 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Honghao Zhang | + 1106 word(s) | 1106 | 2020-11-04 07:13:46 | | | |
| 2 | Vicky Zhou | -120 word(s) | 986 | 2021-02-18 02:19:14 | | | | |
| 3 | Honghao Zhang | + 120 word(s) | 1106 | 2021-02-20 16:24:28 | | |
Video Upload Options
Ellis-van Creveld syndrome is a rare, recessive genetic disorder. The typical signs in affected patients are dwarfism, polydactyly, dysmorphic faces and dental anomalies.
- Introduction
First described in 1940, Ellis-van Creveld syndrome (EVC; MIM ID #225500) is a rare, recessive congenital disorder that results in a type of disproportionate dwarfism [1]. Affected individuals represent approximately 1 in every 60,000–200,000 live births with higher occurrence reported in offspring of consanguineous (e.g., first cousin) unions or monotypic populations [2,3]. In addition to short stature, other conspicuous signs of EVC include extra digits (i.e., polydactyly), dysmorphic faces, and dental anomalies [4]. While none of these directly confer shortened longevity, approximately 60% of patients suffer from underlying cardiac conditions, indicating widespread developmental effects.
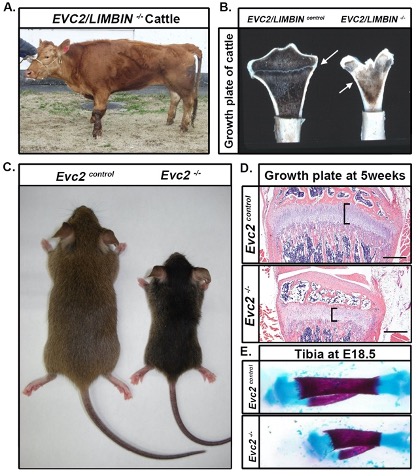
Figure 1. LIMBIN mutant cattle and Evc2/Limbin mutant mice share similar dwarfism. (A) A LIMBIN mutant cattle bears apparent short legs. (B) LIMBIN mutant cattle have decreased size of growth plate (marked in white arrow) in appendicular bones. (C) Evc2/Limbin mutant mice with littermate controls exhibit a smaller body size at 5 weeks old. (D) The growth plate of tibia from Evc2/Limbin mutant mice demonstrate shorter and disorganized structure at 5 weeks old compared with control mice. Bars indicate 200 um. (E) Tibiae from E18.5 mouse embryos were stained with alcian blue for cartilage and alizarin red for bone. Tibiae from Evc2/Limbin mutant embryos are shorter than those in control littermates, although body size in these two groups are similar at this stage.
Although EVC was known to occur more frequently in certain families and/or communities (e.g., the Pennsylvania Amish), several decades passed before the causative gene(s) (i.e., EVC and EVC2/LIMBIN) were identified. Mutations in either of these head-to-head genes located on human chromosome 4p were found to contribute to EVC and a dominant but phenotypically milder form called Weyers acrodental dysostosis (also known as Curry–Hall syndrome, WAD; MIM ID #193530)) [3,5]. The primary difference between EVC and WAD is the severity of clinical phenotypes and pattern of inheritance. Though mutations in EVC and EVC2 account for the majority of the patients, there are still patients that present with EVC symptoms despite no mutations in either EVC or EVC2 [6,7]. EVC is an autosomal recessive disorder and is associated with a more severe phenotype, specifically heart abnormalities, which are the main cause of morbidity. Since EVC and EVC2 share limited homology, EVC and EVC2 were named based on a historic timeline of mapping and cloning of causative genes for EVC, and thus they do not form a gene family. In this manuscript, we use both EVC2 and LIMBIN to describe this gene, because pathophysiologic function of this gene was firstly identified as a causative gene of Bovine chondrodysplastic dwarfism and named as LIMBIN [8].
Animal models of EVC have been critical for providing insight into disease pathogenesis (detailed in subsequent sections) [9,10,11]. The initial observation was from EVC2/LIMBIN mutant cattle that developed bovine chondrodysplastic dwarfism (BCD) (Figure 1A, B). Studies of mice revealed phenotypes similar to cattle (Figure 1C, D, E), which is the result of reduced, but not eliminated, hedgehog (Hh) signaling. Both EVC and EVC2/LIMBIN proteins have also been shown to form a complex at the bottom of primary cilia. This has led to the classification of EVC as a ciliopathy or disorder of the primary cilia. Because the management of patients with EVC is currently limited to corrective therapy, deepened understanding of disease pathogenesis from animal models raises the possibility of targeted therapy and improved clinical outcomes.
- EVC-Like Disorders in Other Non-Human Species
Examples of dwarfism or small stature resulting from a medial or genetic condition are not uncommon throughout the animal kingdom. However, while news headlines are dominated by the discovery of insular dwarf species and deliberate selection has led to the development of numerous toy and true “dwarf” animal varieties, chondrodysplastic cases attributable to mutations in animal orthologs (specifically EVC2/LIMBIN) are restricted to two breeds of cattle. However, recent comparative studies have the potential to spark interest in the etiology of EVC-like disorders and reveal the evolutionary significance of EVC2/LIMBIN in other species.
The Japanese brown is a horned cattle breed esteemed for its use in beef production, known as wagyu. Starting in the late 1980s, calves from a specific region of Japan were born with a type of dwarfism (i.e., ateliosis) characterized by disproportionately short limbs and joint abnormalities [57]. Termed bovine chondrodysplastic dwarfism (BCD), this unusual phenotype was inherited in an autosomal recessive fashion and was later found to be caused by either deletion or frameshift mutations in a gene (i.e., LIMBIN) located on chromosome 6 [57,58]. LIMBIN was subsequently identified as an ortholog of human EVC2 despite the lessened severity of symptoms in bovine vs. human patients [8]. Identification and targeted manipulation of the murine ortholog laid the groundwork for the generation and phenotypic characterization of non-human models of EVC (see previous sections) that have been invaluable in our mechanistic understanding of disease development.
A similar form of disproportionate dwarfism in Tyrol grey cattle was reported in the early 21st century. The Tyrol grey is a horned, dual purpose breed of Austrian and Italian origin considered endangered due to its small population (<4000 individuals vs. >20,000 for Japanese brown) [59,60]. Affected calves could be traced back to a single ancestor and displayed the characteristic small stature and short limbs seen in both Japanese BCD and human EVC examples [59]. In addition to growth plate abnormalities in long bones indicative of dysplastic chondrocyte function, some Tyrol BCD specimens also displayed acetabular joint laxity, urogenital abnormalities, and cardiac defects reminiscent of human patients with EVC [59,61]. However, orofacial, ectodermal, and digit findings in these cattle were unremarkable and appendages were shortened in a proximal vs. distal manner as indicated by the shortened femur and humerus. Though genetic analyses identified an EVC2/LIMBIN mutation (i.e., deletion) at a different location from their Japanese counterparts, these cattle provided an animal model of an EVC-like disorder and expanded insight into the role of EVC2/LIMBIN in skeletogenesis.
Penguins are flightless birds that have undergone numerous morphological adaptations suited to an aquatic lifestyle. These include modification of the upper appendages into robust, flipper-like wings and pronounced shortening of femoral and lower extremity length, characteristics that are superficially like those seen in patients with EVC (see previous sections). Genetic analysis of two penguin species (i.e., Pygoscelis adeliae and P. forsteri) surprisingly revealed five penguin-specific amino acid changes in EVC2/LIMBIN, the highest amongst all analyzed limb-related genes [62]. An additional amino acid change was seen in the penguin EVC ortholog. These results suggest important evolutionary roles of EVC and EVC2/LIMBIN orthologs due to their conserved effects on vertebrate (e.g., mammalian and avian) limb development.
References
- Ellis, R.W.B.; Van Creveld, S. A syndrome characterized by ectodermal dysplasia, polydactyly, chondro-dysplasia and congenital morbus cordis: Report of three cases. Arch. Dis. Child. 1940, 15, 65–84.
- McKusick, V.A. Dwarfism in the Amish I. The Ellis-van Creveld Syndrome. Bull. Johns Hopkins Hosp. 1964, 115, 306–336.
- Ruiz-Perez, V.L.; Ide, S.E.; Strom, T.M.; Lorenz, B.; Wilson, D.; Woods, K.; King, L.; Francomano, C.; Freisinger, P.; Spranger, S.; et al. Mutations in a new gene in Ellis-van Creveld syndrome and Weyers acrodental dysostosis. Nat. Genet. 2000, 24, 283–286.
- Baujat, G.; Merrer, M.L. Ellis-Van Creveld syndrome. Orphanet J. Rare Dis. 2007, 2.
- Ruiz-Perez, V.L.; Goodship, J.A. Ellis-van Creveld syndrome and Weyers acrodental dysostosis are caused by cilia-mediated diminished response to Hedgehog ligands. Am. J. Med. Genet. Part C Semin. Med. Genet. 2009, 151, 341–351.
- Tompson, S.W.; Ruiz-Perez, V.L.; Blair, H.J.; Barton, S.; Navarro, V.; Robson, J.L.; Wright, M.J.; Goodship, J.A. Sequencing EVC and EVC2 identifies mutations in two-thirds of Ellis-van Creveld syndrome patients. Hum. Genet. 2007, 120, 663–670.
- D’Asdia, M.C.; Torrente, I.; Consoli, F.; Ferese, R.; Magliozzi, M.; Bernardini, L.; Guida, V.; Digilio, M.C.; Marino, B.; Dallapiccola, B.; et al. Novel and recurrent EVC and EVC2 mutations in Ellis-van Creveld syndrome and Weyers acrofacial dyostosis. Eur. J. Med. Genet. 2013, 56, 80–87.
- Takeda, H.; Takami, M.; Oguni, T.; Tsuji, T.; Yoneda, K.; Sato, H.; Ihara, N.; Itoh, T.; Kata, S.R.; Mishina, Y.; et al. Positional cloning of the gene LIMBIN responsible for bovine chondrodysplastic dwarfism. Proc. Natl. Acad. Sci. USA 2002, 99, 10549–10554.
- Zhang, H.; Takeda, H.; Tsuji, T.; Kamiya, N.; Rajderkar, S.; Louie, K.A.; Collier, C.; Scott, G.; Ray, M.; Mochida, Y.; et al. Generation of Evc2/Limbin global and conditional KO mice and its roles during mineralized tissue formation. Genesis 2015, 53, 612–626.
- Ruiz-Perez, V.L.; Blair, H.J.; Rodrigues-Andres, M.E.; Blanco, M.J.; Wilson, A.; Liu, Y.N.; Miles, C.; Peters, H.; Goodship, J.A. Evc is a positive mediator of Ihh-regulated bone growth that localises at the base of chondrocyte cilia. Development 2007, 134, 2903–2912.
- Caparrós-Martín, J.A.; Valencia, M.; Reytor, E.; Pacheco, M.; Fernandez, M.; Perez-Aytes, A.; Gean, E.; Lapunzina, P.; Peters, H.; Goodship, J.A.; et al. The ciliary EVC/EVC2 complex interacts with smo and controls hedgehog pathway activity in chondrocytes by regulating Sufu/Gli3 dissociation and Gli3 trafficking in primary cilia. Hum. Mol. Genet. 2013, 22, 124–139.
- Morimoto, Y.; Ishibashi, T.; Ashizawa, H.; Shibata, T. Chondrodysplastic Dwarfism in Japanese Brown Cattle. J. Jpn. Vet. Med. Assoc. 1989, 42, 173–177.
- Yoneda, K.; Moritomo, Y.; Takami, M.; Hirata, S.; Kikukawa, Y.; Kunieda, T. Localization of a locus responsible for the bovine chondrodysplastic dwarfism (bcd) on chromosome 6. Mamm. Genome 1999, 10, 597–600.
- Murgiano, L.; Jagannathan, V.; Benazzi, C.; Bolcato, M.; Brunetti, B.; Muscatello, L.V.; Dittmer, K.; Piffer, C.; Gentile, A.; Drögemüller, C. Deletion in the EVC2 gene causes chondrodysplastic dwarfism in Tyrolean grey cattle. PLoS ONE 2014, 9, e0094861.
- Gotoh, T.; Nishimura, T.; Kuchida, K.; Mannen, H. The Japanese Wagyu beef industry: Current situation and future prospects—A review. Asian-Australas. J. Anim. Sci. 2018, 31, 933–950.
- Muscatello, L.V.; Benazzi, C.; Dittmer, K.E.; Thompson, K.G.; Murgiano, L.; Drögemüller, C.; Avallone, G.; Gentile, A.; Edwards, J.F.; Piffer, C.; et al. Ellis–van Creveld Syndrome in Grey Alpine Cattle: Morphologic, Immunophenotypic, and Molecular Characterization. Vet. Pathol. 2015, 52, 957–966.
- Li, C.; Zhang, Y.; Li, J.; Kong, L.; Hu, H.; Pan, H.; Xu, L.; Deng, Y.; Li, Q.; Jin, L.; et al. Two Antarctic penguin genomes reveal insights into their evolutionary history and molecular changes related to the Antarctic environment. GigaScience 2014, 3, 1–15.



