Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Zaneta Kimber-Trojnar and Version 2 by Jessie Wu.
Type 2 diabetes mellitus (T2DM) represents a common metabolic disorder that is characterized by chronic hyperglycemia. For more than half a century, the link between insulin resistance and T2DM has been well recognized. Insulin resistance is not only the most powerful predictor of future development of T2DM, but it is also a therapeutic target. On the other hand, gestational diabetes mellitus (GDM) is one of the most common metabolic disorders of pregnancy and its incidence has considerably increased by 10–100% in the last 20 years. There is ample scientific evidence to suggest a link between the fatty acid-binding protein 4 (FABP4) and insulin resistance, gestational (GDM), and type 2 (T2DM) diabetes mellitus.
- fatty acid-binding protein 4
- insulin resistance
- type 2 diabetes mellitus
1. Introduction
Fatty acid-binding protein 4 (FABP4) plays a crucial role in the regulation of lipid-mediated actions, such as the initialization of inflammation and oxidative stress processes [1][33]. The expression of this adipogenic protein can be induced by vascular endothelial growth factor (VEGF) signaling [2][47]. FABP4 increases the ciatriphosphate-binding cassette A1 pathway [3][32]. Emerging data suggests an essential involvement of FABP4 in endothelial dysfunction [2][47]. Exogenous FABP4 interferes with insulin stimulated production of nitric oxide in endothelial cells [4][48]. Cabré et al. [5][44] observed that increased FABP4 concentrations were associated with excessive oxidative stress and inflammatory markers in diabetes.
FABP4 negatively regulates PPARγ in macrophages and adipocytes, affecting adipocyte differentiation. Higher levels of FABP4 and lower levels of PPARγ in visceral adipose tissue, when compared with subcutaneous adipose tissue, suggest a causative link between FABP4 and the metabolic syndrome. It may also explain certain morphological and functional differences between the adipocytes found in these two types of fat tissue. Visceral preadipocytes are known to proliferate and differentiate into mature adipocytes less actively than those from subcutaneous fat. Visceral adipose tissue secretes more proinflammatory cytokines [6][35].
It has also been postulated that the transcription factor forkhead box protein O1 (FOXO1) is involved in lipid metabolism. The available scientific evidence suggests that metformin may have a protective effect against lipid accumulation in macrophages as it decreases FABP4 expression at the mRNA level. The exact mechanism is not yet fully understood; it may deal with decreases in transcription or by promotion of mRNA degradation. For this reason, some authors suggest that metformin should also be perceived as a therapeutic agent for the prevention and targeting of atherosclerosis in metabolic syndrome [7][49].
FABP4 contributes to the accumulation of short-chain free fatty acids and suppresses the activity of relevant proteins in the phosphatidylinositol 3′-kinase (PI3K)-AKT signal pathway. Accordingly, FABP4 inhibits the presence of glucose oxidation and glycolysis and decreases the uptake and utilization of glucose in human organs, such as in muscles and the liver [8][50]. It is known that FABP4 can bind various intracellular fatty acids and probably mediates intracellular lipid trafficking between cellular compartments [9][51]. It might also modulate the availability and composition of fatty acids in muscles and adipose tissues [10][52]. In the myocytes and adipose tissue in mice, improved glucose homeostasis after the ablation of FABP4 was documented [11][53].
Increased FABP4 production could contribute to macrophage activity, possibly by activation of inflammatory pathways, resulting in inflammation [12][42]. An alternative hypothesis, more in line with the published evidence, is that the elevated circulating FABP4 reflects the increased cellular production in both adipocytes and macrophages in response to greater lipid availability, with increased Kupffer cell production of FABP4 triggering an increased inflammatory response [12][13][42,54].
FABP4 plays a crucial role in mediating the endoplasmic reticulum stress observed in macrophages upon lipotoxic signal exposure, which contributes to atherosclerosis, inflammation, and perhaps plaque vulnerability [14][55]. High levels of FABP4 may contribute to adverse prognosis via the induction of endoplasmic reticulum stress in macrophages and upregulation of pro-inflammatory cytokine production. Indeed, FABP4 modulates inflammatory responses in macrophages through a positive feedback loop involving c-Jun NH2-terminal kinases and activator protein-1 [12][15][42,56]. FABP4 is also implicated in modulating the eicosanoid balance by affecting both cyclooxygenase 2 (COX2) activity and leukotriene A4 (LTA4) stability, and upregulates uncoupling protein 2 (UCP2); all these processes influence macrophage function and adipose tissue inflammation. FABP4 can also interact with HSL and Janus kinase 2 (JAK2) [16][57] (Figure 1). Two different mechanisms of the latter action have been proposed. The first one includes direct protein–protein interactions between FABP4 and HSL and JAK2 [17][18][58,59]. FABP4 and HSL may also interact indirectly via two protein kinases A and G pathways [19][29].
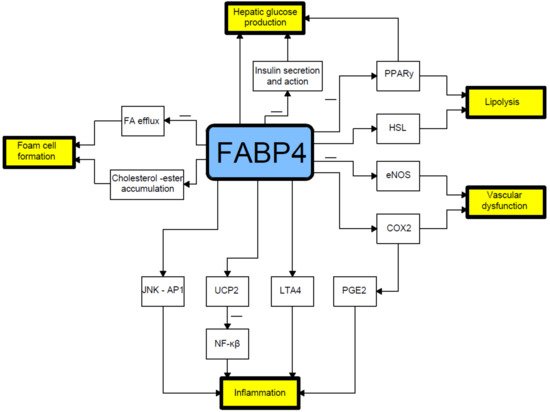
Figure 1. Relationship between fatty acid-binding protein 4 and pathophysiology of type 2 diabetes mellitus. COX2- cyclooxygenase-2; eNOS- endothelial nitric oxide synthase; FA- fatty acid; HSL- hormone sensitive lipase; JNK AP1-Jun N-terminal kinase-activator protein 1; LTA4-leukotriene A4; NF-κB-nuclear factor-kappa B; PGE2-prostaglandin E2; PPARγ–peroxisome proliferator-activated receptor γ; UCP2-uncoupling protein 2.
Several proinflammatory stimuli have been reported to upregulate FABP4 expression in macrophages, including oxidized low-density lipoproteins, toll-like receptor (TLR) agonists, and PPARγ agonists [20][21][22][23][60,61,62,63]. In this respect, lipopolysaccharide (LPS), a TLR4 ligand, stimulates FABP4 expression and the activated FABP4, reciprocally, enhances the LPS-TLR4 signaling-evoked JNK inflammatory pathways [15][56]. In addition, prolonged hyperglycemia has been shown to induce FABP4 expression in mesangial cells and trigger the release of proinflammatory cytokines [20][24][60,64] (Figure 2).
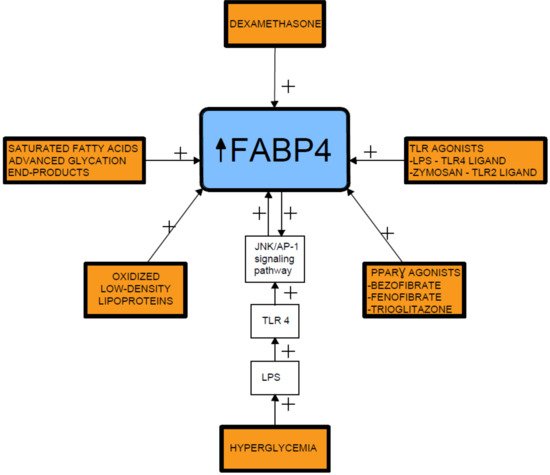
Figure 2.
Fatty acid-binding protein 4 inducing factors. TLR- toll-like receptor; LPS- lipopolysaccharide; FABP4-fatty acid-binding protein 4.
2. Relationship between Fatty Acid-Binding Protein 4 (FABP4) and Insulin Resistance
Fatty acid-binding protein 4 (FABP4) is a cytoplasmic fatty acid chaperone clearly engaged in the onset of insulin resistance [6][35]. Studies in animal models suggest that FABP4 is important for glucose homeostasis [8][50]. Deletion of the FABP4 gene protected mice against insulin resistance as well as hyperinsulinemia associated with both diet-induced obesity and genetic obesity [8][11][13][25][26][27][45,50,53,54,65,66]. A reduced ability of adipocytes to take up and retain free fatty acids, leading to ectopic lipid accumulation, and abnormalities in the release of adipokines by adipocytes are critical factors for insulin resistance and the development of T2DM [27][66].
FABP4 was also detected in apoptotic granulosa cells in atretic antral follicles of the mouse ovary, which suggests a potential link to polycystic ovary syndrome (PCOS), which is known to be frequently associated with insulin resistance [28][11]. Expression of FABP4 mRNA in isolated granulosa cells was found to be higher in patients diagnosed with PCOS than in controls [29][67].
It has been proven that FABP4 is negatively correlated with the glucose-disposal rate (GDR) [30][68]. In non-DM subjects, serum FABP concentrations were negatively correlated with the mean rate of glucose infusion during the last 30 min of the clamp test, which reflects insulin sensitivity [31][69]. Nakamura et al. [6][35] also showed that circulating FABP4 concentrations were negatively correlated with GDR, which is a marker of insulin resistance in skeletal muscles in individuals with T2DM. On the contrary, FABP4 concentration was positively related to the insulinogenic index in non-diabetic participants. Wu et al. [32][70] reported that circulating FABP4 concentrations were correlated with glucose-stimulated insulin secretion in healthy controls.
It has been suggested that insulinotropic potential of FABP4 is similar to the effects of glucagon-like peptide-1 (GLP-1) [6][35]. To maintain glucose homeostasis, FABP4 may stimulate β cells and alter insulin secretion. Moreover, FABP4 presented a positive relationship with insulin secretion at an early stage in the non-diabetic group, which may be due to the fact that insulin secretion is damaged relatively early in T2DM.
Nakamura et al. [6][35] found the most pronounced negative correlation between FABP4 and GDR when compared to some other markers of insulin resistance or body composition in T2DM. FABP4 has been reported to have a negative correlation with GDR in type 1 diabetes mellitus, T2DM, and the controls of Asian Americans [33][71]. FABP4 represents an important molecule dealing with insulin resistance in T2DM.