 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Krzysztof Czaja | + 2949 word(s) | 2949 | 2021-01-08 05:25:40 | | | |
| 2 | Bruce Ren | -21 word(s) | 2928 | 2021-01-15 03:47:54 | | |
Video Upload Options
The microbiome lies at the forefront of scientific research, as researchers work to uncover its mys-terious influence on human development and disease.
1. The Microbiome: Not New, Just Newly Defined
The human “microbiome”, as it is often referred to today, is the collection of 10–100 trillion symbiotic microbes that live on and within the human body [1]. This microbiome represents the direct link between all living things on the planet and presents a unique avenue for researchers to understand this living link. As early as 1683, Dutch scientist Antonie van Leeuwenhoek noted in his own oral and fecal samples, “little living animalcules, very prettily a-moving” [2][3][4]. The scientific community, building upon these centuries of research, has now come to understand that the human microbiome plays a profound role in human development and growth, and in adult health. Microbiota can be found on nearly every surface of the human body, and diversely influence critical events in early development, the digestion of food, and the onset of disease [5]. While the progress made since Leewenhoek’s original discovery has been exponential, researchers have only begun to understand the role the human microbiome plays in our daily function.
The Current Stage of Microbiome Research.
2. Animals Models
Rodent models have found popularity in gut microbiome research (Figure 1), having been subjected to a wide range of methods to change their microbiota. A popular field applied to research in rodents is gnotobiology, the study of animals that are completely devoid of microorganisms or those that contain a defined and known set of microbial species. In such experiments, the mice are either germ-free (GF) and thus contain no species at all, monoxenic with just one known colonized species, or have defined microbiota with all species known [6]. There is a clear advantage in using GF mice when compared with a mono-associated (monoxenic) or defined microbiota. However, the microbiome plays a crucial role in development and GF mice exhibit related physiological consequences, including underdeveloped immune systems. A study found that GF 129S6 and GF C57BL/6 mice contained nearly no Th17 cells, and GF Swiss–Webster mice completely lacked any presence of Th17 cells [7]. Transplanting fecal matter from normal Jackson B6 or Taconic B6 mice into C57BL/6 mice via oral gavage resulted in normal Th17 cell levels after 10–12 days. However, these abnormalities in the development of gnotobiotic models, as well as their high cost, have limited their use in research.
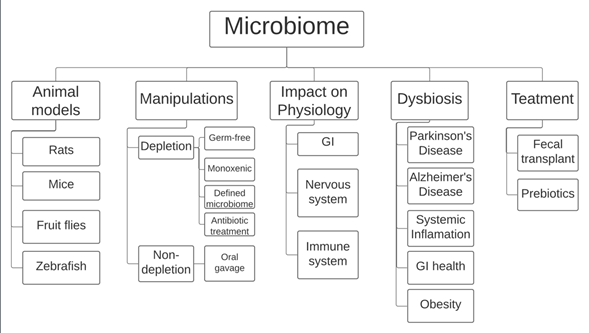
Figure 1. Currently, the field of microbiome research employs rodents (mice and rats), the fly Drosophila melanogaster, and the fish zebrafish as animal models. Most manipulations involve either complete/partial depletion of the microbiome using specific rodent models or microbiome transplants using oral gavage. Research has shown that the microbiome is heavily involved in improper development of the gastrointestinal tract and the nervous and immune systems. A link has also been made between imbalances in the gut microbiome, also known as dysbiosis, and certain diseases, including Parkinson’s and Alzheimer’s disease and obesity, among others. Gut dysbiosis is currently treated in humans using prebiotics and/or fecal transplants. GI: gastro-intestinal tract.
There are two major ways to deplete the gut microbiome in mice to study its relationship with disease and physiology. The first is the previously mentioned GF model. The other model is an antibiotic treatment that can destroy the entire microbiome, often referred to as antibiotic-induced microbiome depletion (AIMD). The antibiotic model helps to avoid several issues that arise in GF models, the most significant advantage being that mice can undergo a normal colonization and full postnatal development following birth [8]. Allowing the rodents to fully develop before depleting the gut microbiome ensures that many of the immune system complications that arise in GF models do not interfere with the study.
The most popular method to manipulate the gut microbiome that does not involve depletion is oral gavage-assisted fecal transplants (Figure 1), in which a large tube is placed through the mouth and into the digestive tract of the rodent to deliver the fecal matter. With the ability to accurately control the dosage and placement of fecal matter and other compounds, the stressful side effects of gavage are often overlooked. Researchers have studied the process of oral gavage on its own to precisely identify how it directly impacts the physiological state in a rodent. A study reported that daily manual restraint and oral gavage in mice did not have a negative impact in body weight change, nylon bone toy weight change, fecal corticosterone levels, and response to the anxiety drug chlordiazepoxide in a conditioned place preference test [9]. However, although several studies have concluded that there is no stressful impact induced by oral gavage on rodents, they still represent the minority. Wide-spanning literature analysis of various laboratory procedures performed on rodents reveals that orogastric gavage, among other practices, has a significant effect on stress-related responses such as increased heart rate, blood pressure, and corticosterone concentrations [10]. These orogastric gavage studies span from rodents given dry and aqueous gavage to those that are administered gavage either while awake or under anesthesia. An objectively less stressful alternative to orogastric gavage to administer a fecal matter transplant (FMT) is the use of pills, which the subject rodents can simply swallow. However, a switch to pill consumption as a method of FMT has not been popular due to the chemical absorption of the pill as it descends through the GI tract.
While rodent models are the most popular model for studying the microbiome currently, researchers continue to develop alternate models to research this area. One organism that has made a recent introduction into research regarding microbiome manipulation is that of the Drosophila melongaster (D. melanogaster), commonly known as the fruit fly (Figure 1). One of the major reasons that the D. melanogaster fly has emerged as a model for manipulation is its ability to create and sustain a thriving environment for microbial species. This practice is known as nutritional mutualism, and researchers can use this ability to harvest bacterial contents that originate directly from the D. melanogaster microbiome [11]. While the ability of the fruit fly to readily provide highly concentrated samples of microbiota to researchers makes them a useful model, the absence of any resident commensal strains makes it difficult to draw larger conclusions on host–microbe interactions independent of nutritional influences [12].
Along with the fruit fly and rodent, the zebrafish (Danio rerio) has emerged as a key model organism in the study of the microbiome due to its substantial physiological and genetic similarities with humans [13]. In particular, zebrafish possess innate and adaptive immune systems similar to our own. Thus, as microbiome research continues to establish a connection between gut health and the nervous system, the zebrafish model has grown in popularity. The ease with which germ-free zebrafish can be produced in addition to their similar nervous system physiology make them a powerful model in the study of the microbiome (Figure 1). However, the zebrafish model possesses several limitations, primarily due to the aquatic nature of the organism. Zebrafish acquire microbiota solely from aquatic habitats, maintain a temperature of 28 °C at all times, and lack mammary glands and lungs. By contrast, humans derive their microbiome primarily from terrestrial origins, maintain a body temperature nearly ten degrees higher, and could not function without organs such as the lungs. These discrepancies between human and zebrafish physiology greatly limit the direct application of zebrafish studies to clinical research [14].
Thus, current methodology for microbiome research is still lacking. Gnotobiotic animals and heavy antibiotic regimens each yield significant problems in the efficacy of the animal model. To use a subject animal that would begin the study with altered brain function, cytokine expression, and physiology would cast doubt on any conclusions as they relate to humans or even a typical rodent model. Similarly, oral gavage has been shown to be inefficient at best and dangerous at worst in current models. Clearly, there remains a need for a new methodology for microbiome research, one that allows for easy, repeated access to the subject animals’ microbiota.
3. The Invisible Manipulator: How the Microbiome Influences Human Physiology
The vast and diverse nature of the microbiome plays many significant roles in human development (Figure 1). Current research shows select microbiota are present in humans as early as at birth. As the newborn is pushed through the vaginal opening, a direct pathway for intergenerational microbial transmission is formed following the rupturing of the chorioamniotic membrane [15]. This exchange provides a foundation for the microbiota to grow over the course of early childhood, with a powerful upsurge in microbiome development at a young age.
Research has also shown breastfeeding to be a natural pathway for intergenerational sharing of microbiota. A longitudinal study based on gut microbiome development in early childhood defined distinct phases of microbiota progression, which are significantly influenced by breastmilk reception. The microbiome profiles determined by 16S rRNA sequencing of infant stool samples revealed that breastfeeding contributed to significant increases of Bifidobacterium species during the first 14 months of life. By an average of 31 months of age, the microbiome reaches a stable state of bacterial diversity [16]. As the field has expanded, research of the early microbiome has revealed evolutionary links between the microbiome and development of human organ systems, especially that of the nervous, circulatory, and gastrointestinal systems.
Composed of the brain and spinal cord, the central nervous system represents the central compartment of our sensory input, cognition, and motor output. A study comparing the behavioral patterns between GF and conventional mice displayed several instances for the necessity of a microbiota in the development of the nervous system. Results from an elevated maze test suggested that a lack of a microbiome negatively effects cognitive development, as GF mice spent significantly more time in the maze than specific pathogen free (SPF) mice. Additionally, the GF mice displayed an altered expression of motor control and anxiety-like behavior, which is involved in second messenger pathways and synaptic long-term potentiation. Components of gut microbiota appear to regulate the proteins PSD-95 and synaptophysin, which are located in the striatum. These proteins are necessary for synaptogenesis, which is the formation of synapses between neurons within the central nervous system. The results of this study suggest a powerful mechanism for microbial influence on proper brain development [17].
Additionally, the microbiome has a significant influence on the function of glial cells within the nervous system. Glial cells are foundational within the central nervous system, as they form the myelin sheath necessary to protect neurons and allow them to project neural impulses efficiently throughout the body. Profiling microglia from the brains of animals raised under GF conditions revealed significant downregulation of genes necessary for cell activation. Additionally, diminished microglia cell development of GF mice was observed in comparison with SPF mice with normal gut microbiota, emphasizing the necessity of a microbiome in nervous system development [18]. The mechanism of glial cell function plays an important role in modulating spatial and temporal responses to stimuli. A study examining the microglial expression between GF and conventionally raised mice showed that GF mice had a severely diminished response to lipopolysaccharide-induced macrophage activation. Under GF conditions, mice are unable to exhibit functions of glial cells, which is detrimental to the development of myelinating cells throughout the central nervous system [19].
Critical in maintaining an organism’s homeostasis after development of the nervous system is the blood–brain barrier, which describes the selectively permeable nature of blood vessel exchange with the central nervous system. Evidence from studies comparing adult GF mice and conventional mice shows that the gut microbiota also plays a role in the modulation of the blood–brain barrier (BBB). GF mice expressed an increase in blood–brain barrier permeability compared to conventional mice, suggesting that the BBB does not strictly regulate the entry of compounds in the absence of a gut microbiome . Qualitative analysis of mouse embryos from both pathogen-free and GF mothers presented evidence for how the maternal gut microbiota influences the blood–brain barrier of offspring. Antibody infrared-labeled immunoglobin G2b (IgG2b) was administered to the mothers during pregnancy, and the presence of the antibody was assessed in their fetal offspring in order to determine the integrity of the prenatal BBB. Results showed that the IgG2b antibody in GF mothers penetrated to offspring, suggesting a lack of tight junctions in the BBB. Additionally, the expression of occludin, a tight junction protein that helps regulate this epithelial barrier, was significantly lower in the fetal offspring of GF mice [20].
The blood–brain barrier makes up just a small fraction of the blood vessels responsible for transport of nutrients throughout the body. The circulatory system comprises the entirety of these networks, including blood, blood vessels, and the heart itself. Previous research of the gut microbiota has revealed distinct microbial influences on angiogenesis, the development of the blood vessels that comprise our circulatory system. A 2002 study examined the post-natal capillary development of several GF mice using cryosections for Syto61-positive nuclei. Adult GF mice from this study were shown to have arrested growth of capillary networks, suggesting the necessity for a gut microbiome in capillary development. Researchers employed a unique approach to this discovery and were able to induce earlier development of capillary networks in GF mice by inoculation of the gut microbe Bacteroides thetaiotaomicron from the cecums of conventionally raised mice. The mechanism linked to this occurrence of angiogenesis is associated with Paneth cells in the intestinal epithelium, which secrete a variety of proteins for immunity and inflammatory control of the mucosa. The GF mice used in this study were determined to be deficient in their overall composition of Paneth cells, so researchers employed an additional comparison of capillary networks using a set of GF mice with intact Paneth cells. Although both sets of mice lacked a microbiome, those lacking Paneth cells had significantly less developed capillary networks, suggesting the importance of Paneth cells during angiogenesis [21].
A nutritional study on GF mice revealed that the gut microbiota is necessary in order to regulate the activity of Paneth cells. A group of C57BL/6 mice received an antibiotic cocktail, after which intestinal contents were transferred to GF mice via oral gavage. The antibiotic treatment appeared to decrease lysozyme proteins in the C57BL/6 mice; however, the effects were much more severe in the GF mice. Without a gut microbiome, the GF mice not only had lowered lysozyme levels but also severe atrophy of the small intestine, as well as reduced interleukin mRNA levels. This suggests that the immune defense activities of the Paneth cells were inhibited in the absence of a gut microbiome [22]. This causal link in the function of the intestinal Paneth cells is a pathway in which the gut microbiota plays a vital role for circulatory system development.
At the very focal point of microbiome research is the gastrointestinal (GI) system, containing the organs necessary for digestion. Gut bacteria are beginning to be understood as a prominent link in how this change in the gut microbiome contributes to further development of the GI system itself. Intestinal stem cells of the epithelium are able to grow and differentiate among tissues of the GI system throughout life. Analysis of the D. melanogaster fly shows evidence of microbiota-mediated cell proliferation [23]. Acetic acid production is an inherent mediator in the early stages of gut microbiome development. Acetobacter pomorum, a common species of gut microbiota within the D. melanogaster, is an acetic acid bacteria with PQQ-ADH activity, which modulates insulin/insulin-like growth factor signaling (IIS). Insulin growth factors such as IIS have a role in GI development through the proliferation and differentiation of intestinal stem cells. Supplementing the simple sugar diet of wild-type, pomorum-monoassociated, and P3G5-monoassociated flies revealed how acetic acid induces IIS activation in P3G5-monoassociated larvae. The activation of IIS was confirmed by examining the membrane targeting of the pleckstrin homology–green fluorescent protein (PH-GFP) [24]. Evidence present in other species link these inherent gut microbes to the development of the GI system. Specialized cells responsible for the growth of the epithelium utilize G-protein-coupled receptors. An additional study on the microbiota-mediated mechanism for stem cell proliferation used mice fed lactic-acid-producing bacteria (LAB), including aforementioned Bifidobacterium. Results derived from in situ hybridization staining showed that the lactic acid produced by these bacteria targeted specific G-protein-coupled receptors through Wnt-3/beta-catenin signaling pathways. The targeting of receptor 81 was found to mediate stem cell proliferation [25].
Continued research on intestinal stem cells through a study that housed both GF and SPF mice at weaning age introduces more ways in which the gut microbiota drives GI tissue development. Variable regrowth of intestinal stem cells following radiation-reduced injury revealed the necessity for a microbiome to promote growth of the epithelium. Results revealed that the erythroid differentiation regulator-1 (Erdr1) gene was only induced in the SPF mice, not GF mice. This link was attributed to the Erdr1 gene. Although evidence does not point to a specific interaction between elements within the microbiome and the Erdr1 gene, the presence of a microbiome in itself is necessary for organoid growth within the intestines [26]. The aforementioned mechanisms define the foundational necessity for a microbiome in GI development. Research on the GF rat as far back as 1984 reveals that the cecum can expand up to five times its size in the absence of a microbiome. Alongside the muscle atrophy that occurs without the bacteria necessary for growth, this junction between the small and large intestine experiences a disturbance in solute-water resorption that causes a severe increase in osmotic pressure. The discovery of this abdominal distension in GF rats presents an example of the physical manifestations that can occur due to abnormal GI system development [27].
References
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810, doi:10.1038/nature06244.
- Leeuwenhoek, A.V. An abstract of a letter from antonie van leeuwenhoek about animals in the scrurf of the teeth. Philos. Trans. R. Soc. Lond. 1683, 14, 568.
- Tomayko, E.; Pillsbury, L.; Pray, L. The Human Microbiome, Diet, and Health: Workshop Summary. In The Human Microbiome, Diet, and Health: Workshop Summary; National Academies Press: Washington, DC, USA, 2013;
- Dobell, C. The discovery of the intestinal protozoa of man. Proc. R. Soc. Med.1920, 13, 1, doi:10.1177/003591572001301601.
- Ursell, L.K.; Metcalf, J.L.; Parfrey, L.W.; Knight, R. Defining the human microbiome. Nutr. Rev. 2012, 70 (Suppl. 1), S38–44, doi:10.1111/j.1753-4887.2012.00493.x.
- Ericsson, A.C.; Franklin, C.L. Manipulating the gut microbiota: Methods and challenges. ILAR J. 2015, 56, 205–217.
- Ivanov, I.I.; de Llanos Frutos, R.; Manel, N.; Yoshinaga, K.; Rifkin, D.B.; Sartor, R.B.; Finlay, B.B.; Littman, D.R. Specific microbiota direct the differentiation of IL-17-producing T-helper cells in the mucosa of the small intestine. Cell Host Microbe 2008, 4, 337–349.
- Kennedy, E.A.; King, K.Y.; Baldridge, M.T. Mouse microbiota models: Comparing germ-free mice and antibiotics treatment as tools for modifying gut bacteria. Front. Physiol. 2018, 9, 1534.
- Turner, P.V.; Vaughn, E.; Sunohara-Neilson, J.; Ovari, J.; Leri, F. Oral gavage in rats: Animal welfare evaluation. J. Am. Assoc. Lab. Anim. Sci. 2012, 51, 25–30.
- Balcombe, J.P.; Barnard, N.D.; Sandusky, C. Laboratory routines cause animal stress. J. Am. Assoc. Lab. Anim. Sci. 2004, 43, 42–51.
- Ma, D.; Leulier, F. The importance of being persistent: The first true resident gut symbiont in Drosophila. PLoS Biol. 2018, 16, e2006945.
- Ludington, W.B.; Ja, W.W. Drosophila as a model for the gut microbiome. PLoS Pathog. 2020, 16, e1008398, doi:10.1371/journal.ppat.1008398.
- Mohanta, L.; Das, B.C.; Patri, M. Microbial communities modulating brain functioning and behaviors in zebrafish: A mechanistic approach. Microb. Pathog. 2020, 145, 104251, doi:10.1016/j.micpath.2020.104251.
- Kamareddine, L.; Najjar, H.; Sohail, M.U.; Abdulkader, H.; Al-Asmakh, M. The Microbiota and Gut-Related Disorders: Insights from Animal Models. Cells 2020, 9, 2401.
- Dominguez-Bello, M.G.; Godoy-Vitorino, F.; Knight, R.; Blaser, M.J. Role of the microbiome in human development. Gut 2019, 68, 1108–1114, doi:10.1136/gutjnl-2018-317503.
- Stewart, C.J.; Ajami, N.J.; O'Brien, J.L.; Hutchinson, D.S.; Smith, D.P.; Wong, M.C.; Ross, M.C.; Lloyd, R.E.; Doddapaneni, H.; Metcalf, G.A.; et al. Temporal development of the gut microbiome in early childhood from the TEDDY study. Nature 2018, 562, 583–588, doi:10.1038/s41586-018-0617-x.
- Diaz Heijtz, R.; Wang, S.; Anuar, F.; Qian, Y.; Bjorkholm, B.; Samuelsson, A.; Hibberd, M.L.; Forssberg, H.; Pettersson, S. Normal gut microbiota modulates brain development and behavior. Proc. Natl. Acad. Sci. USA 2011, 108, 3047–3052, doi:10.1073/pnas.1010529108.
- Erny, D.; Hrabe de Angelis, A.L.; Jaitin, D.; Wieghofer, P.; Staszewski, O.; David, E.; Keren-Shaul, H.; Mahlakoiv, T.; Jakobshagen, K.; Buch, T.; et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat. Neurosci. 2015, 18, 965–977, doi:10.1038/nn.4030.
- Heiss, C.N.; Olofsson, L.E. The role of the gut microbiota in development, function and disorders of the central nervous system and the enteric nervous system. J. Neuroendocr. 2019, 31, e12684, doi:10.1111/jne.12684.
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Toth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158, doi:10.1126/scitranslmed.3009759.
- Stappenbeck, T.S.; Hooper, L.V.; Gordon, J.I. Developmental regulation of intestinal angiogenesis by indigenous microbes via Paneth cells. Proc. Natl. Acad. Sci. USA 2002, 99, 15451–15455, doi:10.1073/pnas.202604299.
- Wang, J.; Tian, F.; Wang, P.; Zheng, H.; Zhang, Y.; Tian, H.; Zhang, L.; Gao, X.; Wang, X. Gut Microbiota as a Modulator of Paneth Cells During Parenteral Nutrition in Mice. JPEN J. Parenter Enter. Nutr. 2018, 42, 1280–1287, doi:10.1002/jpen.1162.
- Tan, Y.; Wei, Z.; Chen, J.; An, J.; Li, M.; Zhou, L.; Men, Y.; Zhao, S. Save your gut save your age: The role of the microbiome in stem cell ageing. J. Cell Mol. Med. 2019, 23, 4866–4875, doi:10.1111/jcmm.14373.
- Shin, S.C.; Kim, S.H.; You, H.; Kim, B.; Kim, A.C.; Lee, K.A.; Yoon, J.H.; Ryu, J.H.; Lee, W.J. Drosophila microbiome modulates host developmental and metabolic homeostasis via insulin signaling. Science 2011, 334, 670–674, doi:10.1126/science.1212782.
- Lee, Y.S.; Kim, T.Y.; Kim, Y.; Lee, S.H.; Kim, S.; Kang, S.W.; Yang, J.Y.; Baek, I.J.; Sung, Y.H.; Park, Y.Y.; et al. Microbiota-Derived Lactate Accelerates Intestinal Stem-Cell-Mediated Epithelial Development. Cell Host Microbe 2018, 24, 833–846.e836, doi:10.1016/j.chom.2018.11.002.
- Abo, H.; Chassaing, B.; Harusato, A.; Quiros, M.; Brazil, J.C.; Ngo, V.L.; Viennois, E.; Merlin, D.; Gewirtz, A.T.; Nusrat, A.; et al. Erythroid differentiation regulator-1 induced by microbiota in early life drives intestinal stem cell proliferation and regeneration. Nat. Commun. 2020, 11, 513, doi:10.1038/s41467-019-14258-z.
- Kohn, D.F.; Clifford, C.B. Chapter 4—Biology and Diseases of Rats. In Laboratory Animal Medicine, 2nd ed.; Fox, J.G.; Anderson, L.C., Loew, F.M., Quimby, F.W., Eds.; Academic Press: Burlington, UK, 2002; pp. 121–165, doi:10.1016/B978-012263951-7/50007-7.

