 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Seong-Joon Park | + 2574 word(s) | 2574 | 2021-01-11 10:30:27 | | | |
| 2 | Karina Chen | + 1 word(s) | 2575 | 2021-01-12 09:07:12 | | |
Video Upload Options
Melatonin, the nocturnally released pineal hormone, has attracted great attention due to its pleiotropic roles. Although its major role is the regulation of the circadian rhythm, this indoleamine engages in multiple mechanisms, including neurogenesis, antioxidation, and inflammatory responses. Apart from its versatility, this molecule has also gained a reputation as being potentially tumor suppressive, espe-cially in hormone-dependent cancers . Melatonin is able to selectively neutralize the estrogenic effects on the breast. The oncostatic roles of melatonin are based on its diverse effects, including its antioxidative properties, apoptosis induction, and an-ticancer immunity . Interestingly, melatonin is able to protect normal cells from the cytotoxicity accompanied by ionizing radiation (IR). This non-targeted effect fol-lowing IR is alleviated via melatonin-mediated regulation of prostaglandins, Toll-like receptors (TLRs), and transcription factors . Additionally, melatonin has been proposed to enhance the therapeutic efficacy of IR, suggesting that this hormone could be a potential radiosensitizer . The radiosensitization of breast cancer cells by mel-atonin is mediated through reduced cell proliferation, the facilitation of cell cycle ar-rest, downregulated DNA repair, and increased p53 mRNA levels .
1. Melatonin Modulates the Estrogenic Effects in Breast Cancer
Since estrogens play a role in breast cancer growth, the estrogen signaling pathway has become a main focus of cancer therapy. In order to neutralize the effects of estrogens, melatonin could be utilized as it regulates the enzymes responsible for local estrogen synthesis [1]. In human mammary tumors, aromatase, sulfatase, and 17β-HSD1, which are in charge of converting androgens into estrogens, hydrolyzing estrone sulfates to estrone, and transforming estrone to the potent 17β-estradiol, respectively, are reportedly overexpressed [2]. In contrast, estrogen sulfotransferase tends to be decreased, presumably leading to the accumulation of 17β-estradiol in mammary tumor tissue [48]. Since estrogen sulfotransferase is an enzyme that inactivates 17β-estradiol, the levels of 17β-estradiol in mammary tumor tissue are increased [3]. Interestingly, melatonin reduces the activity of aromatase, sulfatase, and 17β-HSD1, and increases estrogen sulfotransferase expression, followed by declining estrogenic effects [2][3][4].
2. Melatonin Exerts Anti-Aromatase Roles via Regulating Cyclooxygenase (COX) Gene Activity
The pineal hormone melatonin is known to have oncostatic functions in hormone-dependent mammary tumors [5]. The regulatory functions of melatonin on the estrogen signaling pathway have made this hormone one of the potential tumor suppressive molecules [6]. Melatonin has been reported to act both as a SERM and a SEEM [7]. Moreover, this pineal hormone facilitates adipocyte differentiation and reduces aromatase activity by reducing estrogen production in the cells adjacent to tumor cells [8]. In addition, melatonin indirectly regulates estrogen synthesis by inhibiting aromatase activity. As described above, the activation of aromatase promoters II and I.3 causes increased aromatase expression in cancerous breast tissue [9]. Furthermore, increases in aromatase promoters II and I.3 are associated with increased cAMP levels via cancerous prostaglandin E2 (PGE2)-secreting epithelial cells [8][10]. It has been discovered that melatonin downregulates the gene expression of aromatase promoters II and I.3 [44,63,64]. Melatonin is known to downregulate the activation of the upstream COX2 pathways, including ERK1/2, JNK, p38 MAPK, and NF–NF-κB [11]. Melatonin-mediated downregulation of COX enzymes reduces PGE2, which is a tumor promoter generated by COX2 and is responsible for cell proliferation, death, and angiogenesis [12][13]. Then, diminished PGE2 reduces intracellular cAMP, leading to the decreased activation of promoters I.3 and II, as well as decreased aromatase levels. In turn, estrogen levels are also decreased due to the reduction in aromatase, ultimately followed by the suppression of breast cancer progression (Figure 1) [9][13].
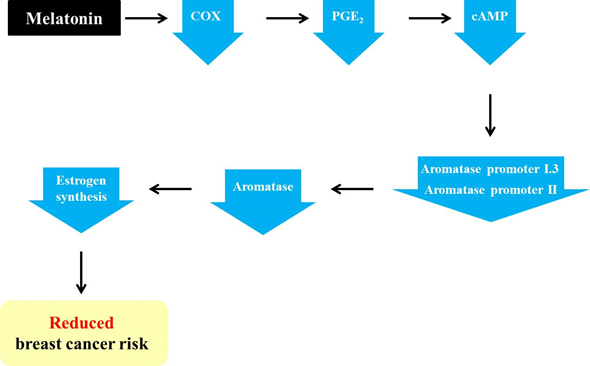
Figure 1. Melatonin-induced reduction in the risk of breast cancer via its regulation of aromatase promoters and related genes. Melatonin downregulates the levels of cyclooxygenase (COX) enzymes and leads to a reduction in prostaglandin E2 (PGE2). In turn, cAMP is also decreased, resulting in a decline in aromatase promoters I.3 and II and, ultimately, aromatase. Then, the decreased aromatase level impedes estrogen synthesis, thereby reducing the risk of breast cancer.
3. Melatonin Increases the Efficiency of Conventional SEEMs and SERMs
As described above, aromatase transforms testosterone and androstenedione into estradiol and estrone [14]. Among these hormones, androstenedione and estrone are low-activity steroids, while testosterone and estradiol possess a high level of activity [15][16]. Low-activity steroids (androstenedione and estrone) are converted into high-activity steroids (testosterone and estradiol) through catalyzation by 17β-hydroxysteroid dehydrogenase type 1, while 17β-hydroxysteroid dehydrogenase type 2 catalyzes the transformation of testosterone and estradiol into androstenedione and estrone, respectively [[15][16]. In breast cancer, the production of these steroids is biased toward the creation of more active steroids [17]. In this way, the production of steroids with high activity is suppressed by melatonin. This hormone inhibits the expression and activity of 17β-hydroxysteroid dehydrogenase type 1, which controls the production of active estrogens from low-activity steroids [18] Melatonin acts as a SEEM by suppressing the expression of enzymes in charge of producing more active steroids from those with a low biological activity, including aromatase, 17β-HSD1, and estrogen sulfatase [18][19]. In other words, melatonin may act as a SEEM, suppressing the biased production of active estrogens. In addition to the role of melatonin as a SEEM, this hormone may also be able to act as a SERM. Melatonin suppresses the binding of estrogen to Erα, meaning that melatonin regulates the proliferation, invasion, protein levels, growth factor expression, and expression of proto-oncogenes, including hTERT, p53, p21, TGFβ, and E-cadherin, of estrogen-dependent breast cancer cells [19]. These SERM actions of melatonin are known to be exerted in an MT1 melatonin receptor-dependent manner, rather than in an ERα-dependent manner. When melatonin binds to the MT1 receptor, this melatonin–MT1 complex reduces ligand–receptor transactivation, interrupting estrogen–ERα binding [20][21][22]. Interestingly, exposure to estrogen in rat ovaries has been shown to cause the downregulation of MT1 melatonin receptors [23]. As well as working as a SERM and SEEM, melatonin also enhances the effects of conventional SEEMs and SERMs. The antiestrogenic effect of tamoxifen has been found to be augmented by the physiological concentration of melatonin, suggesting that this hormone could act as a sensitizing molecule to conventional SEEM [24]. Similarly, the non-steroidal aromatase inhibitor aminoglutethimide shows increased efficiency when cells are pretreated with melatonin, as evidenced by the reduced aromatase mRNA expression in MCF7 cells following pre-exposure to 1 nM of melatonin [25]. Moreover, melatonin seems to be able to alleviate the side effects of aromatase inhibitors. It has been found that the hepatic function disturbances and hepatic toxicity caused by the aromatase inhibitor letrozole are improved by subcutaneously injected melatonin (0.5 mg/kg/day) in female rats [26]. In mammary tumors, adipocytes are found adjacent to tumor cells, and they may engage in cancer development with the potential of triggering cancer cell proliferation, migration, invasion, and resistance [80,81]. It has been documented that irradiation exposure inhibits adipocyte differentiation by downregulating the expression of two major adipogenic regulators, i.e., peroxisome proliferator-activated receptors (PPARγ) and cytidine-cytidine-adenosine-adenosin-thymidine (CCAAT)enhancer-binding proteins α (C/EBPα) [8][27]. Furthermore, radiation has been found to elevate the expression of TNFα, which is known to facilitate breast cancer cell duplication through NF-kB-dependent pathways [28][29]. As described in Section 2.2, cytokines such as TNFα secreted from malignant cells hamper the normal differentiation of mammary adipose cells and increase aromatase activity. Radiation is able to reduce the mRNA levels of aromatase promoter II and COX1/2 expression, leading to decreased aromatase activity [8]. When co-treated with radiation, melatonin counteracts radiation-triggered adipogenic inhibition by increasing PPARγ and C/EBPα and by decreasing TNFα expression [8]. Moreover, this hormone enhances the radiation-induced reduction in aromatase promoter II, COX1, and COX2 expression. In this regard, the hormone melatonin is considered to counteract the side effects, and also to encourage the positive functions provided by radiation by promoting adipocyte differentiation and by repressing aromatase activation (Figure 2).
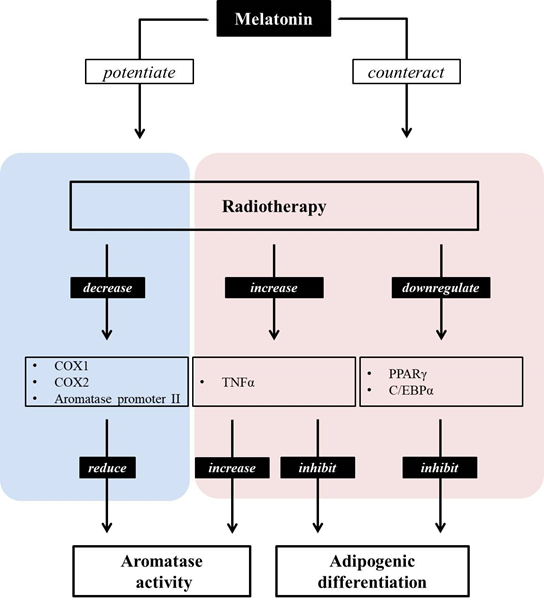
Figure 2. Melatonin can both potentiate and counteract the effects of radiotherapy. Radiation induces a decrease in adipocyte differentiation via downregulating PPARγ and C/EBPα, as well as by increasing TNFα. Furthermore, it reduces aromatase activity by decreasing COX1/2 and aromatase promoter expression. The pineal hormone melatonin potentiates the inhibitory role of aromatase activity, counteracting its suppressive role in the adipocyte differentiation of radiotherapy. Moreover, the increase in aromatase activity following enhanced TNFα levels due to radiotherapy may be counteracted by melatonin.
4. Melatonin Potentiates the Anti-Aromatase Effect of Radiation and Its Presumable Link to p53
Radiotherapy is one of the most widely used therapeutic choices for cancer, though it accompanies numerous unavoidable side effects in spite of recent technological advancements. Therefore, researchers have concentrated on how to reduce the harmful side effects on normal tissues. Several studies have insisted that melatonin is able to enhance the therapeutic effect of radiation and simultaneously protect non-cancerous cells from side effects following radiotherapy [1]. Indeed, melatonin has been proven to potentiate the inhibitory effect of radiation on aromatase expression and activity. The irradiation of 8 Gy combined with pretreatment with a physiological concentration of melatonin (1 nM) leads to a comparatively drastic reduction in aromatase activity and mRNA expression; radiation alone causes 40% and 50% reductions, whereas pretreatment with melatonin before radiotherapy leads to 70% and 75% reductions in aromatase activity and mRNA expression, respectively, leading to the sensitization of breast cancer cells to radiotherapy [30]. The tumor suppressor p53, which is known to play a role in apoptosis, DNA repair, and cell cycle arrest has been reported to be a negative modulator of aromatase in breast cancer [31][32][33]. As melatonin treatment prior to radiotherapy sensitizes breast cancer cells to radiation by downregulating DNA repair and promoting cell cycle arrest, p53 upregulation and consequent aromatase downregulation might represent the link between melatonin and its radiation-sensitizing effect on breast cancer cells.
5. Melatonin Enhances Anti-Angiogenic Function and Suppresses the Disadvantages of Chemotherapeut[34]ic Agents
It has been widely demonstrated that angiogenesis is crucial in cancer treatment, as tumor progression is closely associated with increased angiogenesis. Angiogenesis results from the interplay between breast cancer, endothelial cells, and sex hormones [35]. The extensive formation of new blood vessels represents the vigorous growth of a tumor [36]. The angiogenesis-stimulatory property of estrogens in breast cancer has been well documented. The influence of estradiol on angiogenesis is mediated via its regulation of angiogenic ligands, including vascular endothelial growth factor (VEGF), the soluble form of the VEGF receptor-1 (sVEGF-1), and VEGF receptor-2 (VEGF-2). Concretely, estradiol is able to stimulate VEGF and VEGF-2 and inhibits VEGFR-1, which is a negative regulator of VEGF-mediated angiogenesis, indicating the angiogenesis-favorable characteristics exerted by the hormone [[35]. Melatonin appears to have an inhibitory role in angiogenesis under pathological conditions, including cancer [37]. Its neovascularization-suppressive properties have been suggested to be derived from the downregulation of hypoxia-inducible factor-1α (HIF-1α) and its downstream gene, vascular endothelial growth factor (VEGF) [38]. VEGF has been suggested to be a key factor of angiogenesis in cancer, owing to its neovasculatory function in human breast cancer [39]. Melatonin-induced prevention of HIF-1α nuclear translocation and the subsequent decrease in VEGF expression may hinder the angiogenic gene complex consisting of HIF-1α, phosphor-STAT3, and CBP/p300 in cancer [40].
One chemotherapeutic agent, vinorelbine, disrupts capillary tubule area formation by approximately 75% in human umbilical vein endothelial cells (HUVECs) as compared with non-treated controls, and this effect of vinorelbine is intensified by 1 mM melatonin treatment, suggesting that this pineal molecule could be a potential anti-angiogenic agent [41]. Apart from its benefits, vinorelbine may increase aromatase activity. Intriguingly, melatonin has been found to counteract the stimulatory effect of vinorelbine on aromatase activity by downregulating aromatase promoter I.7, which is one of the major aromatase promoters provoking aromatase transcription in breast cancer [41]. In other words, melatonin supplements the shortcomings, as well as highlights the advantages of chemotherapeutic agents.
Vascular endothelial cells adjacent to tumoral cells are a presumable source of estrogens, as they express aromatase [42]. These endothelial cells express aromatase promoter I.7, which is an endothelial-specific aromatase promoter region in breast cancer [43]. This promoter region may correlate with the angiogenesis of cancerous breast tissue [43]. Excessive expression of aromatase resulting from aromatase promoter I.7 in vascular endothelial tissue adjacent to cancerous breast tissue causes the development of breast tumors via the following two mechanisms: (1) Peripheral estrogen levels may be increased by excessive aromatase activity, leading to direct tumoral growth. (2) An increased concentration of estrogen due to excessive aromatase activity may encourage angiogenesis [44]. Aromatase mRNA is upregulated in endothelial cells via aromatase promoter I.7 [45]. Through the facilitation of the estrogen biosynthesis of endothelial cells, angiogenesis may contribute to the growth of ERα-positive tumors [45], that is, excessive aromatase expression and the consequent enhancement of estrogen synthesis and extensive angiogenesis seem to imply rapid tumoral growth in breasts. Melatonin is reported to downregulate aromatase promoter I.7, the endothelial-specific aromatase promoter region, and also to exert antiangiogenic effects [43]. Through the melatonin-induced downregulation of VEGF expression in mammary tumor cells, VEGF levels in vascular endothelial cells are also reduced [98]. Then, diminished VEGF levels play a suppressive role in the estrogen-producing cells adjacent to tumor cells [31]. In this way, decreased VEGF levels are considered to be vital for angiogenesis reduction in cancer [43].
6. Overexpression of the MT1 Melatonin Receptor has an Aromatase-Suppressive Role and Mediates Oncostatic Action of Melatonin in the MCF7 Human Breast Cancer Cell Line
Several in vivo and in vitro studies have shown the downregulatory role of melatonin in aromatase activity. In MCF7 cells, RT-PCR analysis has revealed that the steady-state level of aromatase mRNA is downregulated by melatonin [46]. Likewise, melatonin-triggered aromatase suppression, a resultant decrease in estrogen synthesis, and reduced tumor growth have been found in mammary tumor rat models [47]. These oncostatic functions of melatonin are known to be exerted through specific receptors [48]. Melatonin exhibits its actions by activating its two high-affinity receptors, MT1 and MT2, throughout the tissues of the body [49][50]. These two melatonin receptors are the most well-characterized melatonin targets, belonging to the G protein-coupled receptor (GPCR) superfamily [50]. As described in Section 3.3, the SERM role of melatonin is associated with the MT1 melatonin receptor, rather than ERα [20][21][22]. Therefore, it can be inferred that the MT1 melatonin receptor is closely related to aromatase activity. The MT1 melatonin receptor is reportedly expressed in the estrogen receptor-positive human breast cancer cell line MCF7, which is sensitive to melatonin-mediated antiproliferation and anti-aromatase effects [22]. This human breast cancer cell line expresses aromatase [51][52], and the MT1 melatonin receptor [53]. Melatonin exerts an inhibitory role on the growth of MCF7 cells. However, it has been demonstrated that this growth-suppressive property of melatonin is enhanced when the MT1 melatonin receptor is overexpressed. Indeed, MT1-transfected MCF7 cells show a significantly enhanced suppression of growth [22]. The activation of the MT1 receptor leads to a reduction in cAMP [54]. Then, cAMP-mediated stimulation of aromatase promoters II and I.3 is increased, since cAMP is involved in the regulation of these aromatase promoters in breast cancer cells [55][56][57]. In this respect, it is speculated that melatonin has the potential to weaken aromatase activity by reducing cAMP levels. The enhanced tumor growth-suppressive role of melatonin appears to be associated with a reduction in aromatase activity following MT1 melatonin receptor overexpression. In MT1-transfected MCF7 cells, a 50% reduction in aromatase activity has been found as compared with vector-transfected cells, indicating that the MT1 melatonin receptor may enhance the inhibitory role of melatonin in relation to aromatase activity and expression [48]. In the same manner, MT1-transfected MCF7 cells in a medium without estrogen show enhanced aromatase suppression as compared with vector-transfected cells in the same medium [43]. Considering that this medium is used to measure aromatase activity, it appears to be more sensitive to melatonin in MT1-overexpressed cells. Plus, implantation with MT1-overexpressed MCF7 cells in mice followed by melatonin administration leads to a 60% reduction in palpable tumors as compared with mice injected with vector-transfected cells [58]. In short, the MT1 melatonin receptor is critical in order for melatonin to exert its oncostatic functions in breast cancer, suggesting that the MT1 melatonin receptor could be a key mediator in breast cancer-related melatonin pathways.
References
- Alonso-Gonzalez, C.; Gonzalez, A.; Menendez-Menendez, J.; Martinez-Campa, C.; Cos, S. Melatonin as a Radio-Sensitizer in Cancer. Boimedicines 2020, 8, 247. [Google Scholar] [CrossRef]
- Cos, S.; Alvarez-García, V.; González, A.; Alonso-González, C.; Martínez-Campa, C. Melatonin modulation of crosstalk among malignant epithelial, endothelial and adipose cells in breast cancer. Oncol. Lett. 2014, 8, 487–492. [Google Scholar] [CrossRef]
- Gonzalez, A.; Cos, S.; Martinez-Campa, C.; Alonso-Gonzalez, C.; Sanchez-Mateos, S.; Mediavilla, M.D.; Sanchez-Barcel, E.J. Selective estrogen enzyme modulator actions of melatonin in human breast cancer cells. J. Pineal Res. 2008, 45, 86–92. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.; Alvarez-Garcia, V.; Martinez-Campa, C.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Sanchez-Barcelo, E.J.; Cos, S. In vivo inhibition of the estrogen sulfatase enzyme and growth of DMBA-induced mammary tumors by melatonin. Curr. Cancer Drug Targets 2010, 10, 279–286. [Google Scholar] [CrossRef]
- Blask, D.E.; Sauer, L.A.; Dauch, R.T. Melatonin as a chronobiotic/anticancer agent: Cellular, biochemical and molecular mechanisms of action and their implications for circadian-based cancer therapy. Curr. Top. Med. Chem. 2002, 2, 113–132. [Google Scholar] [CrossRef]
- Cos, S.; Gonzalez, A.; Martınez-Campa, C.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Sanchez-Barcelom, E.J. Melatonin as a selective estrogen enzyme modulator. Curr. Cancer Drug. Targets 2008, 8, 691–702. [Google Scholar] [CrossRef]
- Menendez-Menendez, J.; Martinez-Campa, C. Melatonin: An anti-tumor agent in hormone-dependent cancers. Int. J. Endocrinol. 2018, 2018, 3271948. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, A.; Nieto, E.G.; Gonzalez, A.; Sanchez-Fernandez, C.; Alonso-Gonzalez, C.; Menendez-Menendez, J.; Gomez-Arozamena, J.; Cos, S.; Martinez-Campa, C. Melatonin Modulation of Radiation and Chemotherapeutics-induced Changes on Differentiation of Breast Fibroblasts. Int. J. Mol. Sci. 2019, 20, 3935. [Google Scholar] [CrossRef]
- Martinez-Campa, C.; Gonzalez, A.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Alvarez-Garcia, V.; Sanchez-Barcel, E.J.; Cos, S. Melatonin inhibits aromatase promoter expression by regulating cyclooxygenases expression and activity in breast cancer cells. Br. J. Cancer. 2009, 101, 1613–1619. [Google Scholar] [CrossRef]
- Brueggemeier, R.W.; Richards, J.A.; Petrel, T.A. Aromatase and cyclooxygenases: Enzymes in breast cancer. J. Steroid Biochem. Mol. Biol. 2003, 86, 501–507. [Google Scholar] [CrossRef]
- Yao, C.; Liu, X.; Zhou, Z.; Xiang, Y.; Yuan, S.; Xie, W.; Zhou, M.; Hu, Z.; Li, Y.; Ji, A.; et al. Melatonin attenuates expression of cyclooxygenase-2 (COX-2) in activated microglia induced by lipopolysaccharide (LPS). J. Toxicol. Environ. Health A 2019, 82, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Dubois, R.N. Prostaglandins and cancer. Gut 2006, 55, 115–122. [Google Scholar] [CrossRef]
- Amin, N.; Shafabakhsh, R.; Reiter, R.J.; Asemi, Z. Melatonin is an appropriate candidate for breast cancer treatment: Based on known molecular mechanisms. J. Cell. Biochem. 2019, 120, 12208–12215. [Google Scholar] [CrossRef]
- Conley, A.; Hinshelwood, M. Mammalian aromatases. Reproduction 2001, 121, 685–695. [Google Scholar] [CrossRef] [PubMed]
- Miettinen, M.M.; Mustonen, M.V.; Poutanen, M.H.; Isomaa, V.V.; Vihko, R.K. Human 17 β-hydroxysteroid dehydrogenase type 1 and type 2 isoenzymes have opposite activities in cultured cells and characteristic cell-and tissue-specific expression. Biochem. J. 1996, 314, 839–845. [Google Scholar] [CrossRef]
- Hilborn, E.; Stål, O.; Jansson, A. Estrogen and androgen-converting enzymes 17β-hydroxysteroid dehydrogenase and their involvement in cancer: With a special focus on 17β-hydroxysteroid dehydrogenase type 1, 2, and breast cancer. Oncotarget 2017, 8, 30552–30562. [Google Scholar] [CrossRef]
- Pasqualini, J.R.; Chetrite, G.S. Recent insight on the control of enzymes involved in estrogen formation and transformation in human breast cancer. J. Steroid Biochem. Mol. Biol. 2005, 93, 221–236. [Google Scholar] [CrossRef]
- González-González, A.; Mediavilla, M.D.; Sánchez-Barceló, E.J. Melatonin: A molecule for reducing breast cancer risk. Molecules 2018, 23, 336. [Google Scholar] [CrossRef]
- Sanchez-Barcelo, E.J.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Rueda, N. Breast cancer therapy based on melatonin. Recent Pat. Endocr. Metab. Immune Drug Discov. 2012, 6, 108–116. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Barceló, E.J.; Cos, S.; Mediavilla, D.; Martinez-Campa, C.; Gonzalez, A.; Alonso-Gonzalez, C. Melatonin–estrogen interactions in breast cancer. J. Pineal Res. 2005, 38, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Treeck, O.; Haldar, C.; Ortmann, O. Antiestrogens modulate MT1 melatonin receptor expression in breast and ovarian cancer cell lines. Oncol. Rep. 2006, 15, 231–235. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Collins, A.R.; Dai, J.; Dubocovich, M.L.; Hill, S.M. MT1 melatonin receptor overexpression enhances the growth suppressive effect of melatonin in human breast cancer cells. Mol. Cell. Endocrinol. 2002, 192, 147–156. [Google Scholar] [CrossRef]
- Clemens, J.W.; Jarzynka, M.J.; Witt-Enderby, P.A. Down-regulation of mt1 melatonin receptors in rat ovary following estrogen exposure. Life Sci. 2001, 69, 27–35. [Google Scholar] [CrossRef]
- Wilson, S.T.; Blask, D.E.; Lemus-Wilson, A.M. Melatonin augments the sensitivity of MCF-7 human breast cancer cells to tamoxifen in vitro. J. Clin. Endocrinol. Metab. 1992, 75, 669–670. [Google Scholar]
- Martínez-Campa, C.; Gonzalez, A.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Sanchez-Barcelo, E.J.; Cos, S. Melatonin enhances the inhibitory effect of aminoglutethimide on aromatase activity in MCF-7 human breast cancer cells. Breast Cancer Res. Treat. 2005, 94, 249–254. [Google Scholar] [CrossRef]
- Aydin, M.; Oktar, S.; Ozkan, O.V.; Alçin, E.; Oztürk, O.H.; Nacar, A. Letrozole induces hepatotoxicity without causing oxidative stress: The protective effect of melatonin. Gynecol. Endocrinol. 2011, 27, 209–215. [Google Scholar] [CrossRef]
- Bindhu, J.; Arunava, D. An edible fungi Pleurotus ostreatus inhibits adipogenesis via suppressing expression of PPAR γ and C/EBP α in 3T3-L1 cells: In vitro validation of gene knock out of RNAs in PPAR γ using CRISPR spcas9. Biomed. Pharmacother. 2019, 116, 109030. [Google Scholar]
- Dong, X.R.; Wang, J.N.; Liu, L.; Chen, X.; Chen, M.-S.; Chen, J.; Ren, J.-H.; Li, Q.; Han, J. Modulation of radiation-induced tumour necrosis factor-α and transforming growth factor β1 expression in the lung tissue by Shengqi Fuzheng injection. Mol. Med. Rep. 2010, 3, 621–627. [Google Scholar]
- Esquivel-Velazquez, M.; Ostoa-Saloma, P.; Palacios-Arreola, M.I.; Nava-Castro, K.E.; Castro, J.I.; Morales-Montor, J. The role of cytokines in breast cancer development and progression. J. Interferon. Cytokine Res. 2015, 35, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Alonso-Gonzalez, C.; Gonzalez, A.; Martinez-Campa, C.; Menendez-Menendez, J.; Gomez-Arozamena, J.; Garcia-Vidal, A.; Cos, S. Melatonin enhancement of the radiosensitivity of human breast cancer cells is associated with the modulation of proteins involved in estrogen biosynthesis. Cancer Lett. 2016, 370, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Docanto, M.M.; Sasano, H.; Lo, C.; Simpson, E.R.; Brown, K.A.; Kathleen Cuningham Foundation Consortium for Research into Familial Breast Cancer. Prostaglandin E2 inhibits p53 in human breast adipose stromal cells: A novel mechanism for the regulation of aromatase in obesity and breast cancer. Cancer Res. 2015, 75, 645–655. [Google Scholar] [CrossRef]
- Speidel, D. The role of DNA damage responses in p53 biology. Arch. Toxikol. 2015, 89, 501–517. [Google Scholar] [CrossRef] [PubMed]
- Berger, C.; Qian, Y.; Chen, X. The p53-estrogen receptor loop in cancer. Curr. Mol. Med. 2013, 13, 1229–1240. [Google Scholar] [CrossRef]
- Endogenous Hormones and Breast Cancer Collaborative Group. Sex hormones and risk of breast cancer in premenopausal women: A collaborative reanalysis of individual participant data from seven prospective studies. Lancet Oncol. 2013, 14, 1009–1019. [Google Scholar] [CrossRef]
- Garvin, S.; Nilsson, U.W.; Dabrosin, C. Effects of oestradiol and tamoxifen on VEGF, soluble VEGFR-1, and VEGFR-2 in breast cancer and endothelial cells. Br. J. Cancer. 2005, 93, 1005–1010. [Google Scholar] [CrossRef]
- Turner, H.E.; Harris, A.L.; Melmed, S.; Wass, J.A.H. Angiogenesis in endocrine tumors. Endocr. Rev. 2003, 24, 600–632. [Google Scholar] [CrossRef]
- Ma, Q.; Reiter, R.J.; Chen, Y. Role of melatonin in controlling angiogenesis under physiological and pathological conditions. Angiogenesis 2020, 23, 91–104. [Google Scholar] [CrossRef]
- Cheng, J.; Yang, H.-L.; Gu, C.-J.; Liu, Y.-K.; Shao, J.; Zhu, R.; He, Y.-Y.; Zhu, X.-Y.; Li, M.-Q. Melatonin restricts the viability and angiogenesis of vascular endothelial cells by suppressing HIF-1α/ROS/VEGF. Int. J. Mol. Med. 2019, 43, 945–955. [Google Scholar] [CrossRef] [PubMed]
- Folkman, J. Angiogenesis in cancer, vascular, rheumatoid and other disease. Nat. Med. 1995, 1, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Goradel, N.H.; Asghari, M.H.; Moloudizargari, M.; Negahdari, B.; Haghi-Aminjan, H.; Abdollahi, M. Melatonin as an angiogenesis inhibitor to combat cancer: Mechanistic evidence. Toxicol. Appl. Pharmacol. 2017, 335, 56–63. [Google Scholar] [CrossRef]
- Gonzalez-Gonzalez, A.; Gonzalez, A.; Rueda, N.; Alonso-Gonzalez, C.; Menendez-Menendez, J.; Martinez-Campa, C.; Mitola, S.; Cos, S. Usefulness of melatonin as complementary to chemotherapeutic agents at different stages of the angiogenic process. Sci. Rep. 2020, 10, 4790. [Google Scholar] [CrossRef] [PubMed]
- Harada, N.; Sasano, H.; Murakami, H.; Ohkuma, T.; Nagura, H.; Takagi, Y. Localized expression of aromatase in human vascular tissues. Circ. Res. 1999, 84, 1285–1291. [Google Scholar] [CrossRef] [PubMed]
- Cos, S.; Martinez-Campa, C.; Gonzalez, A.V.; Alvarez-Garcia, V.; Alonso-Gonzalez, C.; Mediavilla, M.D.; Sanchez-Barcelo, E.J. Melatonin and aromatase in breast cancer. Clin. Cancer Drugs 2014, 1, 54–64. [Google Scholar] [CrossRef]
- Alvarez-García, V.; Gonzalez, A.; Martinez-Campa, C.; Alonso-Gonzalez, C.; Cos, S. Melatonin modulates aromatase activity and expression in endothelial cells. Oncol. Rep. 2013, 29, 2058–2064. [Google Scholar] [CrossRef] [PubMed]
- Irahara, N.; Miyoshi, Y.; Taguchi, T.; Tamaki, Y.; Noguchi, S. Quantitative analysis of aromatase mRNA expression derived from various promoters (I. 4, I. 3, PII and I. 7) and its association with expression of TNF-α, IL-6 and COX-2 mRNAs in human breast cancer. Int. J. Cancer 2006, 118, 1915–1921. [Google Scholar] [CrossRef] [PubMed]
- Cos, S.; Martinez-Campa, C.; Mediavilla, M.D.; Sanchez-Barcelo, E.J. Melatonin modulates aromatase activity in MCF-7 human breast cancer cells. J. Pineal Res. 2005, 38, 136–142. [Google Scholar] [CrossRef]
- Cos, S.; Gonzalez, A.; Guezmes, A.; Mediavilla, M.D.; Martinez-Campa, C.; Alonso-Gonzalez, C.; Sanchez-Barcelo, E.J. Melatonin inhibits the growth of DMBA-induced mammary tumors by decreasing the local biosynthesis of estrogens through the modulation of aromatase activity. Int. J. Cancer 2006, 118, 274–278. [Google Scholar] [CrossRef]
- González, A.; Martinez-Campa, C.; Mediavilla, M.D.; Alonso-Gonzalez, C.; Sanchez-Mateos, S.; Hill, S.M.; Sanchez-Barcelo, E.J.; Cos, S. Effects of MT1 melatonin receptor overexpression on the aromatase-suppressive effect of melatonin in MCF-7 human breast cancer cells. Oncol. Rep. 2007, 17, 947–953. [Google Scholar] [CrossRef]
- Jockers, R.; Delagrange, P.; Dubocovich, M.L.; Markus, R.P.; Renault, N.; Tosini, G.; Cecon, E.; Zlotos, D.P. Update on melatonin receptors: IUPHAR Review 20. Br. J. Pharmacol. 2016, 173, 2702–2725. [Google Scholar] [CrossRef]
- Stauch, B.; Johansson, L.C.; Cherezov, V. Structural insights into melatonin receptors. FEBS J. 2020, 287, 1496–1510. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Wang, J.; Chen, E.; Murai, J.; Siiteri, P.K.; Chen, S. Aromatase gene is amplified in MCF-7 human breast cancer cells. J. Steroid Biochem. Mol. Biol. 1993, 46, 147–153. [Google Scholar] [CrossRef]
- Sonne-Hansen, K.; Lykkesfeldt, A.E. Endogenous aromatization of testosterone results in growth stimulation of the human MCF-7 breast cancer cell line. J. Steroid Biochem. Mol. Biol. 2005, 93, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Ram, P.T.; Kiefer, T.; Silverman, M.; Song, Y.; Brown, G.M.; Hill, S.M. Estrogen receptor transactivation in MCF-7 breast cancer cells by melatonin and growth factors. Mol. Cell. Endocrinol. 1998, 141, 53–64. [Google Scholar] [CrossRef]
- Kiefer, T.; Ram, P.T.; Yuan, L.; Hill, S.M. Melatonin inhibits estrogen receptor transactivation and cAMP levels in breast cancer cells. Breast Cancer Res. Treat. 2002, 71, 37–45. [Google Scholar] [CrossRef]
- Zhao, Y.; Agarwal, V.R.; Mendelson, C.R.; Simpson, E.R. Estrogen biosynthesis proximal to a breast tumor is stimulated by PGE2 via cyclic AMP, leading to activation of promoter II of the CYP19 (aromatase) gene. Endocrinology 1996, 137, 5739–5742. [Google Scholar] [CrossRef]
- Zhou, D.; Clarke, P.; Wang, J.; Chen, S. Identification of a promoter that controls aromatase expression in human breast cancer and adipose stromal cells. J. Biol. Chem. 1996, 271, 15194–15202. [Google Scholar] [CrossRef]
- Bulun, S.E.; Sebastian, S.; Takayama, K.; Suzuki, T.; Sasano, H.; Shozu, M. The human CYP19 (aromatase P450) gene: Update on physiologic roles and genomic organization of promoters. J. Steroid Biochem. Mol. Biol. 2003, 86, 219–224. [Google Scholar] [CrossRef]
- Collins, A.; Yuan, L.; Kiefer, T.L.; Cheng, Q.; Lai, L.; Hill, S.M. Overexpression of the MT1 melatonin receptor in MCF-7 human breast cancer cells inhibits mammary tumor formation in nude mice. Cancer Lett. 2003, 189, 49–57. [Google Scholar] [CrossRef]
- Endogenous Hormones and Breast Cancer Collaborative Group. Sex hormones and risk of breast cancer in premenopausal women: A collaborative reanalysis of individual participant data from seven prospective studies. Lancet Oncol. 2013, 14, 1009–1019. [Google Scholar] [CrossRef]


