Epizootic Hemorrhagic Disease (EHD) of ruminants is a viral pathology that has significant welfare, social, and economic implications. The causative agent, epizootic hemorrhagic disease virus (EHDV), belongs to the Orbivirus genus and leads to significant regional disease outbreaks among livestock and wildlife in North America, Asia, Africa, and Oceania, causing significant morbidity and mortality. Here, we comprehensively review the current knowledge on EHDV.
- EHDV
- epidemiology
- vaccines
- white-tailed deer
- cattle
1. Introduction
2. EHDV, the Etiological Agent of EHD
Similar to members of the Orbivirus genus, EHDV non-enveloped virions present a structure characterized by an icosahedral capsid, which is divided into three consecutive protein layers: the inner and intermediate layers (core) and an outer capsid [1] (Figure 1). The genome is located inside the core particle and comprises ten linear double-strand RNA segments that encode for seven structural (VP1–VP7) and at least four non-structural proteins (NS1, NS2, NS3/NS3A, NS4, and probably the putative NS5 as for BTV [2]) [1,3,4][1][3][4]. As for the prototype BTV, the outer protein layer is made of 60 trimers of VP2, the most exposed virion protein, and 120 trimers of VP5 [5]. The inner capsid is made of the subcore, formed by VP3, and the intermediate layer, constituted by VP7, along with three minor structural proteins with enzymatic activities: VP1 (RNA-dependent RNA polymerase), VP4 (capping enzyme), and VP6 (helicase) [4,6,7,8,9][4][6][7][8][9].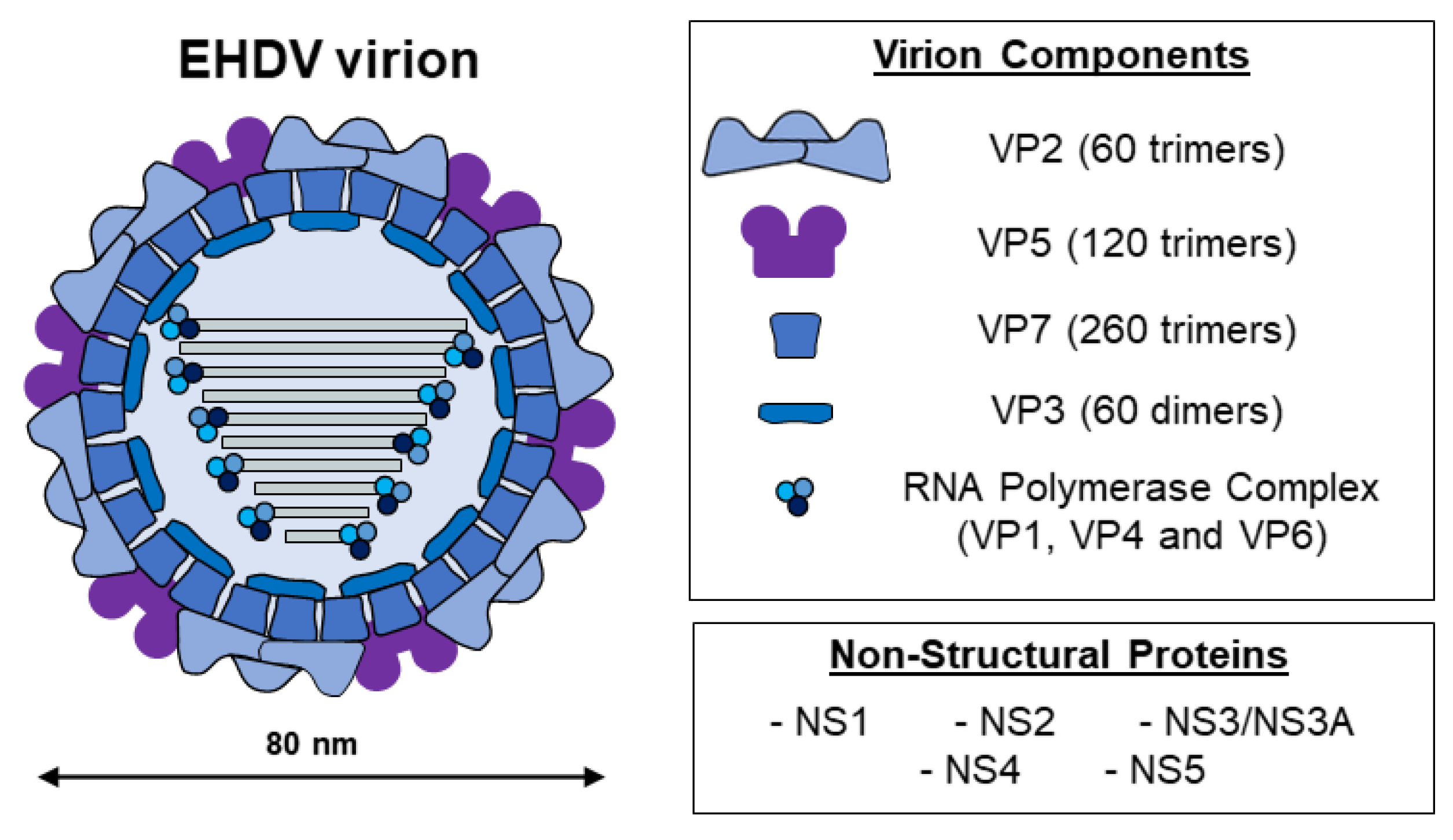
3. Changes in EHDV Epidemiology
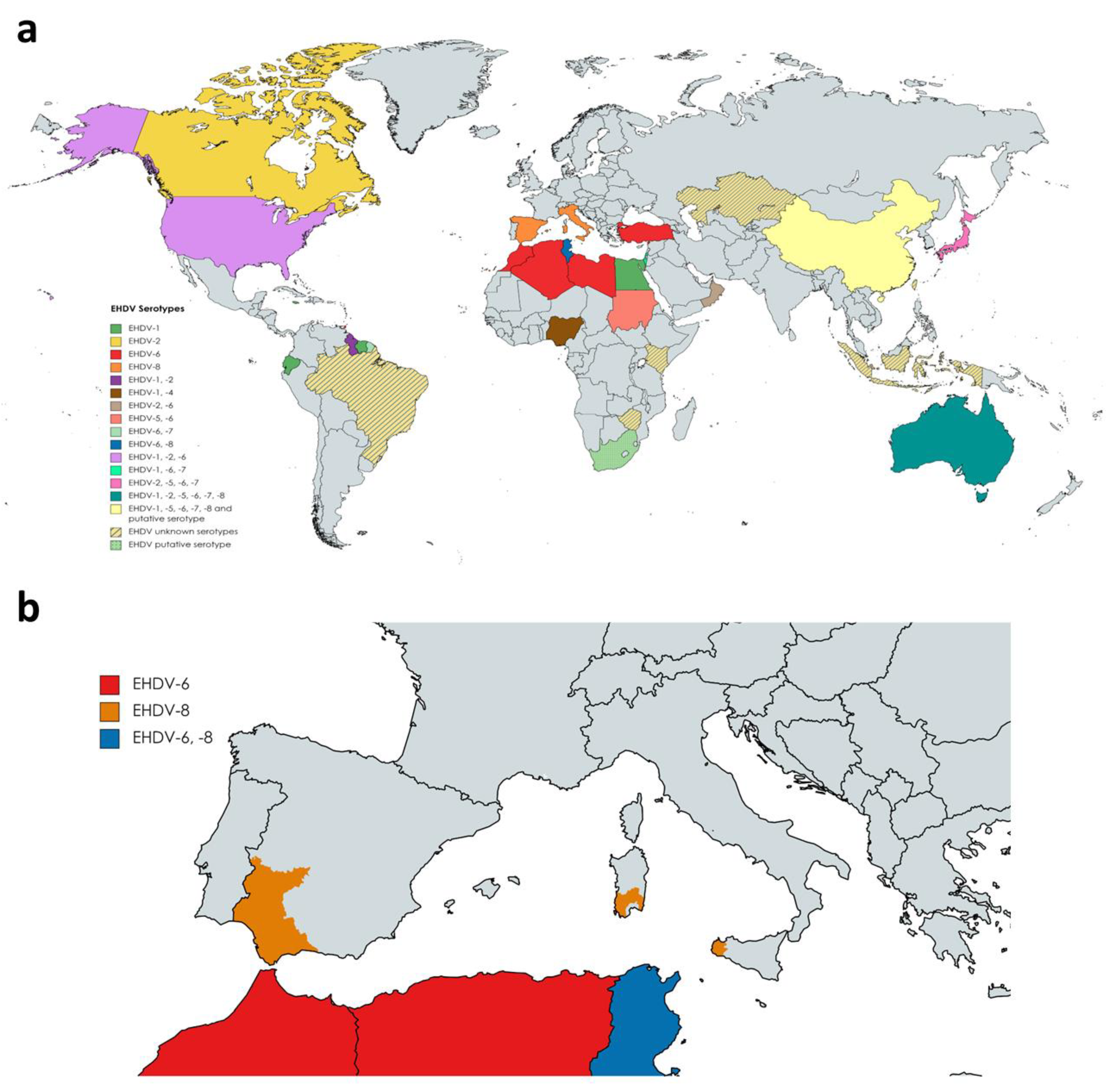
To date, seven serotypes of EHDV have been identified, named as 1–2 and 4–8, designated based on extensive phylogenetic studies, sequencing data, and cross-neutralization assays [10]. Genetic analyses demonstrated that previously identified serotype 3 [34][32] (Nigerian strain Ib Ar 22619) was serotype 1 [10]. EHDV has been isolated in North and South America, Africa, Asia, the Middle East, and Oceania (Figure 2a). To date, it is endemic in parts of North America, Australia, and certain countries of Asia and Africa [35][33]. EHDV was first detected in the USA in 1955 when WTD were severely affected showing high mortality [36][34]. Among the seven serotypes proposed, EHDV-1, 2, and 6 have been reported to be present in North America, where WTD is the most severely affected host and the scale of individual outbreaks increased with time [37,38,39][35][36][37]. In Australia, six out of seven serotypes (EHDV-1, -2, -5, -6, -7, and -8) have been detected over the years [35,40][33][38]. Globally, the presence of EHDV has been noted in Japan (serotypes 2 and 7, and serotypes 5 and 6 recently isolated from Culicoides insect vectors), China (serotypes 1, 5, 6, 7, 8), Morocco (serotype 6), Algeria (serotype 6), Libya (serotype 6), Turkey (serotype 6), Tunisia (serotypes 6 and 8), Egypt (serotype 1), Oman (serotype 2 and 6), Sudan (serotype 5 and 6), Nigeria (serotypes 1 and 4), the island of Mayotte (serotype 6), French Guiana (serotypes 6 and 7), Ecuador (serotype 1), Trinidad (serotype 6) and Israel (serotypes 1, 6 and 7) [41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59][39][40][41][42][43][44][45][46][47][48][49][50][51][52][53][54][55][56][57]. Genome detection or serological evidences also indicate the presence of EHDV (unknown serotypes) in Indonesia, Taiwan, Zimbabwe, Kenya, Kazakhstan, and Brazil [60,61,62,63,64][58][59][60][61][62]. Nucleotide sequencing and neutralization tests suggest novel strains of EHDV identified in South Africa and China as new putative serotypes [65,66][63][64].
Outbreaks during 2020 in Turkey and North Africa were originated by serotype 6, while serotype 8 was the causative virus of several outbreaks in cattle in Tunisia during 2021 [50][48]. This was the first evidence of EHDV-8 circulation since 1982, when this serotype was isolated in Australia. In Europe, there was no evidence of the presence of EHDV. However, it emerged on the continent for the first time in October 2022. After BTV-like clinical signs were noticed in some animals, EHDV was identified as the causative agent of cattle disease outbreaks in Sicily and southwestern Sardinia (Figure 2b). Subsequently, it was confirmed that these outbreaks were caused by EHDV-8, an identical strain to the one circulating in Tunisia between 2021 and 2022, pointing to North Africa as the direct origin [80][65]. Consecutively, EHDV outbreaks were detected in southern Spain, with EHDV serotype 8 confirmed and notified to the WOAH as the causative agent (Figure 2b).
4. Disease and Pathology
5. Experimental Animal Models of EHDV
5.1. White-Tailed Deer (WTD) and Other Cervid Species
As the global incidence of EHD is constantly increasing, the study of pathogenesis, transmission, and diagnosis as well as the evaluation of vaccine candidates in natural wildlife hosts is a key issue. In this sense, cervids have been used for EHDV study in natural hosts (Table 1). Different species of cervids, including red, fallow, roe, and muntjac deer, were experimentally infected with the New Jersey strain of EHDV-1, which is highly virulent in WTD. In contrast, these animals did not display any signs of disease, although productive infection could be detected [123][87]. Among the species of cervids susceptible to EHDV infection, WTD stand out as the most affected host of EHDV. Experimental infection of WTD with EHDV followed the first outbreak detected in the USA. WTD inoculated with EHDV-1 displayed severe illness in most cases, resembling the clinical disease observed in nature and displaying gross and histopathological lesions [124,125][88][89]. A high mortality among infected WTD was recorded [125][89]. In general, experimental infection of WTD with EHDV induces acute disease leading to high mortality rates, independent from the age or sex of the infected animals. WTD infected with the reassortant North American EHDV-6 resembled infection and clinical disease caused by the US strains of EHDV-1 and EHDV-2. Researchers also characterized virological parameters of EHDV infection, detecting viraemia from the third day post-inoculation until more than two weeks later (viraemia was detectable for more than seven weeks) in surviving animals, even in presence of high nAb titers. Viral RNA was detected in tissue samples of organs that presented macro- and microscopic lesions [126][90]. A field isolate of EHDV-7 that caused intense and widespread epizootic in domestic cattle in Israel was also assessed to determine whether WTD was susceptible to infection. This virus strain led to a clinical disease identical to that observed in experimental infections with North American isolates, and virological parameters were similar to those of EHDV-6 infected WTD [127][91]. The fact that “exotic” strains of EHDV can productively infect WTD inducing fatal clinical disease highlights the marked susceptibility of WTD to EHDV infection. Nonetheless, it is important to note that not all subspecies of WTD share the same susceptibility grade, with some of them, such as the subspecies Odocoileus virginianus texanus (which inhabits regions where EDHV has been endemic for a long period), showing innate resistance to EHDV [128][92]. Differential expression of pro-inflammatory cytokines could explain this resistance [115][81]. Host genetic factors can also influence susceptibility to EHDV infection [119][85]. This illustrates the complexity of understanding the pathogenesis and virulence of EHDV even in WTD.5.2. Cattle and Other Farm Animals
WTD and other wildlife experimental models involve limitations (reviewed in [138][93]) that are not shared by suitable traditional livestock experimental animals widely used in infectious disease research. As stated above, during recent EHDV outbreaks in North America, the Mediterranean Basin, and Reunion Island, an apparent increase in pathogenicity of EHDV in cattle raised concerns. Therefore, cattle can be considered as a more accessible and cheaper alternative to WTD for studying viral pathology and evaluating vaccine efficacy. In later studies, cattle, sheep, pigs, and goats were inoculated with the New Jersey strain of EHDV-1, which was virulent in deer [123][87]. Despite technical limitations concomitant with that time, researchers were able to observe viraemia in inoculated sheep and cattle while none of the goats or pigs were viraemic. Interestingly, the virus was recovered from the vulva of a recently lambed viraemic sheep. Clinical disease was not observed in any animal [123][87]. Similarly, other researchers observed that inoculation of cattle with the EHDV-1 New Jersey strain did not induce clinical disease, but the virus could be isolated from day 9 to day 23 post-inoculation and viraemia was detected by gel-based reverse transcriptase-PCR between days 3 and 28 post-infection. Similar results were observed for cattle infected with the Alberta strain of EHDV-2 [139][94]. In another work, subsequent inoculation of cattle with two US isolates led to transient viraemia as well as the induction of a neutralizing immune response in absence of clinical disease [140][95].5.3. Mouse Models
Availability of appropriate laboratory animal models is a major concern when studying disease pathogenesis and developing efficient and safe therapies against viral diseases. Mouse models are a reliable tool that reproduce or, at least, partially mimic the disease pathogenesis in a variety of cases [148][96]. For vaccine development, utilization of valid mouse models endorses the basis of every traditional vaccine development procedure as it implies several advantages, such as reduction of costs and time, facility to handle and accessibility of a huge number of optimal reagents. When considering vaccine evaluation against veterinary diseases, mouse models offer a more accessible and affordable animal housing compared to natural hosts. Immunocompromised mouse models, like mice deficient in the type I IFN (IFN-α/β) receptor (IFNAR(−/−)), have been extensively used for vaccine efficacy assessment. The IFNAR(−/−) knock-out receptor mouse model has been employed to study viral infection, disease, pathogenesis and vaccine testing against a plethora of viral diseases [148][96]. This laboratory animal model has been exploited for extensive study of BTV and AHSV [149,150,151][97][98][99]. Infection of IFNAR(−/−) mice with BTV and AHSV reproduces the pathology observed in natural hosts of these viruses.6. Classic and Novel Vaccine Approaches against EHDV
Vaccination entails the most effective countermeasure to successfully contain several human and veterinary viral diseases. In the case of EHDV, vaccines based on conventional approaches have been developed and their commercialization has been circumscribed to regions where the virus has circulated causing a significant economic impact. In Japan, two vaccines against EHDV-2 are commercially available: a monovalent live attenuated vaccine and an inactivated bivalent vaccine (against EHDV-2 and bovine ephemeral fever, caused by bovine ephemeral fever virus). LAVs (live attenuated vaccines) against BTV are used in the United States, Turkey, the Republic of South Africa, India, and Israel, among others [161][100]. However, BTV LAVs, which show highly immunogenicity, are often associated with several drawbacks relating toanimal welfare and transmission to insect vectors (reviewed elsewhere [161][100]). For these reasons, and after being used to control several outbreaks of BTV over the years, immunization with BTV LAVs was reduced and, eventually, completely substituted by inactivated vaccines in the European Union (EU) [162][101]. Therefore, LAVs are not a recommended choice to consider in EHDV vaccination campaigns in the EU, as the possibility of virus spillover to unaffected regions through uptake and spread by midges or in-contact transmission is clear. Inactivated vaccines against BTV are produced and licensed in Europe, and, although some pitfalls exist (reviewed in [162][101]), this approach has demonstrated more than enough efficacy to control this disease [162][101]. Next-generation vaccines against EHDV must overcome inherent disadvantages of classical approaches that occur with BTV and AHSV. First, they must allow differentiation between naturally infected and vaccinated animals (DIVA), which has fundamental implications in the economic field. Second, these newly generated vaccines should induce protection against multiple EHDV serotypes, whose expansion to non-endemic latitudes is highly probable. To date, the unique vaccine candidate that has been evaluated is based on recombinant VP2 protein of EHDV-2 [164][102]. This DIVA subunit vaccine has shown promising results in terms of immunogenicity and protection in the primarily EHDV affected host, WTD, preventing it from EHDV clinical disease, infection, and viraemia. Prime-boost immunization with rVP2 of serotype 2 induced high titers (ranging from 1:240 to 1:320) of homologous nAbs in immunized WTD. After viral challenge with virulent EHDV-2, immunized animals did not display EHD-related signs of disease and showed steady rectal temperatures and peripheral lymphocyte counts. No viraemia nor RNA were detected in EHDV-target organs, and immunized WTD showed an absence of gross and histopathological lesions. Although not yet evaluated, its efficacy for avoiding transmission to the insect vector seems plausible as no RNA was detected in blood. This EHDV-2 rVP2-based vaccine is currently under field trial in the USA. Importantly, cattle immunized following the same immunization strategy also developed a potent humoral response. Not only that, but the authors also achieved the expression and purification of the rVP2 of EHDV-6, which induced high titers of homologous nAbs in cattle. In this sense, bi- or multivalent vaccines could be formulated as proposed in the study [164][102]. Different vaccine platforms widely used for novel BTV and AHSV vaccines [152,165][103][104] should be applied for generation of novel EHDV vaccines, e.g., subunit vaccine and viral vector-based vaccines.7. Conclusions
EHDV, an important arthropod-transmitted RNA virus that infects different wild and domestic ruminants, has experienced a northern spread into novel areas in the last 20 years. Global warming may result in expansion of vector species to previously vector-free regions, and in altered vector competence of some midge species. These factors and others related with human activities are likely to increase the risk of EHDV outbreaks in new territories. Worryingly, the virus has recently been detected for the first time in the European Union in October 2022. White-tailed deer are especially susceptible to severe illness caused by EHDV infection; nevertheless, disease can also occur in bovines. The increased virulence of certain EHDV strains observed in cattle and the expansion of competent vectors involved in EHDV transmission make necessary further investigation regarding the development of new diagnostic techniques, safe DIVA vaccines, and the evaluation of laboratory animal models that will facilitate the study of the protective capacity of new vaccine candidates against EHDV.References
- Huismans, H.; Bremer, C.W.; Barber, T.L. The Nucleic Acid and Proteins of Epizootic Haemorrhagic Disease Virus. Onderstepoort J. Vet. Res. 1979, 46, 95–104.
- Stewart, M.; Hardy, A.; Barry, G.; Pinto, R.M.; Caporale, M.; Melzi, E.; Hughes, J.; Taggart, A.; Janowicz, A.; Varela, M.; et al. Characterization of a Second Open Reading Frame in Genome Segment 10 of Bluetongue Virus. J. Gen. Virol. 2015, 96, 3280–3293.
- Belhouchet, M.; Mohd Jaafar, F.; Firth, A.E.; Grimes, J.M.; Mertens, P.P.C.; Attoui, H. Detection of a Fourth Orbivirus Non-Structural Protein. PLoS ONE 2011, 6, e25697.
- Mecham, J.O.; Dean, V.C. Protein Coding Assignment for the Genome of Epizootic Haemorrhagic Disease Virus. J. Gen. Virol. 1988, 69 Pt 6, 1255–1262.
- Zhang, X.; Boyce, M.; Bhattacharya, B.; Zhang, X.; Schein, S.; Roy, P.; Zhou, Z.H. Bluetongue Virus Coat Protein VP2 Contains Sialic Acid-Binding Domains, and VP5 Resembles Enveloped Virus Fusion Proteins. Proc. Natl. Acad. Sci. USA 2010, 107, 6292–6297.
- Urakawa, T.; Ritter, D.G.; Roy, P. Expression of Largest RNA Segment and Synthesis of VP1 Protein of Bluetongue Virus in Insect Cells by Recombinant Baculovirus: Association of VP1 Protein with RNA Polymerase Activity. Nucleic Acids Res. 1989, 17, 7395–7401.
- Stäuber, N.; Martinez-Costas, J.; Sutton, G.; Monastyrskaya, K.; Roy, P. Bluetongue Virus VP6 Protein Binds ATP and Exhibits an RNA-Dependent ATPase Function and a Helicase Activity That Catalyze the Unwinding of Double-Stranded RNA Substrates. J. Virol. 1997, 71, 7220–7226.
- Sutton, G.; Grimes, J.M.; Stuart, D.I.; Roy, P. Bluetongue Virus VP4 Is an RNA-Capping Assembly Line. Nat. Struct. Mol. Biol. 2007, 14, 449–451.
- Wu, W.; Roy, P. Sialic Acid Binding Sites in VP2 of Bluetongue Virus and Their Use during Virus Entry. J. Virol. 2022, 96, e0167721.
- Anthony, S.J.; Maan, S.; Maan, N.; Kgosana, L.; Bachanek-Bankowska, K.; Batten, C.; Darpel, K.E.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Outer-Coat Proteins VP2 and VP5 of Epizootic Haemorrhagic Disease Virus (EHDV): Comparison of Genetic and Serological Data to Characterise the EHDV Serogroup. Virus Res. 2009, 145, 200–210.
- Forzan, M.; Marsh, M.; Roy, P. Bluetongue Virus Entry into Cells. J. Virol. 2007, 81, 4819–4827.
- Maan, N.S.; Maan, S.; Potgieter, A.C.; Wright, I.M.; Belaganahalli, M.; Mertens, P.P.C. Development of Real-Time RT-PCR Assays for Detection and Typing of Epizootic Haemorrhagic Disease Virus. Transbound. Emerg. Dis. 2017, 64, 1120–1132.
- Sun, F.; Cochran, M.; Beckham, T.; Clavijo, A. Molecular Typing of Epizootic Hemorrhagic Disease Virus Serotypes by One-Step Multiplex RT-PCR. J. Wildl. Dis. 2014, 50, 639–644.
- Anthony, S.J.; Maan, N.; Maan, S.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Core Proteins VP1, VP3, VP4, VP6 and VP7 of Epizootic Haemorrhagic Disease Virus (EHDV). Virus Res. 2009, 145, 187–199.
- Mecham, J.O.; Stallknecht, D.; Wilson, W.C. The S7 Gene and VP7 Protein Are Highly Conserved among Temporally and Geographically Distinct American Isolates of Epizootic Hemorrhagic Disease Virus. Virus Res. 2003, 94, 129–133.
- Bréard, E.; Viarouge, C.; Donnet, F.; Sailleau, C.; Rossi, S.; Pourquier, P.; Vitour, D.; Comtet, L.; Zientara, S. Evaluation of a Commercial ELISA for Detection of Epizootic Haemorrhagic Disease Antibodies in Domestic and Wild Ruminant Sera. Transbound. Emerg. Dis. 2020, 67, 2475–2481.
- Forzan, M.; Pizzurro, F.; Zaccaria, G.; Mazzei, M.; Spedicato, M.; Carmine, I.; Salini, R.; Tolari, F.; Cerri, D.; Savini, G.; et al. Competitive Enzyme-Linked Immunosorbent Assay Using Baculovirus-Expressed VP7 for Detection of Epizootic Haemorrhagic Disease Virus (EHDV) Antibodies. J. Virol. Methods 2017, 248, 212–216.
- Serroni, A.; Ulisse, S.; Iorio, M.; Laguardia, C.; Testa, L.; Armillotta, G.; Caporale, M.; Salini, R.; Lelli, D.; Wernery, U.; et al. Development of a Competitive Enzyme-Linked Immunosorbent Assay Based on Purified Recombinant Viral Protein 7 for Serological Diagnosis of Epizootic Haemorrhagic Disease in Camels. J. Trop. Med. 2022, 2022, 5210771.
- Kusari, J.; Roy, P. Molecular and Genetic Comparisons of Two Serotypes of Epizootic Hemorrhagic Disease of Deer Virus. Am. J. Vet. Res. 1986, 47, 1713–1717.
- Nel, L.H.; Picard, L.A.; Huismans, H. A Characterization of the Nonstructural Protein from Which the Virus-Specified Tubules in Epizootic Haemorrhagic Disease Virus-Infected Cells Are Composed. Virus Res. 1991, 18, 219–230.
- Owens, R.J.; Limn, C.; Roy, P. Role of an Arbovirus Nonstructural Protein in Cellular Pathogenesis and Virus Release. J. Virol. 2004, 78, 6649–6656.
- Boyce, M.; Celma, C.C.P.; Roy, P. Bluetongue Virus Non-Structural Protein 1 Is a Positive Regulator of Viral Protein Synthesis. Virol. J. 2012, 9, 178.
- Anthony, S.J.; Maan, N.; Maan, S.; Sutton, G.; Attoui, H.; Mertens, P.P.C. Genetic and Phylogenetic Analysis of the Non-Structural Proteins NS1, NS2 and NS3 of Epizootic Haemorrhagic Disease Virus (EHDV). Virus Res. 2009, 145, 211–219.
- Theron, J.; Huismans, H.; Nel, L.H. Identification of a Short Domain within the Non-Structural Protein NS2 of Epizootic Haemorrhagic Disease Virus That Is Important for Single Strand RNA-Binding Activity. J. Gen. Virol. 1996, 77, 129–137.
- Uitenweerde, J.M.; Theron, J.; Stoltz, M.A.; Huismans, H. The Multimeric Nonstructural NS2 Proteins of Bluetongue Virus, African Horsesickness Virus, and Epizootic Hemorrhagic Disease Virus Differ in Their Single-Stranded RNA-Binding Ability. Virology 1995, 209, 624–632.
- Theron, J.; Huismans, H.; Nel, L.H. Site-Specific Mutations in the NS2 Protein of Epizootic Haemorrhagic Disease Virus Markedly Affect the Formation of Cytoplasmic Inclusion Bodies. Arch. Virol. 1996, 141, 1143–1151.
- Jensen, M.J.; Wilson, W.C. A Model for the Membrane Topology of the NS3 Protein as Predicted from the Sequence of Segment 10 of Epizootic Haemorrhagic Disease Virus Serotype 1. Arch. Virol. 1995, 140, 799–805.
- Hyatt, A.D.; Zhao, Y.; Roy, P. Release of Bluetongue Virus-like Particles from Insect Cells Is Mediated by BTV Nonstructural Protein NS3/NS3A. Virology 1993, 193, 592–603.
- Jensen, M.J.; Cheney, I.W.; Thompson, L.H.; Mecham, J.O.; Wilson, W.C.; Yamakawa, M.; Roy, P.; Gorman, B.M. The Smallest Gene of the Orbivirus, Epizootic Hemorrhagic Disease, Is Expressed in Virus-Infected Cells as Two Proteins and the Expression Differs from That of the Cognate Gene of Bluetongue Virus. Virus Res. 1994, 32, 353–364.
- Fablet, A.; Kundlacz, C.; Dupré, J.; Hirchaud, E.; Postic, L.; Sailleau, C.; Bréard, E.; Zientara, S.; Vitour, D.; Caignard, G. Comparative Virus-Host Protein Interactions of the Bluetongue Virus NS4 Virulence Factor. Viruses 2022, 14, 182.
- Ratinier, M.; Shaw, A.E.; Barry, G.; Gu, Q.; Di Gialleonardo, L.; Janowicz, A.; Varela, M.; Randall, R.E.; Caporale, M.; Palmarini, M. Bluetongue Virus NS4 Protein Is an Interferon Antagonist and a Determinant of Virus Virulence. J. Virol. 2016, 90, 5427–5439.
- Campbell, C.H.; George, T.D.S. A Preliminary Report of a Comparison of Epizootic Haemorrhagic Disease Viruses from Australia with Others from North America, Japan and Nigeria. Aust. Vet. J. 1986, 63, 233.
- Noronha, L.E.; Cohnstaedt, L.W.; Richt, J.A.; Wilson, W.C. Perspectives on the Changing Landscape of Epizootic Hemorrhagic Disease Virus Control. Viruses 2021, 13, 2268.
- Shope, R.E.; Lester, G.M.; Robert, M. Deer Mortality—Epizootic Hemorrhagic Disease of Deer. N. J. Outdoors 1955, 6, 17–21.
- Ruder, M.G.; Lysyk, T.J.; Stallknecht, D.E.; Foil, L.D.; Johnson, D.J.; Chase, C.C.; Dargatz, D.A.; Gibbs, E.P.J. Transmission and Epidemiology of Bluetongue and Epizootic Hemorrhagic Disease in North America: Current Perspectives, Research Gaps, and Future Directions. Vector Borne Zoonotic Dis. 2015, 15, 348–363.
- Ruder, M.G.; Johnson, D.; Ostlund, E.; Allison, A.B.; Kienzle, C.; Phillips, J.E.; Poulson, R.L.; Stallknecht, D.E. The First 10 Years (2006-15) of Epizootic Hemorrhagic Disease Virus Serotype 6 in the USA. J. Wildl. Dis. 2017, 53, 901–905.
- Schirtzinger, E.E.; Jasperson, D.C.; Ruder, M.G.; Stallknecht, D.E.; Chase, C.C.L.; Johnson, D.J.; Ostlund, E.N.; Wilson, W.C. Evaluation of 2012 US EHDV-2 Outbreak Isolates for Genetic Determinants of Cattle Infection. J. Gen. Virol. 2019, 100, 556–567.
- Weir, R.P.; Harmsen, M.B.; Hunt, N.T.; Blacksell, S.D.; Lunt, R.A.; Pritchard, L.I.; Newberry, K.M.; Hyatt, A.D.; Gould, A.R.; Melville, L.F. EHDV-1, a New Australian Serotype of Epizootic Haemorrhagic Disease Virus Isolated from Sentinel Cattle in the Northern Territory. Vet. Microbiol. 1997, 58, 135–143.
- Ahmed, S.; Mahmoud, M.A.E.-F.; Viarouge, C.; Sailleau, C.; Zientara, S.; Breard, E. Presence of Bluetongue and Epizootic Hemorrhagic Disease Viruses in Egypt in 2016 and 2017. Infect. Genet. Evol. 2019, 73, 221–226.
- Viarouge, C.; Lancelot, R.; Rives, G.; Bréard, E.; Miller, M.; Baudrimont, X.; Doceul, V.; Vitour, D.; Zientara, S.; Sailleau, C. Identification of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus Serotypes in French Guiana in 2011 and 2012. Vet. Microbiol. 2014, 174, 78–85.
- Qi, Y.; Wang, F.; Chang, J.; Zhang, Y.; Zhu, J.; Li, H.; Yu, L. Identification and Complete-Genome Phylogenetic Analysis of an Epizootic Hemorrhagic Disease Virus Serotype 7 Strain Isolated in China. Arch. Virol. 2019, 164, 3121–3126.
- Yadin, H.; Brenner, J.; Bumbrov, V.; Oved, Z.; Stram, Y.; Klement, E.; Perl, S.; Anthony, S.; Maan, S.; Batten, C.; et al. Epizootic Haemorrhagic Disease Virus Type 7 Infection in Cattle in Israel. Vet. Rec. 2008, 162, 53–56.
- Temizel, E.M.; Yesilbag, K.; Batten, C.; Senturk, S.; Maan, N.S.; Mertens, P.P.C.; Batmaz, H. Epizootic Hemorrhagic Disease in Cattle, Western Turkey. Emerg. Infect. Dis. 2009, 15, 317–319.
- Verdezoto, J.; Breard, E.; Viarouge, C.; Quenault, H.; Lucas, P.; Sailleau, C.; Zientara, S.; Augot, D.; Zapata, S. Novel Serotype of Bluetongue Virus in South America and First Report of Epizootic Haemorrhagic Disease Virus in Ecuador. Transbound. Emerg. Dis. 2018, 65, 244–247.
- Rajko-Nenow, P.; Brown-Joseph, T.; Tennakoon, C.; Flannery, J.; Oura, C.A.L.; Batten, C. Detection of a Novel Reassortant Epizootic Hemorrhagic Disease Virus Serotype 6 in Cattle in Trinidad, West Indies, Containing Nine RNA Segments Derived from Exotic EHDV Strains with an Australian Origin. Infect. Genet. Evol. 2019, 74, 103931.
- Mohammed, M.E.; Aradaib, I.E.; Mukhtar, M.M.; Ghalib, H.W.; Riemann, H.P.; Oyejide, A.; Osburn, B.I. Application of Molecular Biological Techniques for Detection of Epizootic Hemorrhagic Disease Virus (EHDV-318) Recovered from a Sentinel Calf in Central Sudan. Vet. Microbiol. 1996, 52, 201–208.
- Aradaib, I.E.; Mohammed, M.E.; Mukhtar, M.M.; Ghalib, H.W.; Osburn, B.I. Serogrouping and Topotyping of Sudanese and United States Strains of Epizootic Hemorrhagic Disease Virus Using PCR. Comp. Immunol. Microbiol. Infect. Dis. 1997, 20, 211–218.
- Sghaier, S.; Sailleau, C.; Marcacci, M.; Thabet, S.; Curini, V.; Ben Hassine, T.; Teodori, L.; Portanti, O.; Hammami, S.; Jurisic, L.; et al. Epizootic Haemorrhagic Disease Virus Serotype 8 in Tunisia, 2021. Viruses 2022, 15, 16.
- Duan, Y.; Yang, Z.; Zhu, P.; Xiao, L.; Li, Z.; Li, Z.; Li, L.; Zhu, J. A Serologic Investigation of Epizootic Hemorrhagic Disease Virus in China between 2014 and 2019. Virol. Sin. 2022, 37, 513–520.
- Mejri, S.; Dhaou, S.B.; Jemli, M.; Bréard, E.; Sailleau, C.; Sghaier, S.; Zouari, M.; Lorusso, A.; Savini, G.; Zientara, S.; et al. Epizootic Haemorrhagic Disease Virus Circulation in Tunisia. Vet. Ital. 2018, 54, 87–90.
- Yamamoto, K.; Hiromatsu, R.; Kaida, M.; Kato, T.; Yanase, T.; Shirafuji, H. Isolation of Epizootic Hemorrhagic Disease Virus Serotype 7 from Cattle Showing Fever in Japan in 2016 and Improvement of a Reverse Transcription-Polymerase Chain Reaction Assay to Detect Epizootic Hemorrhagic Disease Virus. J. Vet. Med. Sci. 2021, 83, 1378–1388.
- Mahmoud, A.; Danzetta, M.L.; di Sabatino, D.; Spedicato, M.; Alkhatal, Z.; Dayhum, A.; Tolari, F.; Forzan, M.; Mazzei, M.; Savini, G. First Seroprevalence Investigation of Epizootic Haemorrhagic Disease Virus in Libya. Open Vet. J. 2021, 11, 301–308.
- Brown-Joseph, T.; Rajko-Nenow, P.; Hicks, H.; Sahadeo, N.; Harrup, L.E.; Carrington, C.V.; Batten, C.; Oura, C.A.L. Identification and Characterization of Epizootic Hemorrhagic Disease Virus Serotype 6 in Cattle Co-Infected with Bluetongue Virus in Trinidad, West Indies. Vet. Microbiol. 2019, 229, 1–6.
- Dommergues, L.; Viarouge, C.; Métras, R.; Youssouffi, C.; Sailleau, C.; Zientara, S.; Cardinale, E.; Cêtre-Sossah, C. Evidence of Bluetongue and Epizootic Haemorrhagic Disease Circulation on the Island of Mayotte. Acta Trop. 2019, 191, 24–28.
- Wilson, W.C.; Ruder, M.G.; Klement, E.; Jasperson, D.C.; Yadin, H.; Stallknecht, D.E.; Mead, D.G.; Howerth, E. Genetic Characterization of Epizootic Hemorrhagic Disease Virus Strains Isolated from Cattle in Israel. J. Gen. Virol. 2015, 96, 1400–1410.
- Murota, K.; Ishii, K.; Mekaru, Y.; Araki, M.; Suda, Y.; Shirafuji, H.; Kobayashi, D.; Isawa, H.; Yanase, T. Isolation of Culicoides- and Mosquito-Borne Orbiviruses in the Southwestern Islands of Japan between 2014 and 2019. Vector Borne Zoonotic Dis. 2021, 21, 796–808.
- Al-Busaidy, S.M.; Mellor, P.S. Epidemiology of Bluetongue and Related Orbiviruses in the Sultanate of Oman. Epidemiol. Infect. 1991, 106, 167–178.
- Gordon, S.J.G.; Bolwell, C.; Rogers, C.W.; Musuka, G.; Kelly, P.; Guthrie, A.; Mellor, P.S.; Hamblin, C. A Serosurvey of Bluetongue and Epizootic Haemorrhagic Disease in a Convenience Sample of Sheep and Cattle Herds in Zimbabwe. Onderstepoort J. Vet. Res. 2017, 84, 1505.
- Toye, P.G.; Batten, C.A.; Kiara, H.; Henstock, M.R.; Edwards, L.; Thumbi, S.; Poole, E.J.; Handel, I.G.; Bronsvoort, B.M.d.C.; Hanotte, O.; et al. Bluetongue and Epizootic Haemorrhagic Disease Virus in Local Breeds of Cattle in Kenya. Res. Vet. Sci. 2013, 94, 769–773.
- Lundervold, M.; Milner-Gulland, E.J.; O’Callaghan, C.J.; Hamblin, C.; Corteyn, A.; Macmillan, A.P. A Serological Survey of Ruminant Livestock in Kazakhstan during Post-Soviet Transitions in Farming and Disease Control. Acta Vet. Scand. 2004, 45, 211.
- Favero, C.M.; Matos, A.C.D.; Campos, F.S.; Cândido, M.V.; Costa, É.A.; Heinemann, M.B.; Barbosa-Stancioli, E.F.; Lobato, Z.I.P. Epizootic Hemorrhagic Disease in Brocket Deer, Brazil. Emerg. Infect. Dis. 2013, 19, 346–348.
- Inaba, Y. Ibaraki Disease and Its Relationship to Bluetongue. Aust. Vet. J. 1975, 51, 178–185.
- Yang, H.; Li, Z.; Wang, J.; Li, Z.; Yang, Z.; Liao, D.; Zhu, J.; Li, H. Novel Serotype of Epizootic Hemorrhagic Disease Virus, China. Emerg. Infect. Dis. 2020, 26, 3081–3083.
- Wright, I.M. Serological and Genetic Characterisation of Putative New Serotypes of Bluetongue Virus and Epizootic Haemorrhagic Disease Virus Isolated from an Alpaca. Master’s Thesis, North-West University, Potchefstroom, South Africa, 2014.
- Lorusso, A.; Cappai, S.; Loi, F.; Pinna, L.; Ruiu, A.; Puggioni, G.; Guercio, A.; Purpari, G.; Vicari, D.; Sghaier, S.; et al. First Detection of Epizootic Haemorrhagic Disease Virus in the European Union, Italy-2022. bioRxiv 2022.
- Allison, A.B.; Goekjian, V.H.; Potgieter, A.C.; Wilson, W.C.; Johnson, D.J.; Mertens, P.P.C.; Stallknecht, D.E. Detection of a Novel Reassortant Epizootic Hemorrhagic Disease Virus (EHDV) in the USA Containing RNA Segments Derived from Both Exotic (EHDV-6) and Endemic (EHDV-2) Serotypes. J. Gen. Virol. 2010, 91, 430–439.
- Allison, A.B.; Holmes, E.C.; Potgieter, A.C.; Wright, I.M.; Sailleau, C.; Breard, E.; Ruder, M.G.; Stallknecht, D.E. Segmental Configuration and Putative Origin of the Reassortant Orbivirus, Epizootic Hemorrhagic Disease Virus Serotype 6, Strain Indiana. Virology 2012, 424, 67–75.
- Howerth, E.W.; Stallknecht, D.E.; Kirkland, P.D. Bluetongue, Epizootic Hemorrhagic Disease, and Other Orbivirus-Related Diseases. In Infectious Diseases of Wild Mammals; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2001; pp. 77–97. ISBN 978-0-470-34488-0.
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Howerth, E.W.; Murphy, M.; Stallknecht, D.E. Cross-Protection between Epizootic Hemorrhagic Disease Virus Serotypes 1 and 2 in White-Tailed Deer. J. Wildl. Dis. 2002, 38, 720–728.
- Brodie, S.J.; Bardsley, K.D.; Diem, K.; Mecham, J.O.; Norelius, S.E.; Wilson, W.C. Epizootic Hemorrhagic Disease: Analysis of Tissues by Amplification and in Situ Hybridization Reveals Widespread Orbivirus Infection at Low Copy Numbers. J. Virol. 1998, 72, 3863–3871.
- Noon, T.H.; Wesche, S.L.; Heffelfinger, J.; Fuller, A.; Bradley, G.A.; Reggiardo, C. Hemorrhagic Disease in Deer in Arizona. J. Wildl. Dis 2002, 38, 177–181.
- Roug, A.; Shannon, J.; Hersey, K.; Heaton, W.; van Wettere, A. Investigation into Causes of Antler Deformities in Mule Deer (Odocoileus hemionus) Bucks in Southern Utah, USA. J. Wildl. Dis. 2022, 58, 222–227.
- Raabis, S.M.; Byers, S.R.; Han, S.; Callan, R.J. Epizootic Hemorrhagic Disease in a Yak. Can. Vet. J. 2014, 55, 369–372.
- Kedmi, M.; Levi, S.; Galon, N.; Bomborov, V.; Yadin, H.; Batten, C.; Klement, E. No Evidence for Involvement of Sheep in the Epidemiology of Cattle Virulent Epizootic Hemorrhagic Disease Virus. Vet. Microbiol. 2011, 148, 408–412.
- Nol, P.; Kato, C.; Reeves, W.K.; Rhyan, J.; Spraker, T.; Gidlewski, T.; VerCauteren, K.; Salman, M. Epizootic Hemorrhagic Disease Outbreak in a Captive Facility Housing White-Tailed Deer (Odocoileus virginianus), Bison (Bison bison), Elk (Cervus elaphus), Cattle (Bos taurus), and Goats (Capra hircus) in Colorado, U.S.A. J. Zoo Wildl. Med. 2010, 41, 510–515.
- Mills, M.K.; Ruder, M.G.; Nayduch, D.; Michel, K.; Drolet, B.S. Dynamics of Epizootic Hemorrhagic Disease Virus Infection within the Vector, Culicoides Sonorensis (Diptera: Ceratopogonidae). PLoS ONE 2017, 12, e0188865.
- Darpel, K.E.; Langner, K.F.A.; Nimtz, M.; Anthony, S.J.; Brownlie, J.; Takamatsu, H.-H.; Mellor, P.S.; Mertens, P.P.C. Saliva Proteins of Vector Culicoides Modify Structure and Infectivity of Bluetongue Virus Particles. PLoS ONE 2011, 6, e17545.
- McLaughlin, B.E.; DeMaula, C.D.; Wilson, W.C.; Boyce, W.M.; MacLachlan, N.J. Replication of Bluetongue Virus and Epizootic Hemorrhagic Disease Virus in Pulmonary Artery Endothelial Cells Obtained from Cattle, Sheep, and Deer. Am. J. Vet. Res. 2003, 64, 860–865.
- Sharma, P.; Stallknecht, D.E.; Howerth, E.W. Epizootic Haemorrhagic Disease Virus Induced Apoptosis in Bovine Carotid Artery Endothelium Is P53 Independent. Vet. Ital. 2016, 52, 363–368.
- Shai, B.; Schmukler, E.; Yaniv, R.; Ziv, N.; Horn, G.; Bumbarov, V.; Yadin, H.; Smorodinsky, N.I.; Bacharach, E.; Pinkas-Kramarski, R.; et al. Epizootic Hemorrhagic Disease Virus Induces and Benefits from Cell Stress, Autophagy, and Apoptosis. J. Virol. 2013, 87, 13397–13408.
- Sharma, P.; Stallknecht, D.E.; Murphy, M.D.; Howerth, E.W. Expression of Interleukin-1 Beta and Interleukin-6 in White-Tailed Deer Infected with Epizootic Hemorrhagic Disease Virus. Vet. Ital. 2015, 51, 283–288.
- Howerth, E.W. Cytokine Release and Endothelial Dysfunction: A Perfect Storm in Orbivirus Pathogenesis. Vet. Ital. 2015, 51, 275–281.
- Sánchez-Cordón, P.J.; Pedrera, M.; Risalde, M.A.; Molina, V.; Rodríguez-Sánchez, B.; Núñez, A.; Sánchez-Vizcaíno, J.M.; Gómez-Villamandos, J.C. Potential Role of Proinflammatory Cytokines in the Pathogenetic Mechanisms of Vascular Lesions in Goats Naturally Infected with Bluetongue Virus Serotype 1. Transbound. Emerg. Dis. 2013, 60, 252–262.
- Quist, C.F.; Howerth, E.W.; Stallknecht, D.E.; Brown, J.; Pisell, T.; Nettles, V.F. Host Defense Responses Associated with Experimental Hemorrhagic Disease in White-Tailed Deer. J. Wildl. Dis. 1997, 33, 584–599.
- Wessels, J.E.; Ishida, Y.; Rivera, N.A.; Stirewalt, S.L.; Brown, W.M.; Novakofski, J.E.; Roca, A.L.; Mateus-Pinilla, N.E. The Impact of Variation in the Toll-like Receptor 3 Gene on Epizootic Hemorrhagic Disease in Illinois Wild White-Tailed Deer (Odocoileus virginianus). Genes 2023, 14, 426.
- Stallknecht, D.E.; Howerth, E.W.; Kellogg, M.L.; Quist, C.F.; Pisell, T. In Vitro Replication of Epizootic Hemorrhagic Disease and Bluetongue Viruses in White-Tailed Deer Peripheral Blood Mononuclear Cells and Virus-Cell Association during in Vivo Infections. J. Wildl. Dis. 1997, 33, 574–583.
- Gibbs, E.P.J.; Lawman, M.J.P. Infection of British Deer and Farm Animals with Epizootic Haemorrhagic Disease of Deer Virus. J. Comp. Pathol. 1977, 87, 335–343.
- Fletch, A.L.; Karstad, L.H. Studies on the Pathogenesis of Experimental Epizootic Hemorrhagic Disease of White-Tailed Deer. Can. J. Comp. Med. 1971, 35, 224–229.
- Shope, R.E.; Macnamara, L.G.; Mangold, R. A virus-induced epizootic hemorrhagic disease of the virginia white-tailed deer (Odocoileus virginianus). J. Exp. Med. 1960, 111, 155–170.
- Ruder, M.G.; Stallknecht, D.E.; Allison, A.B.; Mead, D.G.; Carter, D.L.; Howerth, E.W. Host and Potential Vector Susceptibility to an Emerging Orbivirus in the United States: Epizootic Hemorrhagic Disease Virus Serotype 6. Vet. Pathol. 2016, 53, 574–584.
- Ruder, M.G.; Allison, A.B.; Stallknecht, D.E.; Mead, D.G.; McGraw, S.M.; Carter, D.L.; Kubiski, S.V.; Batten, C.A.; Klement, E.; Howerth, E.W. Susceptibility of White-Tailed Deer (Odocoileus virginianus) to Experimental Infection with Epizootic Hemorrhagic Disease Virus Serotype 7. J. Wildl. Dis. 2012, 48, 676–685.
- Gaydos, J.K.; Davidson, W.R.; Elvinger, F.; Mead, D.G.; Howerth, E.W.; Stallknecht, D.E. Innate Resistance to Epizootic Hemorrhagic Disease in White-Tailed Deer. J. Wildl. Dis. 2002, 38, 713–719.
- Palmer, M.V.; Cox, R.J.; Waters, W.R.; Thacker, T.C.; Whipple, D.L. Using White-Tailed Deer (Odocoileus virginianus) in Infectious Disease Research. J. Am. Assoc. Lab. Anim. Sci. 2017, 56, 350–360.
- Aradaib, I.E.; Sawyer, M.M.; Osburn, B.I. Experimental Epizootic Hemorrhagic Disease Virus Infection in Calves: Virologic and Serologic Studies. J. Vet. Diagn. Investig. 1994, 6, 489–492.
- Bowen, R.A. Serologic Responses of Calves to Sequential Infections with Epizootic Hemorrhagic Disease Virus Serotypes. Am. J. Vet. Res. 1987, 48, 1449–1452.
- Marín-Lopez, A.; Calvo-Pinilla, E.; Moreno, S.; Utrilla-Trigo, S.; Nogales, A.; Brun, A.; Fikrig, E.; Ortego, J. Modeling Arboviral Infection in Mice Lacking the Interferon Alpha/Beta Receptor. Viruses 2019, 11, 35.
- Calvo-Pinilla, E.; Rodríguez-Calvo, T.; Anguita, J.; Sevilla, N.; Ortego, J. Establishment of a Bluetongue Virus Infection Model in Mice That Are Deficient in the Alpha/Beta Interferon Receptor. PLoS ONE 2009, 4, e5171.
- Marín-López, A.; Bermúdez, R.; Calvo-Pinilla, E.; Moreno, S.; Brun, A.; Ortego, J. Pathological Characterization of IFNAR(-/-) Mice Infected with Bluetongue Virus Serotype 4. Int. J. Biol. Sci. 2016, 12, 1448–1460.
- Castillo-Olivares, J.; Calvo-Pinilla, E.; Casanova, I.; Bachanek-Bankowska, K.; Chiam, R.; Maan, S.; Nieto, J.M.; Ortego, J.; Mertens, P.P.C. A Modified Vaccinia Ankara Virus (MVA) Vaccine Expressing African Horse Sickness Virus (AHSV) VP2 Protects against AHSV Challenge in an IFNAR−/− Mouse Model. PLoS ONE 2011, 6, e16503.
- McVey, D.S.; MacLachlan, N.J. Vaccines for Prevention of Bluetongue and Epizootic Hemorrhagic Disease in Livestock: A North American Perspective. Vector Borne Zoonotic Dis. 2015, 15, 385–396.
- van Rijn, P.A. Prospects of Next-Generation Vaccines for Bluetongue. Front. Vet. Sci. 2019, 6, 407.
- Sunwoo, S.Y.; Noronha, L.E.; Morozov, I.; Trujillo, J.D.; Kim, I.J.; Schirtzinger, E.E.; Faburay, B.; Drolet, B.S.; Urbaniak, K.; McVey, D.S.; et al. Evaluation of A Baculovirus-Expressed VP2 Subunit Vaccine for the Protection of White-Tailed Deer (Odocoileus virginianus) from Epizootic Hemorrhagic Disease. Vaccines 2020, 8, 59.
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Calvo-Pinilla, E.; Moreno, S.; Nogales, A.; Ortego, J.; Marín-López, A. Viral Vector Vaccines against Bluetongue Virus. Microorganisms 2021, 9, 42.
- Jiménez-Cabello, L.; Utrilla-Trigo, S.; Barreiro-Piñeiro, N.; Pose-Boirazian, T.; Martínez-Costas, J.; Marín-López, A.; Ortego, J. Nanoparticle- and Microparticle-Based Vaccines against Orbiviruses of Veterinary Importance. Vaccines 2022, 10, 1124.