Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Manish Tripathi | -- | 4539 | 2024-03-12 23:18:47 | | | |
| 2 | Camila Xu | Meta information modification | 4539 | 2024-03-13 02:38:58 | | | | |
| 3 | Camila Xu | + 2 word(s) | 4541 | 2024-03-18 08:51:34 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Anilkumar, A.K.; Vij, P.; Lopez, S.; Leslie, S.M.; Doxtater, K.; Khan, M.M.; Yallapu, M.M.; Chauhan, S.C.; Maestre, G.E.; Tripathi, M.K. Long Non-Coding RNAs in Neurodegenerative Diseases. Encyclopedia. Available online: https://encyclopedia.pub/entry/56180 (accessed on 28 July 2026).
Anilkumar AK, Vij P, Lopez S, Leslie SM, Doxtater K, Khan MM, et al. Long Non-Coding RNAs in Neurodegenerative Diseases. Encyclopedia. Available at: https://encyclopedia.pub/entry/56180. Accessed July 28, 2026.
Anilkumar, Adithya K., Puneet Vij, Samantha Lopez, Sophia M. Leslie, Kyle Doxtater, Mohammad Moshahid Khan, Murali M. Yallapu, Subhash C. Chauhan, Gladys E. Maestre, Manish K. Tripathi. "Long Non-Coding RNAs in Neurodegenerative Diseases" Encyclopedia, https://encyclopedia.pub/entry/56180 (accessed July 28, 2026).
Anilkumar, A.K., Vij, P., Lopez, S., Leslie, S.M., Doxtater, K., Khan, M.M., Yallapu, M.M., Chauhan, S.C., Maestre, G.E., & Tripathi, M.K. (2024, March 12). Long Non-Coding RNAs in Neurodegenerative Diseases. In Encyclopedia. https://encyclopedia.pub/entry/56180
Anilkumar, Adithya K., et al. "Long Non-Coding RNAs in Neurodegenerative Diseases." Encyclopedia. Web. 12 March, 2024.
Copy Citation
Long non-coding RNAs (lncRNAs) are novel genetic biomarkers that can be used as exclusionary tools specific to Neurodegenerative diseases (NDDs). These historical biomarkers have been there for years, so a change in the approach is necessary to better diagnose and treat these NDDs.
long non-coding (Lnc) RNAs
neurodegenerative diseases (NDDs)
Alzheimer’s disease (AD)
1. Long Non-Coding RNAs: An Overview
Long non-coding RNAs, classified as non-coding RNA composed of more than 200 nucleotides, have been highlighted as important biomolecules in the cellular mechanisms responsible for normal development to disease progression. LncRNAs, being just one of the types of non-coding RNA, can be further divided into multiple categories by their genomic position. These categories include intergenic lncRNAs, antisense lncRNAs, intronic lncRNAs, and bidirectional lncRNAs [1]. Along with varying genomic position, the localization of lncRNAs can also differ with studies showing nuclear and cytoplasmic localization [2][3]. The primary mechanism that drives nuclear localization is unknown, but it can potentially be attributed to the recruitment of nuclear retention factors facilitated by sequence motifs [4][5]. When these sequences were removed, lncRNA exportation was favorable, and cytoplasmic localization was possible. Compared to mRNA, the structure and biogenesis of lncRNA are similar due to the addition of poly-A tails, 5′ 7-methylguanoasine capping, and transcription by RNA Polymerase II [6]. Unlike mRNA, however, lncRNAs are relatively less abundant, lack the open reading frame typically found on mRNA, and, most importantly, lack a protein-coding ability [6]. Previously, this last feature of lncRNA, along with other non-coding RNA, led to their classification as “junk” DNA with no discernible function. This has been refuted by recent research showing the roles that lncRNAs plays in various biological processes, such as the progression of cancer, neurodegenerative diseases, and normal development [7][8][9]. It has been shown that lncRNAs play a crucial role in the regulation of diseases course and normal cellular processes through gene regulation. Figure 1 shows the schematic representation of lncRNA function.
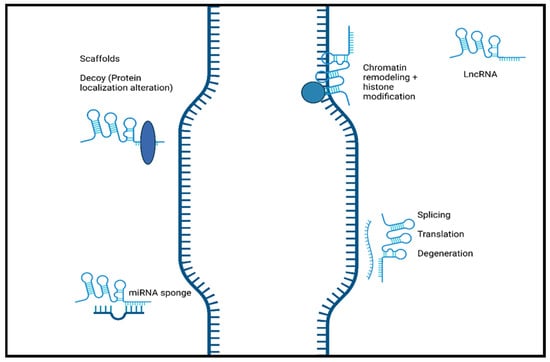
Figure 1. The schematic representation of the function of long non-coding RNA. LncRNAs induce chromatin remodeling and histone modification. Interaction with mRNA. LncRNA hybridization may lead to alternatively spliced transcripts, translation, and mRNA degeneration. LncRNAs interact with proteins/biological molecules to modulate their activity by binding to specific proteins and altering protein localization. They also serve as scaffolds to allow the formation of miRNA sponges. Created with BioRender.com.
Additionally, lncRNA localization correlates highly to function, with nuclear lncRNA functioning primarily as chromatin regulators, while cytoplasmic lncRNAs are more focused on post-translational regulation and the stability of mRNA [5]. LncRNAs have the potential to modulate normal and abnormal development through their mechanisms as decoys, scaffolds, and guides, in addition to direct interaction with DNA and protein molecules [10].
2. LncRNA Mechanisms
LncRNAs play a vital role in disease progression and development that is highly dependent on their well-known functions, including altering gene expression by modulating chromatin structure and regulating transcription and post-transcriptional modifications [11]. These processes can be achieved by lncRNAs acting as guides, decoys, and scaffolds [10][12]. LncRNAs can function as guides and recruit chromatin-remodeling complexes to a specific locus in cis or trans sites [13]. Through recruitment, lncRNAs can target specific sections to alter the structure of chromatin and, therefore, alter the gene expression of that specific gene. LncRNAs can also directly interact with DNA to form hybrid structures such as R-loops or triple helices (triplexes). Triple helices have the potential to mediate gene activation or repression by recruiting remodeling complexes [14]. R-loops can be recognized by a variety of proteins to also promote or suppress gene expression [15]. Furthermore, lncRNAs can interact with proteins and function as scaffolds. As a scaffold, they can aid in the binding of proteins to assemble a complex commonly known as ribonucleoprotein complexes [16]. These complexes have roles in mRNA splicing, translation, and stability, allowing lncRNAs to affect these processes indirectly [2]. LncRNAs can also function as decoys and lower the availability of regulation factors, which can negatively impact gene expression [17]. Furthermore, lncRNAs can regulate pathways and expression through miRNA sponging. This mechanism, enables lncRNAs to act as an miRNA target and bind to miRNA. The result of miRNA sponging is a reduction in the miRNA function, leading to the alteration of signaling pathways that can affect further gene expression [18].
3. LncRNAs in Neurodegenerative Diseases
3.1. Alzheimer’s Disease
Alzheimer’s disease (AD) is a progressive and irreversible neurodegenerative disorder that is the most common form of dementia. This disease is named after Dr. Alois Alzheimer, who noticed that there had been significant changes in the brain of a woman who had unusual symptoms such as memory loss, language problems, and unpredictable behavior. These significant changes are now known as two hallmark characteristics of AD. The first is amyloid plaques, which are produced from amyloid β (Aβ) aggregation, and the second is neurofibrillary tangles (NFT), which are produced from an accumulation of pathological tau [19]. Late-onset AD (LOAD) usually presents as an age-related disease in individuals 65 and over. There is a small portion of individuals with AD who have early-onset Alzheimer’s disease (EOAD), which occurs in people younger than 65 with a genetic predisposition. In the hundred or so years since the discovery of the disease, there have been many studies looking to elucidate the mechanisms of the disease. Although treatments may help relieve some of the symptoms and/or progression associated with NDDs, there are currently no known cures. Therefore, there is a critical need to expand the understanding of what causes neurodegeneration and to develop new approaches for the prevention and treatment of AD [19].
In recent studies, lncRNAs have been shown to be involved in regulatory mechanisms such as transcriptional, posttranscriptional, and translational regulation, as well as a variety of biological functions such as development, differentiation, and metabolism [20]. LncRNAs have also been shown to be involved in neurological diseases such as epilepsy, neurodegenerative diseases, and genetic disorders [21][22][23]. LncRNAs have also been shown to play a role in the pathogenesis of AD, yet the exact mechanism is still unknown. In a microarray analysis study of lncRNAs expressed in the hippocampal region of AD, 315 lncRNAs were found to be dysregulated [24]. Many protein-coding mRNAs have antisense transcript “partners,” which are commonly noncoding RNAs. An example would be the antiscript BACE1-AS, which has been shown to regulate BACE1 mRNA and its protein expression. BACE1 (beta-site amyloid precursor protein cleaving enzyme1) is required for the formation of all monomeric Aβ (1-42) and is also thought to be the cause of toxicity in AD patients [25]. This study shows that lncRNAs are responsible for the increase in Aβ 1-42 in AD, showing that lncRNAs influence the pathogenesis of AD [26].
3.1.1. Competitive Endogenous RNA (ceRNA) Theory
Several studies have reported that lncRNA competes with miRNA target genes by sharing common binding sites [22]. Based on the ceRNA theory, Wang and collaborators created a global triple network where lncRNAs and mRNAs form a triplicate that shares the same miRNA. Based on this network, an AD NFT-associated lncRNA-mRNA network (NFTLMN) was created, mapped, and analyzed, providing three lncRNAs highly related to AD NFT [22]. Gene ontology (GO) function and KEGG pathway enrichment analyses were performed on AP000265.1, KB-1460A1.5, and RP11-145M9.4, showing GO terms for formation and development of the neural tube, neural crest cells, and epithelial tube morphogenesis. Phosphorylation terms were also found during the analysis [22]. A different study found 40 pairs of lncRNAs that shared more than one disordered miRNA, 9 of them correlated with other neurodegenerative disorders, and 5 lncRNAs that could be potential biomarkers for [27]. These machine learning studies have found several lncRNAs that would be suitable for further research as possible biomarkers since they have been identified as showing a correlation with AD genes, but their exact function is unknown.
3.1.2. LncRNA Involvement
Many lncRNAs have been identified as having a role in AD. Although many of these newly identified lncRNAs have been found through bioinformatics, many of these lncRNA functions remain obscure. The lncRNAs related to AD that have been studied have been discovered to be involved in synaptic and neuron exhaustion, neurotrophin depletion, inflammation, mitochondrial impairment, oxidative stress, and DNA damage [28]. lncRNAs, such as NDM29, BC200, 51A, and BACE1-AS, are differentially expressed in AD and correlated with AD progression [29]. Neuroblastoma differentiation marker 29 (NDM29), a lncRNA that promotes the cleavage of BACE and γ-secretase, plays a critical role in AD pathogenesis by inducing an inflammatory response. Studies show NDM29 can induce APP synthesis, increasing Aβ and Aβ-42/Aβ-40 ratio [28][29]. The BACE1 antisense transcript (BACE1-AS) regulates BACE1 mRNA and protein expression when exposed to Aβ-42 [28]. BACE1-AS is significantly upregulated in the cerebellum, hippocampus, entorhinal cortex, and superior frontal gyrus of the AD brain. There is a synergistic mechanism in how BACE1-AS regulates BACE1, which can promote target mRNA or inhibit miRNA [30]. MALAT1 is a highly abundant and evolutionarily conserved lncRNA and regulates a subset of genes involved in synaptic plasticity. Recent studies have reported reduced lncRNA MALAT1 levels in the central spine fluid and brains AD patients compared with a control group [31][32]. Several preclinical studies have provided evidence and support for the potential roles of lncRNA MALAT1 in AD pathogenesis. For instance, studies in experimental AD models indicated enhanced neurite outgrowth, reduced proinflammatory cytokines, decreased neuronal apoptosis, and increased presynaptic bouton density on dendrites with lncRNA MALAT1 overexpression, and vice versa with lncRNA MALAT1 knockdown [33]. Similarly, a recent study suggested the beneficial effect of lncRNA MALAT1 against Aβ1-42-induced toxicity [31]. lncRNA-51A overlaps with SORL1 antisense and could affect amyloid beta formation, which is known to be upregulated in AD [22]. Sortilin-related receptor 1 (SORL1) is one of the genes involved in the processing of amyloid-β protein precursor (APP) and is thought to be a genetic factor of AD. LncRNA-51A downregulates SORL1, which results in abnormal APP processing [34]. BC200 exhibits abnormal subcellular localization and expression levels in specific brain regions in AD patients [21]. BC200 is a protomer-associated RNA that works on the translation level by increasing synapse loss [28]. The loss of synapses is one of the pathological features of AD and is the cause of memory loss in AD patients. In normal aging, BC200 was found to be downregulated, but in AD brains it was found to be significantly upregulated in brain areas related to AD. Furthermore, relative levels of BC200 RNA in AD-affected areas of the brain increased according to the severity of the disease [35]. Li et al., found BC200 could be a positive regulator of BACE1 in AD [36]. Many studies have shown that lncRNA nuclear paraspeckle assembly transport 1 (NEAT1) promotes inflammation and has a role in neurodegenerative disorders. NEAT1 is upregulated in AD and progresses via the miR-124–BACE1 axis showing it can be manipulated to modulate BACE1 expression. This information demonstrates that NEAT1 can be a target for pharmacological therapies and a biomarker for the disease [37]. With the continuing bioinformatic research, several lncRNAs have been identified with tentative functions and correlations. A study by Nana Ma and collaborators identified 487 significantly dysregulated lncRNAs in AD model mice (APP/PS1) brains. These lncRNAs were found to be involved in synaptic plasticity and memory (Akap5), and regulation of amyloid-β induced neuroinflammation (Klf4) [38]. Transcriptomic analysis identified RP3-522J7, MIR3180-2, and MIR3180-3 as lncRNAs which were most highly co-expressed with known AD-related genes [39]. Shi et al., also investigated genomic localization and found that several lncRNAs are located near important protein-coding genes (PCGs) in the human genome. For example, six differentially expressed lncRNAs were within 10 MB of the PCG S100B [39]. These non-coding RNAs have been identified by machine learning and, as such, have no identified function at this time. Figure 2 shows the lncRNAs involved in Alzheimer’s disease.
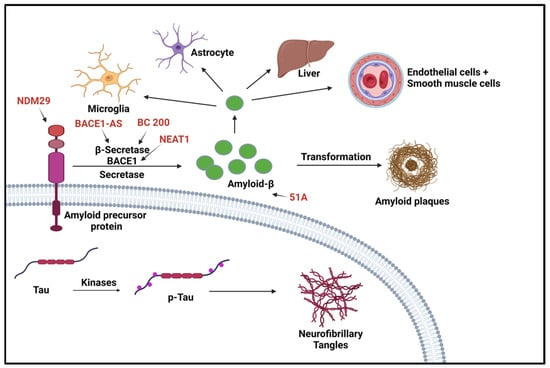
Figure 2. LncRNAs involved in Alzheimer’s disease. Two aspects of this disease are shown here. The first aspect is the production/aggregation of Amyloid-β (Aβ) peptides, which are the final product of amyloid precursor proteins (APPs); the other aspect is the accumulation of neurofibrillary tangles (NFTs) caused by hyperphosphorylated microtubule-associated protein (p-Tau). BACE1 is the rate-limiting enzyme for amyloid precursor proteolysis and is affected by BC200 and BAEC1-AS. NDM29 induces the formation of APP, which in turn promotes an increase in Aβ formation [40]. “Created with BioRender.com”.
3.2. Parkinson’s Disease
Parkinson’s disease (PD) is a progressive neurodegenerative disease that is characterized by death or malfunction of dopaminergic neurons in the substantia nigra and dopamine depletion in the striatum resulting in the loss of motor and non-motor functions. Approximately 60,000 new PD cases are diagnosed each year, joining the 1 million Americans who currently have PD. The direct and indirect annual medical costs for PD approach $25.4 billion and $26.5 billion, respectively. Despite having gained considerable knowledge about the pathological mechanisms of PD over the last several decades, there is little know about how to stop or delay the ongoing neurodegenerative processes. Symptoms typically occur when approximately 70% of the neurons are lost, and other characterizations, such as Lewy bodies composed of α-synuclein, can accumulate in the substantia nigra [41]. Currently, there is no effective cure for PD, and most medications are prescribed for symptom management. Methods of increasing striatal dopamine, such as levodopa, dopamine antagonists, and monoamine oxidase B inhibitors, are used to treat the motor implications of Parkinson’s disease [42]. Although these medications help alleviate motor symptoms, they do not aid in slowing disease progression. Thus, PD patients are in urgent need of disease-modifying therapies that can slow or stop the progression of the neurodegenerative process. The exact molecular mechanisms of PD development are not known. However, lncRNAs have been highly regarded as potential regulators for Parkinson’s disease. This potential is due to their role in biological processes and studies showing altered pathways in PD. Specific lncRNAs have been identified as altering expressions in PD patients compared to controls in samples including brain tissue, blood, and cerebrospinal fluid. Some examples of these lncRNAs include AL049437 and AK021630, which were found to be significantly upregulated and downregulated, respectively, in PD [43]. Mechanistically, lncRNAs found to be involved in PD pathogenicity aid or worsen the disease through various pathways, with most studies targeting primary characteristics in the pathophysiology of PD such as neuronal injury, inflammation, and α-synuclein accumulation.
LncRNA Involvement
Neuronal damage is prevalent in Parkinson’s disease, with most symptoms resulting from the loss of dopaminergic neurons. Some lncRNAs are involved in regulating injury through autophagy and the apoptosis of neuronal cells. LncRNA NEAT1 has been found to be upregulated in mice with MPTP-induced PD, which promotes the stability of PINK1 expression. As a result, it promotes the autophagy caused by MPTP to induce PD [44]. Alternatively, NEAT1 can regulate PD progression in MPP+ SK-N-SH cells by functioning as an miRNA sponge for miR-212-3p and, therefore, modulating the expression of AXIN1 protein to mediate cellular apoptosis [45]. Similarly, lncRNA BDNF-AS, when upregulated, works by downregulating miR-125b-5p to regulate autophagy and apoptosis in cells treated with MPP+ [46]. H19 is also involved in autophagy regulation, where MPP+-induced apoptosis was reduced in H19 overexpressing cells through the negative regulation of miR-585-3p [47]. Additionally, H19 overexpression in induced PD can inhibit the function of miR-301b-3p. When inhibited, it increases the expression of HRPT, usually deficient in PD, through the Wnt/B-catenin pathway, decreasing the loss of dopaminergic neurons [48]. LncRNA Xist is also involved in increasing neuronal injury through the repression of miR-199a-3p. This act allows Xist to induce the expression of transcription factor Sp1, which promotes LRRK2 and has been shown to advance PD progression [49]. Furthermore, lncRNA HOTAIR, which is found to be upregulated, furthered PD through an increase in ROS generation and neuroinflammation; therefore, inducing neuronal injury. HOTAIR potentially increases neuronal injury by regulating the autophagy protein ATG10. The protein expression is promoted through HOTAIR, acting as a sponge for miR-874-5p [50]. Other lncRNA studies have focused on the role lncRNAs play in significant inflammation in PD cases, often regulated by microglial cells, and how this can lead to the progression of neuron injury [51][52]. Patients with PD tend to have a consistent inflammatory response, which could worsen neuron injury [53]. LncRNA MALAT1 was upregulated in MPTP-treated mice, along with increased expression of pro-inflammatory cytokines through epigenetic regulation. In BV2 cells, MALAT1 epigenetically regulated Nrf2 by binding to EZH2. When the expression of Nrf2 is reduced, it increases ROS and inflammation leading to the injury of neurons [54]. LncRNA TUG1 was upregulated in PD-induced mice along with pro-inflammatory cytokines IL-6 and TNF-α [55]. LncRNA UCA1 influences oxidative stress and the expression of TNF-α, IL-6, and IL-1β. In the 6-OHDA PD mouse model, a downregulated UCA1 reduced activation of the PI3K/AKT pathway. The PI3K/AKT pathway is usually involved in a variety of cellular pathways, including neurodegenerative diseases such as PD [56]. Dysregulation of α-synuclein, commonly found in PD, has been correlated to impairment in a variety of cellular processes, such as synaptic vesicles, mitochondria function, and the autophagy-lysosomal pathway, leading to a problem in dopamine levels [57]. These factors make it a therapeutic target for studies determining the involvement of lncRNA regulation and improving PD pathogenesis. LncRNA SNHG1 overexpression reduced miR-15-5p expression and promoted α-synuclein accumulation through the SIAH1 protein. Furthermore, overexpression of lncRNA OIP5-AS1 reduced the α-synuclein accumulation by miR-126 binding and PLK2/α-synuclein autophagy [58]. In a rotenone-induced PD mouse model, lncRNA SNHG14 was discovered to be upregulated with a reversed expression of miR-133b. Upregulation of SNHG14 correlated with increased neuronal injury and an increased a-syn expression through the downregulation of miR-133b [59]. In addition, lncRNA-T199678 expression, through binding and regulation of miR-101-3p, can regulate ROS and apoptosis that was induced by α-syn [60]. Figure 3 shows the lncRNAs involved in Parkinson’s disease.
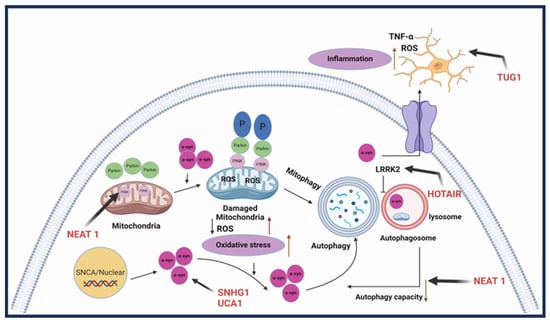
Figure 3. LncRNAs involved in Parkinson’s disease. Parkinson’s disease pathogenesis involves three aspects: the first is dopaminergic neuron death; the second is an aggregation of synuclein-alpha, which forms Lewy bodies; and the final one is neuroinflammation, causing cell death. LncRNA HOTAIR, SNHG1, and UCHL1-AS participate in the accumulation of SNCA. LncRNA NEAT1 causes abnormal autophagy. HOTAIR can also upregulate LRRK2 and thus affect autophagy. LncRNA TUG1 regulates microglia polarization and increases inflammatory cytokine production [40]. Created with BioRender.com.
3.3. Amyotrophic Lateral Sclerosis
Amyotrophic lateral sclerosis, also known as Lou Gehrig’s disease, is a progressive neuromuscular degeneration that primarily affects motor neurons of the somatic nervous system. The gradual erosion of these neuromuscular connections, often starting in distal muscles, encompasses loss in much of the motor functions necessary for daily tasks [61]. Deterioration of the efferent pathways from the brain and spinal cord to effectors incites voluntary muscular atrophy. Though some variation exists in the age of onset, epidemiological studies have shown a positive correlation between age and the number of ALS cases, with the average age of diagnosis being between 55 and 65.1 [62]. Although a generally rare neurodegenerative disorder, ALS has an incidence rate of 2 in 100,000 people annually but increases for older individuals [61][62][63]. Approximately 10–15% of cases result in dual diagnoses of ALS and concomitant frontotemporal dementia (FTD), which shows a wide spectrum of overlapping symptoms and genetics [61][64]. The key differences between ALS and FTD are the targeted locations of deterioration.
Degeneration of the frontal and temporal lobes often corresponds with a range of behavioral changes, whereas ALS is defined by the deterioration of upper and lower motor neurons of the motor cortex that subsequently result in muscle paralysis and atrophy [64]. ALS is known to present itself sporadically (sALS) or genetically. Familial ALS (fALS) constitutes 5–10% of overall cases, where a single allele from a myriad of disease-causing genes will suffice for its onset due to its autosomal dominant pattern of inheritance [61][65]. Unfortunately, individuals suffering from ALS are projected to live 2–5 years after its onset [66]. Genes notorious for their involvement in the development of ALS include mutated or dysregulated ALS2, NEFH, C9orf72, SOD1, FUS, and TARDBP [61][67][68][69].
While genetic biomarkers can provide an assessment of the risk of ALS development, significant heterogeneity has also been associated with the disease, and their exact functions in ALS development and progression are unclear [61][69][70]. Additionally, many other factors are engaged in disease development and progression, including epigenetics, environment, and age-related issues such as oxidative stress [67]. Nevertheless, the average patient dies within 2–5 years [61]. Clinicians are currently restrained from providing palliative care with anti-excitatory drugs to improve patient quality of life, as there is no cure for ALS [65]. With a lack of consensus on standard and specific biomarkers, ALS cases continue to pose a grave threat. The severity of late-stage disease and ambiguity of early-stage symptoms of ALS sets grounds to demand more accurate diagnostic and prognostic biomarkers. LncRNAs have been implicated in regulatory mechanisms leading to ALS development and progression and show high tissue specificity, making them prime targets for more effective diagnostics and therapeutics [71].
LncRNA Involvement
Several studies have shown abnormal RNA metabolism from mutagenic RNA-binding proteins FUS, C9orf72, and TDP-43 to be an innate characteristic of ALS pathogenesis [71][72][73]. Paraspeckles are essential components with protective roles in the cellular stress response of motor neurons (MNs). A functional aspect of paraspeckles is their involvement in nucleoplasmic sequestration of RNA and proteins that directly alters target site expression [74]. The aggregation of paraspeckles in the CNS is a hallmark feature of ALS. LncRNA NEAT1 has inherent roles as a scaffold for paraspeckle formation [72][75]. Enriched lncRNA NEAT1_2 is proved to be the target of both FUS and TDP-43 through its UG-rich sequences; however, mutagenic or dysregulated RNA-binding proteins are associated with distorted or hyper-assembly of paraspeckles in early ALS pathogenesis [73][76]. In vivo studies have identified NEAT1 involvement in specific neurodegenerative pathways such as inflammation and neuronal cell death through p53 regulation; hence, the decreased brain density seen in brain scans and postmortem examinations of many NDD patients [68]. Of course, paraspeckles are primarily absent in healthy MNs because of decreased expression of NEAT1 isoforms essential to their assembly in the central nervous system (CNS), as studied with in vitro post-mitotic neurons [72][76]. Furthermore, NEAT1 expression has roles in neuron-specific pathways, and the aberrant hyperexcitability of affected MNs has been implicated in ALS and other NDDs [72]. The sequestering of genetic regulatory elements, including miRNAs by paraspeckle formation through abnormal NEAT1 expression in response to stressful external stimuli, such as proteasomal inhibition and viral infections, have also been identified in ALS [77]. These findings confer lncRNA NEAT1 in the CNS to be a viable target for ALS therapeutics.
Other discoveries of ncRNA’s influence in the pathophysiology of NDDs include antisense C9orf72 transcripts that have been connected to chromosome 9p-linked ALS. The C9orf72 antisense transcript appears to be highly conducive in fALS development, with 22.5% of fALS cases attributable to the hexanucleotide repeat expansion [78]. Experimental findings show various disease mechanisms the transcript acts through, though it has a significant role in nuclear RNA sequestration, which directly impacts transcription [79]. LncRNA CCND1, a FUS-bound transcript and cell proliferation regulator, is another ncRNA associated with ALS, particularly through the Wnt/β-catenin signaling pathway [80][81][82]. FUS acts as an inhibitor to CCND1 expression to regulate cell cycle progression; however, its dysregulation may trigger apoptosis in ALS [83]. Figure 4 shows the lncRNAs involved in ALS.
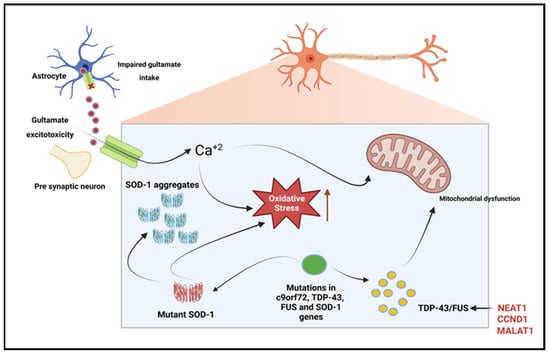
Figure 4. LncRNAs involved in amyotrophic lateral sclerosis. Due to dysfunction of the astrocytic excitatory amino acid transporter 2 (EAAT2), there is reduced uptake of glutamate from the synaptic cleft, which leads to glutamate excitotoxicity. The resulting glutamate-induced excitotoxicity induces neurodegeneration through the activation of Ca+2 dependent pathways. TDP-43, c9orf72, and fused in sarcoma (FUS) gene mutations result in dysregulated RNA metabolism, leading to the formation of intracellular neuronal aggregates. Superoxide dismutase-1 (SOD-1) gene mutations increase oxidative stress and induce mitochondrial dysfunction, leading to intracellular aggregates. Enriched lncRNA NEAT1 is proved to be the target of both FUS and TDP-43 [84]. Created with BioRender.com.
Dingsheng et al. generated an ALS-specific competitive endogenous RNA (ceRNA) network to investigate RNA transcripts with sponging effects on miRNAs to differentially regulate their expression in ALS [85]. Notably, MALAT1 was found to act as a sponge to modulate the expression of 75 genes, 7 of which have been connected to the pathogenesis of ALS by interacting with miRNA [85]. Furthermore, MALAT1, like NEAT1, is bound by TDP-43, an RNA-binding protein involved in ALS [86]. TDP-43 is noted to be a causative agent of mitochondrial dysfunction with implications in neuroinflammation, a common feature of early-stage ALS. MALAT1 is also known to regulate hnRNPA2/B1 and XIAP, both of which are closely related to apoptotic pathways that lead to neurodegeneration in ALS [85]. Experimental findings show that MALAT1 regulates the ATM gene, which is a member of the p53 apoptotic pathway in response to the DNA damage linked to the pathogenesis of ALS. Moreover, the endocytic pathway protein AAK1, which associates with SOD1 mutants and has been implicated in ALS, has also been identified as a regulatory target of MALAT1 [85]. Evidently, MALAT1 is another crucial player that can potentially have a role as a therapeutic target and a potential biomarker for ALS assessments or targeted therapies.
While a large pool of research is accessible for coding transcripts, detailed investigations of lncRNA involvement in ALS pathogenesis are in their infancy. Nevertheless, recent literature introduces differential expression of lncRNAs with impacts on transcriptional regulation pathways. ZEB1-AS1, ZBTB11-AS1, and XXbac-BPG252P9.10 are reported as novel antisense transcripts involved in ALS transcriptional regulation [25]. ZEB1-AS1 gets its significance as an antisense transcriptional regulator of Zinc Finger E-Box Binding Homeobox 1 (ZEB1), which is a highly conserved transcriptional repressor with roles in chromatin and E-Box binding; however, ZEB1-AS1 was downregulated in sALS samples in comparison to healthy control groups [85]. ZBTB11-AS1 is another differentially expressed antisense transcript coupled with the Zinc Finger and BTB Domain Counting 11 gene (ZBTB11), implying its participation in transcriptional regulation. Interestingly, cases of sALS presented with downregulated ZBTB11-AS1 [87]. XXbac-BPG252P9.10 is associated with the nuclear factor-kappa-B/REL (NF-κB) transcription factor family. The NF-κB proteins have critical roles in inflammatory and survival pathways but, in particular, IER3 is known for its regulation of anti-apoptotic genes and roles in ALS. LncRNA XXbac-BPG252P9.10 is an antisense regulator of IER3 and is significantly downregulated in sALS samples [88]. Non-coding antisense RNA expression analyses and co-expression networks involving lncRNAs and mRNAs show a multitude of potential transcripts with involvement in ALS pathogenesis that may prove to be promising therapeutic or diagnostic targets. Some relevant lncRNAs and their involvement in different neurodegenerative diseases have been summarized in Table 1.
Table 1. Summary of relevant lncRNAs and their involvement in different neurodegenerative diseases.
| LncRNA | Type of NDD | Notes | References |
|---|---|---|---|
| BACE1-AS | Alzheimer’s | BACE1-AS has been associated with regulation of BACE1, a key enzyme in amyloid β production | [37][89][90] |
| NEAT1 | Parkinson’s | Associated with modulation of neuronal apoptosis and neuro-inflammation | [37][91][92] |
| MALAT1 | Alzheimer’s | Linked with neuronal apoptosis and neuro-inflammation | [32][93][94] |
| SNHG1 | Alzheimer’s/Parkinson’s | Plays a role in regulation of amyloid β production and neuro-inflammation | [95][96][97] |
| ANRIL | Parkinson’s | Associated with vascular dysfunction and inflammation in CNS | [98] |
| HOTAIR | Alzheimer’s | Associated with dysregulation of synaptic plasticity and neuronal apoptosis, contributing to cognitive decline | [99][100] |
| TUG1 | Parkinson’s | Shown to modulate dopaminergic neuronal cell death suggesting its involvement in pathogenesis of Parkinson’s | [55][101] |
| BC200 | Parkinson’s/Multiple sclerosis | Involved in regulating mRNA translation and synaptic plasticity and contributing to disease progression | [102][103] |
| MEG3 | Alzheimer’s | Implicated in amyloid β-induced neurotoxicity and neuronal apoptosis | [104] |
| PINK-AS | Parkinson’s | Impairment of mitochondrial dynamics due to decrease in the PINK1-AS and neurodegeneration due to ASUCHL1 downregulation | [105] |
| NDM29 | Alzheimer’s | NDM29 expression is enhanced in the cerebral cortex of AD patients | [29][106] |
| LRP1-AS | Alzheimer’s | LRP1 is deeply involved in APP trafficking and Aβ processing | [107][108] |
References
- Khorkova, O.; Hsiao, J.; Wahlestedt, C. Basic biology and therapeutic implications of lncRNA. Adv. Drug Deliv. Rev. 2015, 87, 15–24.
- Rashid, F.; Shah, A.; Shan, G. Long Non-coding RNAs in the Cytoplasm. Genom. Proteom. Bioinform. 2016, 14, 73–80.
- Yu, B.; Shan, G. Functions of long noncoding RNAs in the nucleus. Nucleus 2016, 7, 155–166.
- Statello, L.; Guo, C.J.; Chen, L.L.; Huarte, M. Gene regulation by long non-coding RNAs and its biological functions. Nat. Reviews. Mol. Cell Biol. 2021, 22, 96–118.
- Lubelsky, Y.; Ulitsky, I. Sequences enriched in Alu repeats drive nuclear localization of long RNAs in human cells. Nature 2018, 555, 107–111.
- Quinn, J.J.; Chang, H.Y. Unique features of long non-coding RNA biogenesis and function. Nat. Rev. Genet. 2016, 17, 47–62.
- Jiang, M.C.; Ni, J.J.; Cui, W.Y.; Wang, B.Y.; Zhuo, W. Emerging roles of lncRNA in cancer and therapeutic opportunities. Am. J. Cancer Res. 2019, 9, 1354–1366.
- Wei, C.W.; Luo, T.; Zou, S.S.; Wu, A.S. The Role of Long Noncoding RNAs in Central Nervous System and Neurodegenerative Diseases. Front. Behav. Neurosci. 2018, 12, 175.
- Chen, J.; Wang, Y.; Wang, C.; Hu, J.F.; Li, W. LncRNA Functions as a New Emerging Epigenetic Factor in Determining the Fate of Stem Cells. Front. Genet. 2020, 11, 277.
- Bhat, S.A.; Ahmad, S.M.; Mumtaz, P.T.; Malik, A.A.; Dar, M.A.; Urwat, U.; Shah, R.A.; Ganai, N.A. Long non-coding RNAs: Mechanism of action and functional utility. Non-Coding RNA Res. 2016, 1, 43–50.
- Fernandes, J.C.R.; Acuña, S.M.; Aoki, J.I.; Floeter-Winter, L.M.; Muxel, S.M. Long Non-Coding RNAs in the Regulation of Gene Expression: Physiology and Disease. Non-Coding RNA 2019, 5, 17.
- Tripathi, M.K.; Doxtater, K.; Keramatnia, F.; Zacheaus, C.; Yallapu, M.M.; Jaggi, M.; Chauhan, S.C. Role of lncRNAs in ovarian cancer: Defining new biomarkers for therapeutic purposes. Drug Discov. Today 2018, 23, 1635–1643.
- Kung, J.T.; Colognori, D.; Lee, J.T. Long noncoding RNAs: Past, present, and future. Genetics 2013, 193, 651–669.
- Kuo, C.C.; Hänzelmann, S.; Sentürk Cetin, N.; Frank, S.; Zajzon, B.; Derks, J.P.; Akhade, V.S.; Ahuja, G.; Kanduri, C.; Grummt, I.; et al. Detection of RNA-DNA binding sites in long noncoding RNAs. Nucleic Acids Res. 2019, 47, e32.
- Fazzio, T.G. Regulation of chromatin structure and cell fate by R-loops. Transcription 2016, 7, 121–126.
- Ribeiro, D.M.; Zanzoni, A.; Cipriano, A.; Delli Ponti, R.; Spinelli, L.; Ballarino, M.; Bozzoni, I.; Tartaglia, G.G.; Brun, C. Protein complex scaffolding predicted as a prevalent function of long non-coding RNAs. Nucleic Acids Res. 2018, 46, 917–928.
- Balas, M.M.; Johnson, A.M. Exploring the mechanisms behind long noncoding RNAs and cancer. Non-Coding RNA Res. 2018, 3, 108–117.
- Chan, J.J.; Tay, Y. Noncoding RNA:RNA Regulatory Networks in Cancer. Int. J. Mol. Sci. 2018, 19, 1310.
- Li, D.; Zhang, J.; Li, X.; Chen, Y.; Yu, F.; Liu, Q. Insights into lncRNAs in Alzheimer’s disease mechanisms. RNA Biol. 2021, 18, 1037–1047.
- Cortini, F.; Roma, F.; Villa, C. Emerging roles of long non-coding RNAs in the pathogenesis of Alzheimer’s disease. Ageing Res. Rev. 2019, 50, 19–26.
- Luo, Q.; Chen, Y. Long noncoding RNAs and Alzheimer’s disease. Clin. Interv. Aging 2016, 11, 867–872.
- Wang, L.K.; Chen, X.F.; He, D.D.; Li, Y.; Fu, J. Dissection of functional lncRNAs in Alzheimer’s disease by construction and analysis of lncRNA-mRNA networks based on competitive endogenous RNAs. Biochem. Biophys. Res. Commun. 2017, 485, 569–576.
- Doxtater, K.; Tripathi, M.K.; Khan, M.M. Recent advances on the role of long non-coding RNAs in Alzheimer’s disease. Neural Regen. Res. 2020, 15, 2253–2254.
- Yang, B.; Xia, Z.A.; Zhong, B.; Xiong, X.; Sheng, C.; Wang, Y.; Gong, W.; Cao, Y.; Wang, Z.; Peng, W. Distinct Hippocampal Expression Profiles of Long Non-coding RNAs in an Alzheimer’s Disease Model. Mol. Neurobiol. 2017, 54, 4833–4846.
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; De Vos, A.; et al. The β-Secretase BACE1 in Alzheimer’s Disease. Biol. Psychiatry 2021, 89, 745–756.
- Faghihi, M.A.; Modarresi, F.; Khalil, A.M.; Wood, D.E.; Sahagan, B.G.; Morgan, T.E.; Finch, C.E.; St Laurent, G., 3rd; Kenny, P.J.; Wahlestedt, C. Expression of a noncoding RNA is elevated in Alzheimer’s disease and drives rapid feed-forward regulation of beta-secretase. Nat. Med. 2008, 14, 723–730.
- Huaying, C.; Xing, J.; Luya, J.; Linhui, N.; Di, S.; Xianjun, D. A Signature of Five Long Non-Coding RNAs for Predicting the Prognosis of Alzheimer’s Disease Based on Competing Endogenous RNA Networks. Front. Aging Neurosci. 2020, 12, 598606.
- Chen, L.; Guo, X.; Li, Z.; He, Y. Relationship between long non-coding RNAs and Alzheimer’s disease: A systematic review. Pathol. Res. Pract. 2019, 215, 12–20.
- Massone, S.; Ciarlo, E.; Vella, S.; Nizzari, M.; Florio, T.; Russo, C.; Cancedda, R.; Pagano, A. NDM29, a RNA polymerase III-dependent non coding RNA, promotes amyloidogenic processing of APP and amyloid β secretion. Biochim. Biophys. Acta 2012, 1823, 1170–1177.
- Faghihi, M.A.; Zhang, M.; Huang, J.; Modarresi, F.; Van der Brug, M.P.; Nalls, M.A.; Cookson, M.R.; St-Laurent, G., 3rd; Wahlestedt, C. Evidence for natural antisense transcript-mediated inhibition of microRNA function. Genome Biol. 2010, 11, R56.
- Chanda, K.; Jana, N.R.; Mukhopadhyay, D. Long non-coding RNA MALAT1 protects against Aβ1–42 induced toxicity by regulating the expression of receptor tyrosine kinase EPHA2 via quenching miR-200a/26a/26b in Alzheimer’s disease. Life Sci. 2022, 302, 120652.
- Zhuang, J.; Cai, P.; Chen, Z.; Yang, Q.; Chen, X.; Wang, X.; Zhuang, X. Long noncoding RNA MALAT1 and its target microRNA-125b are potential biomarkers for Alzheimer’s disease management via interactions with FOXQ1, PTGS2 and CDK5. Am. J. Transl. Res. 2020, 12, 5940–5954.
- Ma, P.; Li, Y.; Zhang, W.; Fang, F.; Sun, J.; Liu, M.; Li, K.; Dong, L. Long Non-coding RNA MALAT1 Inhibits Neuron Apoptosis and Neuroinflammation While Stimulates Neurite Outgrowth and Its Correlation With MiR-125b Mediates PTGS2, CDK5 and FOXQ1 in Alzheimer’s Disease. Curr. Alzheimer Res. 2019, 16, 596–612.
- Andersen, O.M.; Reiche, J.; Schmidt, V.; Gotthardt, M.; Spoelgen, R.; Behlke, J.; von Arnim, C.A.; Breiderhoff, T.; Jansen, P.; Wu, X.; et al. Neuronal sorting protein-related receptor sorLA/LR11 regulates processing of the amyloid precursor protein. Proc. Natl. Acad. Sci. USA 2005, 102, 13461–13466.
- Mus, E.; Hof, P.R.; Tiedge, H. Dendritic BC200 RNA in aging and in Alzheimer’s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 10679–10684.
- Li, H.; Zheng, L.; Jiang, A.; Mo, Y.; Gong, Q. Identification of the biological affection of long noncoding RNA BC200 in Alzheimer’s disease. Neuroreport 2018, 29, 1061–1067.
- Zhao, M.Y.; Wang, G.Q.; Wang, N.N.; Yu, Q.Y.; Liu, R.L.; Shi, W.Q. The long-non-coding RNA NEAT1 is a novel target for Alzheimer’s disease progression via miR-124/BACE1 axis. Neurol. Res. 2019, 41, 489–497.
- Ma, N.; Tie, C.; Yu, B.; Zhang, W.; Wan, J. Identifying lncRNA-miRNA-mRNA networks to investigate Alzheimer’s disease pathogenesis and therapy strategy. Aging 2020, 12, 2897–2920.
- Shi, Y.; Liu, H.; Yang, C.; Xu, K.; Cai, Y.; Wang, Z.; Zhao, Z.; Shao, T.; Li, Y. Transcriptomic Analyses for Identification and Prioritization of Genes Associated with Alzheimer’s Disease in Humans. Front. Bioeng. Biotechnol. 2020, 8, 31.
- Yang, S.; Yang, H.; Luo, Y.; Deng, X.; Zhou, Y.; Hu, B. Long non-coding RNAs in neurodegenerative diseases. Neurochem. Int. 2021, 148, 105096.
- Emamzadeh, F.N.; Surguchov, A. Parkinson’s Disease: Biomarkers, Treatment, and Risk Factors. Front. Neurosci. 2018, 12, 612.
- Stoker, T.B.; Torsney, K.M.; Barker, R.A. Emerging Treatment Approaches for Parkinson’s Disease. Front. Neurosci. 2018, 12, 693.
- Ni, Y.; Huang, H.; Chen, Y.; Cao, M.; Zhou, H.; Zhang, Y. Investigation of Long Non-coding RNA Expression Profiles in the Substantia Nigra of Parkinson’s Disease. Cell. Mol. Neurobiol. 2017, 37, 329–338.
- Yan, W.; Chen, Z.Y.; Chen, J.Q.; Chen, H.M. LncRNA NEAT1 promotes autophagy in MPTP-induced Parkinson’s disease through stabilizing PINK1 protein. Biochem. Biophys. Res. Commun. 2018, 496, 1019–1024.
- Liu, T.; Zhang, Y.; Liu, W.; Zhao, J. LncRNA NEAT1 Regulates the Development of Parkinson’s Disease by Targeting AXIN1 Via Sponging miR-212-3p. Neurochem. Res. 2021, 46, 230–240.
- Fan, Y.; Zhao, X.; Lu, K.; Cheng, G. LncRNA BDNF-AS promotes autophagy and apoptosis in MPTP-induced Parkinson’s disease via ablating microRNA-125b-5p. Brain Res. Bull. 2020, 157, 119–127.
- Zhang, Y.; Xia, Q.; Lin, J. LncRNA H19 Attenuates Apoptosis in MPTP-Induced Parkinson’s Disease Through Regulating miR-585-3p/PIK3R3. Neurochem. Res. 2020, 45, 1700–1710.
- Jiang, J.; Piao, X.; Hu, S.; Gao, J.; Bao, M. LncRNA H19 diminishes dopaminergic neuron loss by mediating microRNA-301b-3p in Parkinson’s disease via the HPRT1-mediated Wnt/β-catenin signaling pathway. Aging 2020, 12, 8820–8836.
- Zhou, Q.; Zhang, M.M.; Liu, M.; Tan, Z.G.; Qin, Q.L.; Jiang, Y.G. LncRNA XIST sponges miR-199a-3p to modulate the Sp1/LRRK2 signal pathway to accelerate Parkinson’s disease progression. Aging 2021, 13, 4115–4137.
- Zhao, J.; Li, H.; Chang, N. LncRNA HOTAIR promotes MPP+-induced neuronal injury in Parkinson’s disease by regulating the miR-874-5p/ATG10 axis. EXCLI J. 2020, 19, 1141–1153.
- Wang, S.; Yuan, Y.H.; Chen, N.H.; Wang, H.B. The mechanisms of NLRP3 inflammasome/pyroptosis activation and their role in Parkinson’s disease. Int. Immunopharmacol. 2019, 67, 458–464.
- Zindler, E.; Zipp, F. Neuronal injury in chronic CNS inflammation. Best Pract. Res. Clin. Anaesthesiol. 2010, 24, 551–562.
- Joshi, N.; Singh, S. Updates on immunity and inflammation in Parkinson disease pathology. J. Neurosci. Res. 2018, 96, 379–390.
- Cai, L.J.; Tu, L.; Huang, X.M.; Huang, J.; Qiu, N.; Xie, G.H.; Liao, J.X.; Du, W.; Zhang, Y.Y.; Tian, J.Y. LncRNA MALAT1 facilitates inflammasome activation via epigenetic suppression of Nrf2 in Parkinson’s disease. Mol. Brain 2020, 13, 130.
- Cheng, J.; Duan, Y.; Zhang, F.; Shi, J.; Li, H.; Wang, F.; Li, H. The Role of lncRNA TUG1 in the Parkinson Disease and Its Effect on Microglial Inflammatory Response. Neuromolecular Med. 2021, 23, 327–334.
- Cai, L.; Tu, L.; Li, T.; Yang, X.; Ren, Y.; Gu, R.; Zhang, Q.; Yao, H.; Qu, X.; Wang, Q.; et al. Downregulation of lncRNA UCA1 ameliorates the damage of dopaminergic neurons, reduces oxidative stress and inflammation in Parkinson’s disease through the inhibition of the PI3K/Akt signaling pathway. Int. Immunopharmacol. 2019, 75, 105734.
- Fields, C.R.; Bengoa-Vergniory, N.; Wade-Martins, R. Targeting Alpha-Synuclein as a Therapy for Parkinson’s Disease. Front. Mol. Neurosci. 2019, 12, 299.
- Song, Z.; Xie, B. LncRNA OIP5-AS1 reduces α-synuclein aggregation and toxicity by targeting miR-126 to activate PLK2 in human neuroblastoma SH-SY5Y cells. Neurosci. Lett. 2021, 740, 135482.
- Zhang, L.M.; Wang, M.H.; Yang, H.C.; Tian, T.; Sun, G.F.; Ji, Y.F.; Hu, W.T.; Liu, X.; Wang, J.P.; Lu, H. Dopaminergic neuron injury in Parkinson’s disease is mitigated by interfering lncRNA SNHG14 expression to regulate the miR-133b/α-synuclein pathway. Aging 2019, 11, 9264–9279.
- Bu, L.L.; Xie, Y.Y.; Lin, D.Y.; Chen, Y.; Jing, X.N.; Liang, Y.R.; Peng, S.D.; Huang, K.X.; Tao, E.X. LncRNA-T199678 Mitigates α-Synuclein-Induced Dopaminergic Neuron Injury via miR-101-3p. Front. Aging Neurosci. 2020, 12, 599246.
- Masrori, P.; Van Damme, P. Amyotrophic lateral sclerosis: A clinical review. Eur. J. Neurol. 2020, 27, 1918–1929.
- Iacovetta, C. Neurological Status. In Monitoring and Intervention for the Critically Ill Small Animal; Kirby, R., Linklater, A., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2016; p. 16.
- Longinetti, E.; Fang, F. Epidemiology of amyotrophic lateral sclerosis: An update of recent literature. Curr. Opin. Neurol. 2019, 32, 771–776.
- Chen, K.W.; Chen, J.A. Functional Roles of Long Non-coding RNAs in Motor Neuron Development and Disease. J. Biomed. Sci. 2020, 27, 38.
- Wobst, H.J.; Mack, K.L.; Brown, D.G.; Brandon, N.J.; Shorter, J. The clinical trial landscape in amyotrophic lateral sclerosis-Past, present, and future. Med. Res. Rev. 2020, 40, 1352–1384.
- Butti, Z.; Patten, S.A. RNA Dysregulation in Amyotrophic Lateral Sclerosis. Front. Genet. 2018, 9, 712.
- Agrawal, M.; Biswas, A. Molecular diagnostics of neurodegenerative disorders. Front. Mol. Biosci. 2015, 2, 54.
- Balendra, R.; Isaacs, A.M. C9orf72-mediated ALS and FTD: Multiple pathways to disease. Nat. Rev. Neurol. 2018, 14, 544–558.
- Van Damme, P.; Robberecht, W.; Van Den Bosch, L. Modelling amyotrophic lateral sclerosis: Progress and possibilities. Dis. Models Mech. 2017, 10, 537–549.
- Verma, A. Altered RNA metabolism and amyotrophic lateral sclerosis. Ann. Indian Acad. Neurol. 2011, 14, 239–244.
- Biscarini, S.; Capauto, D.; Peruzzi, G.; Lu, L.; Colantoni, A.; Santini, T.; Shneider, N.A.; Caffarelli, E.; Laneve, P.; Bozzoni, I. Characterization of the lncRNA transcriptome in mESC-derived motor neurons: Implications for FUS-ALS. Stem Cell Res. 2018, 27, 172–179.
- An, H.; Williams, N.G.; Shelkovnikova, T.A. NEAT1 and paraspeckles in neurodegenerative diseases: A missing lnc found? Non-Coding RNA Res. 2018, 3, 243–252.
- Nishimoto, Y.; Nakagawa, S.; Hirose, T.; Okano, H.J.; Takao, M.; Shibata, S.; Suyama, S.; Kuwako, K.; Imai, T.; Murayama, S.; et al. The long non-coding RNA nuclear-enriched abundant transcript 1_2 induces paraspeckle formation in the motor neuron during the early phase of amyotrophic lateral sclerosis. Mol. Brain 2013, 6, 31.
- Bond, C.S.; Fox, A.H. Paraspeckles: Nuclear bodies built on long noncoding RNA. J. Cell Biol. 2009, 186, 637–644.
- Tyzack, G.E.; Manferrari, G.; Newcombe, J.; Luscombe, N.M.; Luisier, R.; Patani, R. Paraspeckle components NONO and PSPC1 are not mislocalized from motor neuron nuclei in sporadic ALS. Brain A J. Neurol. 2020, 143, e66.
- Shelkovnikova, T.A.; Kukharsky, M.S.; An, H.; Dimasi, P.; Alexeeva, S.; Shabir, O.; Heath, P.R.; Buchman, V.L. Protective paraspeckle hyper-assembly downstream of TDP-43 loss of function in amyotrophic lateral sclerosis. Mol. Neurodegener. 2018, 13, 30.
- Jiang, L.; Shao, C.; Wu, Q.J.; Chen, G.; Zhou, J.; Yang, B.; Li, H.; Gou, L.T.; Zhang, Y.; Wang, Y.; et al. NEAT1 scaffolds RNA-binding proteins and the Microprocessor to globally enhance pri-miRNA processing. Nat. Struct. Mol. Biol. 2017, 24, 816–824.
- DeJesus-Hernandez, M.; Mackenzie, I.R.; Boeve, B.F.; Boxer, A.L.; Baker, M.; Rutherford, N.J.; Nicholson, A.M.; Finch, N.A.; Flynn, H.; Adamson, J.; et al. Expanded GGGGCC hexanucleotide repeat in noncoding region of C9ORF72 causes chromosome 9p-linked FTD and ALS. Neuron 2011, 72, 245–256.
- Quan, Z.; Zheng, D.; Qing, H. Regulatory Roles of Long Non-Coding RNAs in the Central Nervous System and Associated Neurodegenerative Diseases. Front. Cell. Neurosci. 2017, 11, 175.
- Chen, Y.; Guan, Y.; Liu, H.; Wu, X.; Yu, L.; Wang, S.; Zhao, C.; Du, H.; Wang, X. Activation of the Wnt/β-catenin signaling pathway is associated with glial proliferation in the adult spinal cord of ALS transgenic mice. Biochem. Biophys. Res. Commun. 2012, 420, 397–403.
- Gonzalez-Fernandez, C.; González, P.; Rodríguez, F.J. New insights into Wnt signaling alterations in amyotrophic lateral sclerosis: A potential therapeutic target? Neural Regen. Res. 2020, 15, 1580–1589.
- Orietta, P.; Stella, G.; Daisy, S.; Cristina, C. RNA Metabolism and Therapeutics in Amyotrophic Lateral Sclerosis. In Amyotrophic Lateral Sclerosis; Muralidhar, L.H., Ed.; IntechOpen: Rijeka, Croatia, 2019; p. Ch. 7.
- Sama, R.R.; Ward, C.L.; Bosco, D.A. Functions of FUS/TLS from DNA repair to stress response: Implications for ALS. ASN Neuro 2014, 6, 1759091414544472.
- van den Bos, M.A.J.; Geevasinga, N.; Higashihara, M.; Menon, P.; Vucic, S. Pathophysiology and Diagnosis of ALS: Insights from Advances in Neurophysiological Techniques. Int. J. Mol. Sci. 2019, 20, 2818.
- Liu, D.; Zuo, X.; Zhang, P.; Zhao, R.; Lai, D.; Chen, K.; Han, Y.; Wan, G.; Zheng, Y.; Lu, C.; et al. The Novel Regulatory Role of lncRNA-miRNA-mRNA Axis in Amyotrophic Lateral Sclerosis: An Integrated Bioinformatics Analysis. Comput. Math. Methods Med. 2021, 2021, 5526179.
- Wilusz, J.E. Long noncoding RNAs: Re-writing dogmas of RNA processing and stability. Biochim. Biophys. Acta 2016, 1859, 128–138.
- Gagliardi, S.; Zucca, S.; Pandini, C.; Diamanti, L.; Bordoni, M.; Sproviero, D.; Arigoni, M.; Olivero, M.; Pansarasa, O.; Ceroni, M.; et al. Long non-coding and coding RNAs characterization in Peripheral Blood Mononuclear Cells and Spinal Cord from Amyotrophic Lateral Sclerosis patients. Sci. Rep. 2018, 8, 2378.
- Ma, M.; Hui, J.; Zhang, Q.Y.; Zhu, Y.; He, Y.; Liu, X.J. Long non-coding RNA nuclear-enriched abundant transcript 1 inhibition blunts myocardial ischemia reperfusion injury via autophagic flux arrest and apoptosis in streptozotocin-induced diabetic rats. Atherosclerosis 2018, 277, 113–122.
- Feng, L.; Liao, Y.T.; He, J.C.; Xie, C.L.; Chen, S.Y.; Fan, H.H.; Su, Z.P.; Wang, Z. Plasma long non-coding RNA BACE1 as a novel biomarker for diagnosis of Alzheimer disease. BMC Neurol. 2018, 18, 4.
- Fotuhi, S.N.; Khalaj-Kondori, M.; Hoseinpour Feizi, M.A.; Talebi, M. Long Non-coding RNA BACE1-AS May Serve as an Alzheimer’s Disease Blood-Based Biomarker. J. Mol. Neurosci. MN 2019, 69, 351–359.
- Li, K.; Wang, Z. lncRNA NEAT1: Key player in neurodegenerative diseases. Ageing Res. Rev. 2023, 86, 101878.
- Shen, H.; Song, H.; Wang, S.; Su, D.; Sun, Q. NEAT1 enhances MPP+-induced pyroptosis in a cell model of Parkinson’s disease via targeting miR-5047/YAF2 signaling. Immun. Inflamm. Dis. 2023, 11, e817.
- Yang, H. LncRNA MALAT1 potentiates inflammation disorder in Parkinson’s disease. Int. J. Immunogenet. 2021, 48, 419–428.
- Guo, D.; Ma, J.; Yan, L.; Li, T.; Li, Z.; Han, X.; Shui, S. Down-Regulation of Lncrna MALAT1 Attenuates Neuronal Cell Death Through Suppressing Beclin1-Dependent Autophagy by Regulating Mir-30a in Cerebral Ischemic Stroke. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 2017, 43, 182–194.
- Gao, Y.; Zhang, N.; Lv, C.; Li, N.; Li, X.; Li, W. lncRNA SNHG1 Knockdown Alleviates Amyloid-β-Induced Neuronal Injury by Regulating ZNF217 via Sponging miR-361-3p in Alzheimer’s Disease. J. Alzheimer’s Dis. JAD 2020, 77, 85–98.
- Qian, C.; Ye, Y.; Mao, H.; Yao, L.; Sun, X.; Wang, B.; Zhang, H.; Xie, L.; Zhang, H.; Zhang, Y.; et al. Downregulated lncRNA-SNHG1 enhances autophagy and prevents cell death through the miR-221/222/p27/mTOR pathway in Parkinson’s disease. Exp. Cell Res. 2019, 384, 111614.
- Cao, B.; Wang, T.; Qu, Q.; Kang, T.; Yang, Q. Long Noncoding RNA SNHG1 Promotes Neuroinflammation in Parkinson’s Disease via Regulating miR-7/NLRP3 Pathway. Neuroscience 2018, 388, 118–127.
- Yang, P.; Lin, G.; Wang, M.; Chen, X.; Huang, J. Long non-coding RNA ANRIL interacts with microRNA-34a and microRNA-125a, and they all correlate with disease risk and severity of Parkinson’s disease. J. Clin. Lab. Anal. 2022, 36, e24037.
- Lu, J.; Liu, L.; Chen, J.; Zhi, J.; Li, J.; Li, L.; Jiang, Z. LncRNA HOTAIR in exercise-induced neuro-protective function in Alzheimer’s disease. Folia Neuropathol. 2022, 60, 414–420.
- Lu, J.; Liu, L.; Chen, J.; Zhi, J.; Li, J.; Li, L.; Jiang, Z. The Involvement of lncRNA HOTAIR/miR-130a-3p Axis in the Regulation of Voluntary Exercise on Cognition and Inflammation of Alzheimer’s Disease. Am. J. Alzheimer’s Dis. Other Dement. 2022, 37, 15333175221091424.
- Zhai, K.; Liu, B.; Gao, L. Long-Noncoding RNA TUG1 Promotes Parkinson’s Disease via Modulating MiR-152-3p/PTEN Pathway. Hum. Gene Ther. 2020, 31, 1274–1287.
- Lyu, Y.; Bai, L.; Qin, C. Long noncoding RNAs in neurodevelopment and Parkinson’s disease. Anim. Models Exp. Med. 2019, 2, 239–251.
- Kamal, A.; Swellam, M.; Shalaby, N.M.; Darwish, M.K.; El-Nahrery, E.M. Long non-coding RNAs BACE1-AS and BC200 in multiple sclerosis and their relation to cognitive function: A gene expression analysis. Brain Res. 2023, 1814, 148424.
- Quan, Y.; Wang, J.; Wang, S.; Zhao, J. Association of the Plasma Long Non-coding RNA MEG3 With Parkinson’s Disease. Front. Neurol. 2020, 11, 532891.
- Scheele, C.; Petrovic, N.; Faghihi, M.A.; Lassmann, T.; Fredriksson, K.; Rooyackers, O.; Wahlestedt, C.; Good, L.; Timmons, J.A. The human PINK1 locus is regulated in vivo by a non-coding natural antisense RNA during modulation of mitochondrial function. BMC Genom. 2007, 8, 74.
- Gu, L.; Guo, Z. Alzheimer’s Aβ42 and Aβ40 peptides form interlaced amyloid fibrils. J. Neurochem. 2013, 126, 305–311.
- Ulery, P.G.; Beers, J.; Mikhailenko, I.; Tanzi, R.E.; Rebeck, G.W.; Hyman, B.T.; Strickland, D.K. Modulation of beta-amyloid precursor protein processing by the low density lipoprotein receptor-related protein (LRP). Evidence that LRP contributes to the pathogenesis of Alzheimer’s disease. J. Biol. Chem. 2000, 275, 7410–7415.
- Liu, Q.; Zerbinatti, C.V.; Zhang, J.; Hoe, H.S.; Wang, B.; Cole, S.L.; Herz, J.; Muglia, L.; Bu, G. Amyloid precursor protein regulates brain apolipoprotein E and cholesterol metabolism through lipoprotein receptor LRP1. Neuron 2007, 56, 66–78.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
621
Revisions:
3 times
(View History)
Update Date:
18 Mar 2024
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No