Bovine mastitis is a polymicrobial disease characterised by inflammation of the udders of dairy and beef cattle. The infection has huge implications to health and welfare of animals, impacting milk and beef production and costing up to EUR 32 billion annually to the dairy industry, globally. Bacterial communities associated with the disease include representative species from Staphylococcus, Streptococcus, Enterococcus, Actinomyces, Aerococcus, Escherichia, Klebsiella and Proteus. Conventional treatment relies on antibiotics, but antimicrobial resistance, declining antibiotic innovations and biofilm production negatively impact therapeutic efficacy. Bacteriophages (phages) are viruses which effectively target and lyse bacteria with extreme specificity and can be a valuable supplement or replacement to antibiotics for bovine mastitis.
- Staphylococcus
- Streptococcus
- Enterococcus
- Actinomyces
- Aerococcus
1. Impact of Bovine Mastitis
Although the main cause of mastitis is bacterial sources, this is not because of any one particular species as there are a number of different species which can cause the condition. The major organisms (represented by >1% of all Isolates) identified in the work of Aarestrup et al. were generally members of the genus Staphylococcus, most commonly S. aureus, S. chromogenes, S. epidermidis, S. haemolyticus, S. simulans, and S. warneri, together with organisms from other genera, namely Streptococcus dysgalactiae, S. uberis, S. canis and Enterococcus faecalis [5]. However, the researchers also succeeded in isolating other members of the genus Staphylococcus at lower frequencies, namely S. auricularis, S. capitis, S. cohnii, S. hominis, S. lentis, S. muscae, S. saprophyticus and S. sciuru, together with non-specified/unidentified members of the same genus. The other sources of infection which they isolated were Actinomyces pyogenes, Aerococcus hydro, A. viridans, E. avium, E. durans, E. faecium, Escherichia coli, Klebsiella pneumoniae, unidentified members of the genus Proteus, S. lactis and S. salivarius. However, this is just an example of a single case study, and other studies have reported cases where some of the organisms found at minor levels were either present at higher relative abundance or as the sole or predominant causal organism, or indeed found other organisms capable of causing mastitis [6].
An example of another organism with the potential to cause mastitis is S. agalactiae. This is an organism which has been isolated from a range of different animals, including non-mammalian species [7], demonstrating that its role goes beyond causing mastitis. The example of S. agalactiae is an interesting one as it is not only able to infect a range of host species, but it can also be the causal organism for more than one clinical condition. For example, as well as being an organism which can lead to bovine mastitis, it has been described as being responsible for both sepsis and meningitis in humans, and also meningoencephalitis in fish [7]. Moreover, even at subclinical levels, S. agalactiae has been shown to have a detrimental impact on milk production, in terms of both quantity and milk quality [8]. Although a fairly large number of species are included in the list above, there are other examples of species which have been associated with the onset of mastitis. These include K. oxytoca [9], Mycobacterium bovis [10], Pseudomonas aeruginosa [11] and S. xylosus [12,13].
The obvious multidrug resistance shown towards a wide range of chemical antimicrobials are compelling and triggers the urgent need to identify and develop alternative strategies to control bovine mastitis safely and effectively [19,20]. A novel and emerging treatment which explores the application of bacteriophages (phages, viruses of bacteria) has been shown to greatly mitigate bacterial resistance and can improve the general health and production capacity of livestock [20,26].
2. The Case for Phage Therapy to Control Bovine Mastitis
2.1. Phage Specificity, Lysis and Amplification
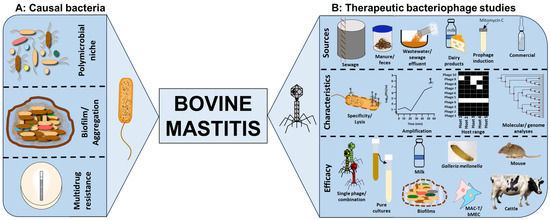
2.2. Isolation of Phages from a Wide Range of Sources
Phages are the most abundant entities on earth with a ~1031 PFU/mL reported concentration [76,77,78][20][21][22]. For bovine mastitis, various sample sources have been explored for the purpose of isolating phages for the various pathogens responsible for the infection. Most of the work has focused on screening raw milk samples obtained from a confirmed mastitis cattle, either directly after centrifuging and filtering of samples or via enrichment procedures to amplify and isolate phages [28,34][23][24]. Phages have been reported to actively bind to, lyse and amplify in milk constituents, and huge successes of phage isolation for S. aureus, S. agalactiae and S. arlettae have been recorded from this source through this method [7,28,29,34,40,41,46,49,63][9][23][24][25][26][27][28][29][30]. However, in one instance, no phage was isolated from the milk samples examined [48][31]. The reason for this may be attributed to the bovine whey protein which may prevent attachment of some phages [58][32]. This may also simply be the lack of phages specific for the bacterial host used as a target for the isolation or the occurrence of the phages in very low titers requiring an enrichment procedure to enable viral amplification and enhance detection [48][31]. Milk products have also been examined and yielded phages for bovine mastitis pathogens Staphylococcus and Streptococcus via enrichment of Cabrales and Peñamellera cheeses, although all phages isolated from this method yielded temperate phages [34][24]. Moreover, S. aureus and S. arlettae phages of lysogenic origin have been isolated from milk as well [34[24][26][27][30][33],40,41,56,63], although for regulatory purposes, lytic phages are preferred to temperate phages due to the possibility of lysogeny occurring and transfer of virulence genes via horizontal gene transfer. However, where strictly lytic phages are not isolated, temperate phages showed potential therapeutic efficacy and are particularly useful for treatment [79,80][34][35]. Sewage, sewage effluent, sewage water, barn flushes, wastewater, cowshed water and manure from dairy farms have yielded a large quantity of phages targeting mastitis-causing pathogens which may be attributed to the microbial richness in these sources [32,37,42,43,48,51,52,53][8][31][36][37][38][39][40][41]. Other very odd sources such as pig manure have been a good source to isolate phages for the infection; this may reveal the interconnection of niches for these organisms [50,54,55][10][42][43]. Phages from commercial sources such as 23361 (ATCC), BP39 (PhageLux) and SAML-4, SAML-12, SAML-150, SAML-4229 and SATA-8505 (StaphLyse) have been investigated for potential usage for bovine mastitis S. aureus [30,31][44][45].2.3. Cocktail Optimisation to Improve Therapeutic Activity
Therapeutic activity of single-phage treatments can significantly reduce bacterial load in many infection models using optimal multiplicity of infection (MOI; the ratio of infecting phages to bacteria in a given infection challenge) as shown in many studies [32,43,48,49,51,52,53,60][18][29][31][36][38][39][40][41]. However, phage resistance was detected within as early as 2 hours after phage treatment as indicated by the regrowth of cultures after lysis which can negatively impact therapeutic efficacy [49][29]. To curtail resistance and lysogeny development, broaden host target coverage and specificity, and to improve lysis efficiency, a cocktail of diverse phages can be optimised [62,81,82,83][46][47][48][49]. This strategy has proven successful, and various combinations of diverse phage morphologies have shown beneficial combinatorial effects in clearing several bacteria causing bovine mastitis. For example, a four-phage cocktail was developed for E. coli and cocktails of two or three phages were shown to be more effective than single-phage treatments for S. aureus [30,34,35,54,55,61,62][24][42][43][44][46][50][51]. Similarly, the therapeutic efficacy of a phage cocktail was shown to be comparable to that of the antibiotic ceftiofur sodium for E. coli in cattle and S. aureus in mice [35,61][50][51]. This has been extrapolated further, with a cocktail of four phages together with the lactic acid bacterium Lactiplantibacillus plantarum proving effective [54][42]. Phage activity on S. aureus was shown to be delayed by IgG-dependent aggregation using single-phage treatment. While in contrast, the use of a cocktail showed no significant effect with or without IgG in milk [30][44].2.4. Characterisation of Phage Lysis and Stability in Pure Cultures
Several therapeutic assessments have investigated the efficacy and safety of phages for the targeted eradication of bovine mastitis. Fundamental research has been conducted regarding phage activity in pure cultures to determine lysis capabilities by individual phages and in combination with other phages. Host range analysis mainly focuses on phage lysis activity using spot test with the double-layer agar method (application of phage samples to confluent cultures of bacteria in a semi-solid agar medium overlayed on solid agar medium). This is to ascertain the range of relevant bacterial strains the phages can lyse with some demonstrating broad or narrower host coverage [28,36,37][23][37][52]. Besides phage coverage on a wide range of strains, other phages of S. aureus showed inter-species lysis, targeting S. sciuri and Rothia terrae [28][23] as well as E. coli [43][38], and K. oxytoca phage P2 lysing E. aerogenes as well [9][19]. Further work was also directed to stability (in various temperature and pH ranges) and killing assays in pure cultures in broth or liquid media and milk (pasteurised and unpasteurised) using MOI assays in a given infection model to provide an insight into the dosage [54,55][42][43]. Data showed a wide range of effectiveness of MOI range of 0.001 to 100 in vitro [43,48,49,51,52,53,63,68][17][29][30][31][38][39][40][41]. However, optimal effectiveness was at MOI of 10 in vivo for some of the data [60][18]. Other reports showed that efficacy was achieved in a phage-dose-dependent manner in milk using an S. aureus phage [31][45].2.5. Phage Therapeutic Activity in Biofilms
The pathogens causing mastitis can aggregate in vitro and in vivo in extracellular polysaccharide-containing biofilm matrixes which restricts antibiotic access to bacteria [17,18,47,84][53][54][55][56]. Phages have been shown to prevent or penetrate established biofilms produced by mastitis bacteria in vitro and in vivo, hence showing the potential to be used as a standalone treatment or to supplement antibiotic use and enhance therapeutic efficacy [47,49][29][55]. The phages can lyse bacteria early in the culture to prevent biofilm formation or may disrupt established biofilms which can enhance bacterial killing or provide pathogens access within the biofilm matrix [72,82][5][48]. In S. aureus biofilms, treatment using a single phage or a cocktail of phages significantly reduced bacterial load in planktonic cultures as well as established biofilms on polystyrene surfaces, in milk and on mammary glands [47,49][29][55]. As well as the issue of potential protection from biofilms, mastitis-causing bacteria have been shown to be afforded some level of protection from bacterial aggregation [56][33], including during the times when S. aureus was exposed to phage infection. However, previous work showed a total kill of S. aureus, which has a few cells and have survived phage treatment, probably by some level of aggregation. This means that the numbers remaining are sufficiently low for them in turn to be removed by the animal’s own immune system [30][44].2.6. Phage Therapeutic Assessments in Mastitis Ex Vivo and In Vivo Models
Phages have low inherent toxicity to the immune system, and they are potentially cheaper to isolate and develop, which provides an economic advantage over antibiotics [72][5]. To contextualise and provide insight into the therapeutic safety and efficacy of phages, relevant ex vivo models involving bovine cells lines were investigated. The studied cell lines for bacterial and phage interactions for this are the mammary alveolar cells-large T antigen (MAC-T) and bovine mammary epithelial (bMEC) cell lines [51,52,65][39][40][57]. A cocktail of two phages, CM8-1 and SJT-2 was shown to reduce K. pneumonia numbers and consequently reduce adhesion, invasion, and cytotoxicity in bMEC cells [65][57]. S. aureus phages were shown to migrate intercellularly and could reach the nucleus within 3 h after exposure to MAC-T cell lines and have an endocytotic activity of 12% in a bovine ex vivo model [51,52][39][40]. Studies on S. aureus-colonised G. mellonella larvae showed a 50% survival rate four days after treatment with a single phage [44][58]. The in vivo model that has been extensively studied for bovine mastitis phage therapy is the mouse model mainly because this model has itself been well established for infection since the 1970s [35,44,50,60,64,68][10][17][18][50][58][59]. Results in mice showed favourable outcomes for phage therapy with reduced colonisation and reduced inflammatory cytokines as soon as 24 hours after treatment. The mouse model has also been reported to be a more time- and cost-effective model than those of larger mammals with comparable symptoms, inflammatory indicators, colonisation, and histopathological characteristics. Therapeutic efficacies have been achieved in cattle as well [57,61][51][60].3. Barriers/Challenges to Therapeutic Phage Application to Control Bovine Mastitis
Researchers outlined the advantages of phage therapy and research work conducted in the area to control bovine mastitis. However, a degree of caution needs to be applied by anyone considering using it as a potential prophylactic treatment. It has been reported that the infusion of a phage sample into unaffected quarters in the udder of lactating dairy cattle resulted in an increase in the somatic cell count in the milk from that quarter [57][60]. This suggests that there has been some form of immune response taking place in that particular quarter of the cow’s udder. A comparable increase in somatic cell count was not seen in animals infused with a phage sample where the animal had some level of mastitis infection, even at a sub-clinical level [57][60]. The situation in terms of using phage as a treatment for mastitis is complicated, yet evidence exists to show that in S. aureus the whey proteins in milk can adhere to the surface of cells, thereby blocking potential attachment sites for the phage [58][32]. Moreover, it was shown that in raw milk, as opposed to milk which has been heat-treated, phage K which has the potential to infect and kill S. aureus was less successful [59][61]. It is thought that this is due to the clumping of the bacteria on fat globules within the milk and some sort of presumed protection from this activity. On the other hand, the lysogenisation of the bacterial host by temperate phages could potentially cause the exchange of virulence factors via horizontal gene transfer as stated above. However, the use of phages can come with additional complications. One such example of this was seen where a phage which entered the lysogenic phase was also found to contain a gene which conferred resistance to multiple types of antibiotics [12][62]. Therefore, although there is a clear potential for usage of phages as a means of killing bacteria causing mastitis infections, there needs to be considerable research undertaken before using these phages as treatments. Temperate phages can access the lytic life cycle via induction through treatment with mitomycin C as shown in S. galactiae [56][33] or activation of the repressor or deletion of the integrase genes. Unfortunately, they are unsuitable for therapeutic purposes in their wild form. However, genetic engineering has provided avenues for genetic manipulation to help develop therapeutically acceptable phages where strictly lytic ones are not available [85,86,87][63][64][65]. The polymicrobial niche of bovine mastitis is also a challenge to overcome [5,6,16][66][67][68]. Most work conducted to date has focused on a single bacterial species in relevant infection model systems, except for example where S. aureus phages showed interspecies lysis on S. sciuri and Rothia terrae [28][23], and E. coli [43][38]. Whilst this is informative and provides useful insights into the therapeutic potential of the phages, it is still unclear how these single bacterial species targets would alleviate bovine mastitis. More work is therefore needed on multispecies targets through phage cocktail optimisations to clear the bacterial communities as standalone treatments or as adjunct to antibiotics for the effective clearance of bovine mastitis infection.References
- Bond, D.M.; Morris, J.M.; Nassar, N. Study protocol: Evaluation of the probiotic Lactobacillus Fermentum CECT5716 for the prevention of mastitis in breastfeeding women: A randomised controlled trial. BMC Pregnancy Childbirth 2017, 17, 148.
- Radostits, O.M.; Gay, C.C.; Hinchcliff, K.W.; Constable, P.D. Veterinary Medicine: A Textbook of the Diseases of Cattle, Horses, Sheep, Pigs and Goats, 10th ed.; Elsevier: St. Louis, MO, USA, 2007.
- Persson Waller, K.; Persson, Y.; Nyman, A.K.; Stengärde, L. Udder health in beef cows and its association with calf growth. Acta Vet. Scand. 2014, 56, 9.
- Rollin, E.; Dhuyvetter, K.C.; Overton, M.W. The cost of clinical mastitis in the first 30 days of lactation: An economic modeling tool. Prev. Vet. Med. 2015, 122, 257–264.
- Loc-Carrillo, C.; Abedon, S.T. Pros and cons of phage therapy. Bacteriophage 2011, 1, 111–114.
- Taslem Mourosi, J.; Awe, A.; Guo, W.; Batra, H.; Ganesh, H.; Wu, X.; Zhu, J. Understanding Bacteriophage Tail Fiber Interaction with Host Surface Receptor: The Key “Blueprint” for Reprogramming Phage Host Range. Int. J. Mol. Sci. 2022, 23, 12146.
- Kizziah, J.L.; Manning, K.A.; Dearborn, A.D.; Dokland, T. Structure of the host cell recognition and penetration machinery of a Staphylococcus aureus bacteriophage. PLoS Pathog. 2020, 16, e1008314.
- Leite, J.A.; Pereira, H.P.; Borges, C.A.V.; Alves, B.R.C.; Ramos, A.; Martins, M.F.; Arcuri, E.F. Lytic bacteriophages as a potential alternative to control Staphylococcus aureus. Pesqui. Agropecu. Bras. 2019, 54.
- Bai, Q.; Zhang, W.; Yang, Y.; Tang, F.; Nguyen, X.; Liu, G.; Lu, C. Characterization and genome sequencing of a novel bacteriophage infecting Streptococcus agalactiae with high similarity to a phage from Streptococcus pyogenes. Arch. Virol. 2013, 158, 1733–1741.
- Wang, Z.F.; Zheng, P.P.; Ji, W.H.; Fu, Q.; Wang, H.G.; Yan, Y.X.; Sun, J.H. SLPW: A Virulent Bacteriophage Targeting Methicillin-Resistant Staphylococcus aureus In vitro and In vivo. Front. Microbiol. 2016, 7, 934.
- Wang, Z.; Xue, Y.; Gao, Y.; Guo, M.; Liu, Y.; Zou, X.; Cheng, Y.; Ma, J.; Wang, H.; Sun, J.; et al. Phage vB_PaeS-PAJD-1 Rescues Murine Mastitis Infected With Multidrug-Resistant Pseudomonas aeruginosa. Front. Cell Infect. Microbiol. 2021, 11, 689770.
- Zhang, Q.; Yu, H.; Sun, Y.; Zhangxiang, L.; Zhang, P.; Liu, G.; Qu, Y.; Tong, Y.; Li, Y. Isolation and characterization of a lytic phage infecting Enterococcus faecium of bovine mastitis. Acta Vet. Zootech. Sin. 2017, 48, 706–713.
- Wang, I.N.; Smith, D.L.; Young, R. Holins: The protein clocks of bacteriophage infections. Annu. Rev. Microbiol. 2000, 54, 799–825.
- Han, J.E.; Kim, J.H.; Hwang, S.Y.; Choresca, C.H., Jr.; Shin, S.P.; Jun, J.W.; Chai, J.Y.; Park, Y.H.; Park, S.C. Isolation and characterization of a Myoviridae bacteriophage against Staphylococcus aureus isolated from dairy cows with mastitis. Res. Vet. Sci. 2013, 95, 758–763.
- Makumi, A.; Mhone, A.L.; Odaba, J.; Guantai, L.; Svitek, N. Phages for Africa: The Potential Benefit and Challenges of Phage Therapy for the Livestock Sector in Sub-Saharan Africa. Antibiotics 2021, 10, 1085.
- Ferriol-González, C.; Domingo-Calap, P. Phage Therapy in Livestock and Companion Animals. Antibiotics 2021, 10, 559.
- Xi, H.Y.; He, D.L.; Li, D.; Liu, S.S.; Wang, G.; Ji, Y.L.; Wang, X.W.; Wang, Z.J.; Bi, L.T.; Zhao, R.H.; et al. Bacteriophage Protects Against Aerococcus viridans Infection in a Murine Mastitis Model. Front. Vet. Sci. 2020, 7, 588.
- Duarte, V.D.; Dias, R.S.; Kropinski, A.M.; Campanaro, S.; Treu, L.; Siqueira, C.; Vieira, M.S.; Paes, I.D.; Santana, G.R.; Martins, F.; et al. Genomic analysis and immune response in a murine mastitis model of vB_EcoM-UFV13, a potential biocontrol agent for use in dairy cows. Sci. Rep. 2018, 8, 6845.
- Fahliyani, S.A.; Beheshti-Maal, K.; Ghandehari, F. Novel lytic bacteriophages of Klebsiella oxytoca ABG-IAUF-1 as the potential agents for mastitis phage therapy. Fems Microbiol. Lett. 2018, 365, fny223.
- Hendrix, R.W.; Smith, M.C.M.; Burns, R.N.; Ford, M.E.; Hatfull, G.F. Evolutionary relationships among diverse bacteriophages and prophages: All the world’s a phage. Proc. Natl. Acad. Sci. USA 1999, 96, 2192–2197.
- Mushegian, A.R. Are There 1031 Virus Particles on Earth, or More, or Fewer? J. Bacteriol. 2020, 202, 10–1128.
- Clokie, M.R.; Millard, A.D.; Letarov, A.V.; Heaphy, S. Phages in nature. Bacteriophage 2011, 1, 31–45.
- Barasuol, B.M.; Cargnelutti, J.F.; Sangioni, L.A.; Pereira, D.I.B.; Varela, A.P.M.; Mayer, F.Q.; Pottker, E.S.; Goncalves, G.F.; Cibulski, S.; Botton, S.D. Characterization of novel of temperate phages of Staphylococcus aureus isolated from bovine milk. Arch. Microbiol. 2022, 204, 680.
- García, P.; Madera, C.; Martínez, B.; Rodríguez, A.; Evaristo Suárez, J. Prevalence of bacteriophages infecting Staphylococcus aureus in dairy samples and their potential as biocontrol agents. J. Dairy Sci. 2009, 92, 3019–3026.
- Basdew, I.H.; Laing, M.D. Investigation of the lytic ability of South African bacteriophages specific for Staphylococcus aureus, associated with bovine mastitis. Biocontrol Sci. Technol. 2015, 25, 429–443.
- Jia, H.; Bai, Q.; Yang, Y.; Yao, H. Complete Genome Sequence of Staphylococcus aureus Siphovirus Phage JS01. Genome Announc. 2013, 1, e00797-13.
- Jia, H.; Dong, W.; Yuan, L.; Ma, J.; Bai, Q.; Pan, Z.; Lu, C.; Yao, H. Characterization and complete genome sequence analysis of Staphylococcus aureus bacteriophage JS01. Virus Genes 2015, 50, 345–348.
- Slanetz, L.W.; Jawetz, E. Isolation and Characteristics of Bacteriophages for Staphylococci of Bovine Mastitis. J. Bacteriol. 1941, 41, 447–455.
- Teng, F.; Xiong, X.; Zhang, S.; Li, G.; Wang, R.; Zhang, L.; Wang, X.; Zhou, H.; Li, J.; Li, Y.; et al. Efficacy Assessment of Phage Therapy in Treating Staphylococcus aureus-Induced Mastitis in Mice. Viruses 2022, 14, 620.
- Han, G.; Zhang, J.; Luo, Z.; Lu, B.; Zhang, P.; Yong, K.; Wang, Y.; Luo, Y.; Yang, Z.; Ren, M.; et al. Characteristics of a novel temperate bacteriophage against Staphylococcus arlettae (vB_SarS_BM31). Int. Microbiol. 2023, 26, 327–341.
- Srujana, A.S.; Sonalika, J.; Akhila, D.S.; Juliet, M.R.; Sheela, P. Isolation of Phages and Study of their In vitro Efficacy on Staphylococcus aureus Isolates Originating from Bovine Subclinical Mastitis. Indian. J. Anim. Res. 2022, 56, 754–758.
- Gill, J.J.; Sabour, P.M.; Leslie, K.E.; Griffiths, M.W. Bovine whey proteins inhibit the interaction of Staphylococcus aureus and bacteriophage K. J. Appl. Microbiol. 2006, 101, 377–386.
- Bai, Q.; Yang, Y.; Lu, C. Isolation and characterization of siphovirus phages infecting bovine Streptococcus agalactiae. Wei Sheng Wu Xue Bao 2016, 56, 317–326.
- Dedrick, R.M.; Guerrero-Bustamante, C.A.; Garlena, R.A.; Russell, D.A.; Ford, K.; Harris, K.; Gilmour, K.C.; Soothill, J.; Jacobs-Sera, D.; Schooley, R.T.; et al. Engineered bacteriophages for treatment of a patient with a disseminated drug-resistant Mycobacterium abscessus. Nat. Med. 2019, 25, 730–733.
- Nale, J.Y.; Clokie, M.R.J. Preclinical data and safety assessment of phage therapy in humans. Curr. Opin. Biotechnol. 2021, 68, 310–317.
- Dias, R.S.; Eller, M.R.; Duarte, V.S.; Pereira, Â.L.; Silva, C.C.; Mantovani, H.C.; Oliveira, L.L.; Silva Ede, A.; De Paula, S.O. Use of phages against antibiotic-resistant Staphylococcus aureus isolated from bovine mastitis. J. Anim. Sci. 2013, 91, 3930–3939.
- Iwano, H.; Inoue, Y.; Takasago, T.; Kobayashi, H.; Furusawa, T.; Taniguchi, K.; Fujiki, J.; Yokota, H.; Usui, M.; Tanji, Y.; et al. Bacteriophage Phi SA012 Has a Broad Host Range against Staphylococcus aureus and Effective Lytic Capacity in a Mouse Mastitis Model. Biology 2018, 7, 8.
- Li, L.P.; Zhang, Z.Y. Isolation and characterization of a virulent bacteriophage SPW specific for Staphylococcus aureus isolated from bovine mastitis of lactating dairy cattle. Mol. Biol. Rep. 2014, 41, 5829–5838.
- Zhang, L.; Bao, H.; Wei, C.; Zhang, H.; Zhou, Y.; Wang, R. Characterization and partial genomic analysis of a lytic Myoviridae bacteriophage against Staphylococcus aureus isolated from dairy cows with mastitis in Mid-east of China. Virus Genes 2015, 50, 111–117.
- Zhang, L.L.; Sun, L.C.; Wei, R.C.; Gao, Q.; He, T.; Xu, C.F.; Liu, X.J.; Wang, R. Intracellular Staphylococcus aureus Control by Virulent Bacteriophages within MAC-T Bovine Mammary Epithelial Cells. Antimicrob. Agents Chemother. 2017, 61, 10–1128.
- Zhang, Q.; Xing, S.Z.; Sun, Q.; Pei, G.Q.; Cheng, S.; Liu, Y.N.; An, X.P.; Zhang, X.L.L.; Qu, Y.G.; Tong, Y.G. Characterization and complete genome sequence analysis of a novel virulent Siphoviridae phage against Staphylococcus aureus isolated from bovine mastitis in Xinjiang, China. Virus Genes 2017, 53, 464–476.
- Titze, I.; Krömker, V. Antimicrobial Activity of a Phage Mixture and a Lactic Acid Bacterium against Staphylococcus aureus from Bovine Mastitis. Vet. Sci. 2020, 7, 31.
- Titze, I.; Lehnherr, T.; Lehnherr, H.; Krömker, V. Efficacy of Bacteriophages Against Staphylococcus aureus Isolates from Bovine Mastitis. Pharmaceuticals 2020, 13, 35.
- Breyne, K.; Honaker, R.W.; Hobbs, Z.; Richter, M.; Żaczek, M.; Spangler, T.; Steenbrugge, J.; Lu, R.; Kinkhabwala, A.; Marchon, B. Efficacy and safety of a bovine-associated Staphylococcus aureus phage cocktail in a murine model of mastitis. Front. Microbiol. 2017, 8, 2348.
- Brouillette, E.; Millette, G.; Chamberland, S.; Roy, J.P.; Ster, C.; Kiros, T.; Hickey, S.; Hittle, L.; Woolston, J.; Malouin, F. Effective Treatment of Staphylococcus aureus Intramammary Infection in a Murine Model Using the Bacteriophage Cocktail StaphLyse™. Viruses 2023, 15, 887.
- Porter, J.; Anderson, J.; Carter, L.; Donjacour, E.; Paros, M. In vitro evaluation of a novel bacteriophage cocktail as a preventative for bovine coliform mastitis. J. Dairy Sci. 2016, 99, 2053–2062.
- Principi, N.; Silvestri, E.; Esposito, S. Advantages and Limitations of Bacteriophages for the Treatment of Bacterial Infections. Front. Pharmacol. 2019, 10, 513.
- Nale, J.Y.; Chutia, M.; Carr, P.; Hickenbotham, P.T.; Clokie, M.R.J. ‘Get in Early’; Biofilm and Wax Moth (Galleria mellonella) Models Reveal New Insights into the Therapeutic Potential of Clostridium difficile Bacteriophages. Front. Microbiol. 2016, 7, 1383.
- Malik, D.J.; Sokolov, I.J.; Vinner, G.K.; Mancuso, F.; Cinquerrui, S.; Vladisavljevic, G.T.; Clokie, M.R.J.; Garton, N.J.; Stapley, A.G.F.; Kirpichnikova, A. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv. Colloid. Interface Sci. 2017, 249, 100–133.
- Geng, H.; Zou, W.; Zhang, M.; Xu, L.; Liu, F.; Li, X.; Wang, L.; Xu, Y. Evaluation of phage therapy in the treatment of Staphylococcus aureus-induced mastitis in mice. Folia Microbiol 2020, 65, 339–351.
- Guo, M.; Gao, Y.; Xue, Y.; Liu, Y.; Zeng, X.; Cheng, Y.; Ma, J.; Wang, H.; Sun, J.; Wang, Z.; et al. Bacteriophage Cocktails Protect Dairy Cows Against Mastitis Caused By Drug Resistant Escherichia coli Infection. Front. Cell Infect. Microbiol. 2021, 11, 690377.
- Hamza, A.; Perveen, S.; Abbas, Z.; Rehman, S.U. The Lytic SA Phage Demonstrate Bactericidal Activity against Mastitis Causing Staphylococcus aureus. Open Life Sci. 2016, 11, 39–45.
- Bissong, M.E.A.; Ateba, C.N. Genotypic and Phenotypic Evaluation of Biofilm Production and Antimicrobial Resistance in Staphylococcus aureus Isolated from Milk, North West Province, South Africa. Antibiotics 2020, 9, 156.
- Fox, L.K.; Zadoks, R.N.; Gaskins, C.T. Biofilm production by Staphylococcus aureus associated with intramammary infection. Vet. Microbiol. 2005, 107, 295–299.
- Song, J.; Ruan, H.; Chen, L.; Jin, Y.; Zheng, J.; Wu, R.; Sun, D. Potential of bacteriophages as disinfectants to control of Staphylococcus aureus biofilms. BMC Microbiol. 2021, 21, 57.
- Pedersen, R.R.; Krömker, V.; Bjarnsholt, T.; Dahl-Pedersen, K.; Buhl, R.; Jørgensen, E. Biofilm Research in Bovine Mastitis. Front. Vet. Sci. 2021, 8, 656810.
- Shi, Y.; Zhao, W.; Liu, G.; Ali, T.; Chen, P.; Liu, Y.; Kastelic, J.P.; Han, B.; Gao, J. Bacteriophages isolated from dairy farm mitigated Klebsiella pneumoniae-induced inflammation in bovine mammary epithelial cells cultured in vitro. BMC Vet. Res. 2021, 17, 37.
- Ngassam-Tchamba, C.; Duprez, J.-N.; Fergestad, M.; De Visscher, A.; L’Abee-Lund, T.; De Vliegher, S.; Wasteson, Y.; Touzain, F.; Blanchard, Y.; Lavigne, R. In vitro and in vivo assessment of phage therapy against Staphylococcus aureus causing bovine mastitis. J. Glob. Antimicrob. Resist. 2020, 22, 762–770.
- Liang, B.; Zhao, W.; Han, B.; Barkema, H.W.; Niu, Y.D.; Liu, Y.; Kastelic, J.P.; Gao, J. Biological and genomic characteristics of two bacteriophages isolated from sewage, using one multidrug-resistant and one non-multidrug-resistant strain of Klebsiella pneumoniae. Front. Microbiol. 2022, 13, 943279.
- Gill, J.J.; Pacan, J.C.; Carson, M.E.; Leslie, K.E.; Griffiths, M.W.; Sabour, P.M. Efficacy and pharmacokinetics of bacteriophage therapy in treatment of subclinical Staphylococcus aureus mastitis in lactating dairy cattle. Antimicrob. Agents Chemother. 2006, 50, 2912–2918.
- O’Flaherty, S.; Coffey, A.; Meaney, W.J.; Fitzgerald, G.F.; Ross, R.P. Inhibition of bacteriophage K proliferation on Staphylococcus aureus in raw bovine milk. Lett. Appl. Microbiol. 2005, 41, 274–279.
- Wipf, J.R.; Schwendener, S.; Perreten, V. The novel macrolide-Lincosamide-Streptogramin B resistance gene erm(44) is associated with a prophage in Staphylococcus xylosus. Antimicrob. Agents Chemother. 2014, 58, 6133–6138.
- Chen, Y.; Batra, H.; Dong, J.; Chen, C.; Rao, V.B.; Tao, P. Genetic Engineering of Bacteriophages Against Infectious Diseases. Front. Microbiol. 2019, 10, 954.
- Gibb, B.; Hyman, P.; Schneider, C.L. The Many Applications of Engineered Bacteriophages—An Overview. Pharmaceuticals 2021, 14, 634.
- Guo, D.; Chen, J.; Zhao, X.; Luo, Y.; Jin, M.; Fan, F.; Park, C.; Yang, X.; Sun, C.; Yan, J.; et al. Genetic and Chemical Engineering of Phages for Controlling Multidrug-Resistant Bacteria. Antibiotics 2021, 10, 202.
- Aarestrup, F.Μ.; Wegener, H.C.; Rosdahl, V.T.; Jensen, Ν.E. Staphylococcal and other Bacterial Species Associated with Intramammary Infections in Danish Dairy Herds. Acta Vet. Scand. 1995, 36, 475–487.
- Aarestrup, F.M.; Wegener, H.C.; Rosdahl, V.T. A comparative study of Staphylococcus aureus strains isolated from bovine subclinical mastitis during 1952–1956 and 1992. Acta Vet. Scand. 1995, 36, 237–243.
- Aarestrup, F.M.; Wegener, H.C.; Rosdahl, V.T. Evaluation of phenotypic and genotypic methods for epidemiological typing of Staphylococcus aureus isolates from bovine mastitis in Denmark. Vet. Microbiol. 1995, 45, 139–150.