in microalgal wastewater treatment. NPJ Clean Water 2023, 6, 43.
- Kusmayadi, A.; Leong, Y.K.; Yen, H.W.; Huang, C.Y.; Chang, J.S. Microalgae as sustainable food and feed sources for animals and humans–biotechnological and environmental aspects. Chemosphere 2021, 271, 129800.
- Cheirsilp, B.; Maneechote, W.; Srinuanpan, S.; Angelidaki, I. Microalgae as Tools for Bio-Circular-Green Economy: Zero-waste Approaches for Sustainable Production and Biorefineries of Microalgal Biomass. Bioresour. Technol. 2023, 387, 129620.
- Idenyi, J.N.; Eya, J.C.; Nwankwegu, A.S.; Nwoba, E.G. Aquaculture sustainability through alternative dietary ingredients: Microalgal value-added products. Eng. Microbiol. 2022, 2, 100049.
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other nutrients from Haematococcus pluvialis—Multifunctional applications. Mar. Drugs 2020, 18, 459.
- Guccione, A.; Biondi, N.; Sampietro, G.; Rodolfi, L.; Bassi, N.; Tredici, M.R. Chlorella for protein and biofuels: From strain selection to outdoor cultivation in a Green Wall Panel photobioreactor. Biotechnol. Biofuels 2014, 7, 84.
- Patel, A.; Krikigianni, E.; Rova, U.; Christakopoulos, P.; Matsakas, L. Bioprocessing of volatile fatty acids by oleaginous freshwater microalgae and their potential for biofuel and protein production. J. Chem. Eng. 2022, 438, 135529.
- Callejón, M.J.J.; Medina, A.R.; Sánchez, M.D.M.; Moreno, P.A.G.; López, E.N.; Cerdán, L.E.; Grima, E.M. Supercritical fluid extraction and pressurized liquid extraction processes applied to eicosapentaenoic acid-rich polar lipid recovery from the microalga Nannochloropsis sp. Algal Res. 2022, 61, 102586.
- Oliver, L.; Fernández-de-Castro, L.; Dietrich, T.; Villaran, M.C.; Barrio, R.J. Production of docosahexaenoic acid and odd-chain fatty acids by microalgae Schizochytrium limacinum grown on waste-derived volatile fatty acids. Appl. Sci. 2022, 12, 3976.