Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Shi, D. RBPs Associated with Cardiomyopathies. Encyclopedia. Available online: https://encyclopedia.pub/entry/56149 (accessed on 24 July 2026).
Shi D. RBPs Associated with Cardiomyopathies. Encyclopedia. Available at: https://encyclopedia.pub/entry/56149. Accessed July 24, 2026.
Shi, De-Li. "RBPs Associated with Cardiomyopathies" Encyclopedia, https://encyclopedia.pub/entry/56149 (accessed July 24, 2026).
Shi, D. (2024, March 12). RBPs Associated with Cardiomyopathies. In Encyclopedia. https://encyclopedia.pub/entry/56149
Shi, De-Li. "RBPs Associated with Cardiomyopathies." Encyclopedia. Web. 12 March, 2024.
Copy Citation
Cardiomyopathies are structural and functional abnormalities of the myocardium and represent a heterogenous group of cardiac disorders, including dilated cardiomyopathy (DCM), hypertrophic cardiomyopathy (HCM), restrictive cardiomyopathy (RCM), and arrhythmogenic cardiomyopathy (ACM). RNA-binding proteins (RBPs) are major regulators of gene expression at the post-transcriptional level and contribute to generating protein abundance and diversity within a cell.
cardiomyopathy
RNA-binding protein
post-transcriptional regulation
RBM20
1. Introduction
Cardiomyopathies are structural and functional abnormalities of the myocardium and represent a heterogenous group of cardiac disorders, including dilated cardiomyopathy (DCM), hypertrophic cardiomyopathy (HCM), restrictive cardiomyopathy (RCM), and arrhythmogenic cardiomyopathy (ACM) [1][2][3]. Although these cardiomyopathy phenotypes are characterized by distinct morphological and functional traits [4], they are all closely associated with heart failure, which represents the clinical syndrome of these cardiovascular diseases, as well as the most frequent cause of hospitalization and death worldwide [5][6]. Mutations or the dysregulated expression of cardiac myofibrillar structural and functional proteins are linked to the pathogenesis of cardiomyopathies [3]. In particular, truncated variants of the giant sarcomeric protein TITIN, which has an important structural role and signaling function in cardiac physiology and disease [7], have been identified as the main genetic cause of DCM [8][9][10][11][12]. The switch from the fetal to adult TITIN isoform plays an important role in cardiac stiffness and is mediated by splicing factors that belong to the family of RNA-binding proteins (RBPs). It is well established that RNA-binding motif protein 20 (RBM20) regulates TITIN alternative splicing and that its mutations are responsible for the DCM phenotype in humans [13]. Increasing evidence suggests that other RBPs also critically contribute to the alternative splicing of sarcomeric genes. Thus, their dysfunctions are potentially involved in cardiomyopathies [14][15][16].
RBPs are major regulators of gene expression at the post-transcriptional level [17] and contribute to generating protein abundance and diversity within a cell [18][19]. They not only show dynamic expression between fetal and adult hearts under healthy conditions, but also become reactivated or repressed in heart failure [20][21], suggesting their important roles in promoting cardiac differentiation and maintaining cardiac homeostasis. Loss or the dysregulated activity of several RBPs have been closely associated with cardiomyopathies in humans or in animal models, as exemplified by RBM20 mutations in DCM patients [13]. With the expanded identification of the cardiac-specific RBPome, the contribution of RBP-mediated post-transcriptional regulation of cardiac-specific gene expression networks has gained considerable interest for better understanding the complexity of heart development and function [22][23][24][25][26]. Importantly, correcting pathogenic mutations in RBPs or manipulating their activity has the strong potential to rescue cardiomyopathy or promote cardiac regeneration after myocardial injury [27][28][29]. Therefore, RBPs have emerged as promising targets for therapeutic interventions for cardiovascular dysfunction [26][30].
2. RBPs Associated with Cardiomyopathies
The identification of gene mutations in human patients, associated with functional studies using reverse genetic approaches in animal models, such as mouse and zebrafish, has uncovered conserved as well as species-specific functions of RBPs in cardiac development. The following sections will discuss the recent understanding of well-characterized RBPs, including RBM20, RBM24, RBPMS, RBPMS2, RBFOX1, RBFOX2, and several readers of mRNA methylation, such as IGF2BP2 and YTHDC1, associated with cardiomyopathy in humans and/or animal models.
2.1. RBM20 Mutations Disrupt Cardiac-Specific Alternative Splicing in Cardiomyopathies
RBM20 contains two putative zinc finger domains and a central RNA recognition motif (RRM), followed by a highly conserved arginine/serine (RS)-rich region (Figure 1). The RRM binds to RNAs with a UCUU sequence to regulate alternative splicing [31]. Since the first report on RBM20 as a DCM-related gene [32], a large number of heterozygous missense mutations, mainly located at an arginine-serine-arginine-serine-proline (RSRSP) stretch within the RS-rich region, have been identified in DCM patients [33][34][35], and their pathogenic effects have been recently validated in animal models or by using an induced pluripotent stem cell line [28][36][37][38][39][40][41]. These mutations perturb alternative splicing of the TITIN gene and many other targets, including the DCM-related CAMK2D (calcium/calmodulin-dependent protein kinase type II delta), leading to disrupted cardiac stiffness and abnormal intracellular calcium handling [13][34][35][42]. It has been shown that changes in the RSRSP stretch affect its phosphorylation, which may be important for the nuclear localization of RBM20 and are causative in DCM [43][44]. Indeed, RBM20 proteins with mutated RS-rich regions display sarcoplasmic localization and accumulate in cytoplasmic processing bodies, thus preventing their nuclear function [40][41][45]. Recent works have proposed other mechanisms underlying the mis-localization of RBM20 RS-domain variants. One study suggests that the RS domain represents a nuclear localization signal that mediates RBM20 nuclear transport independently of phosphorylation [41]. Another study shows that disruption in the RS region impairs the physical interaction of RBM20 with Transportin-3, which functions as a nuclear importer of RBM20 [40]. Although multiple mechanisms may account for the dysregulated post-transcriptional activity of mutant RBM20 proteins, it is particularly interesting that correcting these pathogenic variants through precise base editing or promoting the interaction of RBM20 with its nuclear importer can redirect RBM20 nuclear localization and rescue DCM, at least in animal models [27][28][40].

Figure 1. Schematic representation of human RBM20 protein structure. Functional domains are indicated by colored boxes (P-rich, proline-rich region; L-rich, leucine-rich region; E-rich, glutamate-rich region; ZnF, zinc finger domain) and the alignment of amino acids in the RS-rich region from three vertebrate species (human, NP_001127835.2; mouse, NP_001164318.1; zebrafish, XP_021325295.1). Several validated pathogenic mutations in the RSRSP stretch are shown under the schema (green arrowhead). Conserved residues are shadowed in grey. The RS-rich region is indicated in green and the putative nuclear localization signal in the RS-rich region is underlined [41]. Note that the R634L variant (blue) co-segregates with left ventricular non-compaction cardiomyopathy [46] and the R636H variant (orange) may be also associated with HCM [47].
The human TITIN gene contains 364 exons with 363 coding ones. It produces various protein isoforms with different functions through alternative splicing [48]. Cardiac muscle expresses two main TITIN isoforms: a more compliant N2BA isoform with a longer extensible I-band region and a stiff N2B isoform, which serve as long and short molecular springs, respectively [49]. Different proportions of these isoforms expressed in fetal and adult hearts confer the passive stiffness of cardiac muscle [48][50]. However, changes in their relative levels can affect cardiac function and lead to DCM. RBM20 functions as a splicing repressor of TITIN pre-mRNA [51][52]. It prevents the inclusion of exons 51–218, which encode I-band regions, and leads to the expression of the shorter and stiffer N2B isoform of TITIN [10]. Thus, the dysfunction of RBM20 affects TITIN isoform switching during heart development and disrupts cardiomyocyte stiffness. Nevertheless, mis-localized pathogenic RBM20 variants in DCM not only retain their splicing activity [40], but also mediate distinct mRNA interactions to alter other processes of post-transcriptional regulation that occur in the cytoplasm, such as circular RNA production and alternative polyadenylation [45]. Therefore, RBM20 mutations can cause DCM through splicing-dependent and splicing-independent mechanisms.
There is also evidence that RBM20 may be a candidate gene for HCM [47][53]. Exome sequencing of HCM patients has identified deleterious variants of RBM20, but it seems that these mutations affect other structural and functional domains other than those identified in DCM patients [53]. Thus, it is still unclear how they change RBM20 localization and post-transcriptional regulatory activity. A more recent report shows that the R636H pathogenic variant of RBM20 associated with DCM is also present in HCM [47]. These observations suggest that, to some extent, RBM20 may contribute to HCM through a similar mechanism as in DCM. Further studies are necessary to firmly establish the causal relationship between RBM20 mutations and the pathogenesis of HCM.
2.2. RBM24 Is Associated with DCM
Rbm24 is highly conserved from C. elegans to mammals [54], harboring a single RRM at the N-terminal region and a more divergent C-terminal half but with several characteristic motifs (Figure 2). The RRM binds to AU/U-rich ligands that are present in a wide spectrum of target mRNAs [55]. In vertebrate embryos, the heart-specific expression of Rbm24 can be detected as early as in the heart fields or the cardiac crescent [56][57][58][59][60], and this is activated after the expression of Nkx2.5 [56], suggesting that Rbm24 may be induced by and function downstream of cardiac-specific transcription factors. Zebrafish have two rbm24 paralogs, rbm24a and rbm24b. Rbm24a protein exhibits a higher degree of overall identity and shows an identical expression pattern to Rbm24 in other vertebrates [54][57].

Figure 2. Schematic representation of human RBM24 protein structure and the amino acid alignment of the C-terminal region from three vertebrate species (human, NP_001137414.1; mouse, NP_001074894.1; zebrafish, NP_998030.1). The RRM is indicated in purple and the eukaryotic initiation factor 4E (eIF4E)-binding motif is shown by red color box. Conserved residues are shadowed in grey. The phosphorylable serine residue (S) in the eIF4E-binding motif is indicated in red [61].
The loss of Rbm24 in zebrafish and mice produces a strong phenotype of heart abnormalities, characterized by defective looping, the occurrence of cardiac edema, and impaired valve development [58][61][62][63]. The knockout of Rbm24 in mice leads to the complete exclusion or minor inclusion of muscle-specific exons in E11.5 embryos [63]. Importantly, the affected genes are closely associated with cardiogenesis, sarcomere assembly, and the pathogenesis of DCM, and include Naca, Fxr1, Abcc9, Usp25, and Usp28. This is further supported by the observation that the knockdown or knockout of rbm24 paralogs in zebrafish impairs myofibrillar integrity by disrupting sarcomere organization [60]. Mechanistically, Rbm24 prevents the repression of exon inclusion mediated by PTBP1 (polypyrimidine tract binding protein 1) and hnRNP A1/A2 (heterogeneous nuclear ribonucleoprotein A1/A2) through binding to an intronic splicing enhancer and functioning as an activator of cardiac muscle-specific exon inclusion [63]. The heart-specific conditional knockout of Rbm24 in postnatal mice disrupts the isoform transition of Titin protein, leading to progressive DCM [64]. Different from RBM20, the loss of RBM24 in human embryonic stem cells or in mice prevents the inclusion of exons coding for the N-terminal Z-repeat domain of TITIN, which binds to the major Z-disc component α-ACTININ 2 [64][65]. In addition, at early stages of induced cardiac differentiation from human embryonic stem cells, RBM24 is also involved in the alternative splicing of α-ACTININ 2 transcripts. It promotes the inclusion of exon 6, which codes for an actin-binding domain. This facilitates the TITIN-mediated interaction of α-ACTININ 2 with cardiac-specific MYH6, which can subsequently substitute non-muscle MYH10 [65]. Therefore, RBM24 plays a conserved role in cardiac myofibrillogenesis by regulating sarcomere assembly and integrity.
It is of note that Rbm24 is also involved in regulating CAMK2D splicing. The deletion of Rbm24 in mice causes aberrant expression of the CAMK2C isoform, leading to alterations in calcium handling and prolongation of the QT interval [66]. Thus, Rbm24 deficiency may cause electrophysiological abnormalities and disrupt cardiac rhythm.
In addition to the regulation of isoform switching for sarcomeric proteins, Rbm24 also functions in other post-transcriptional processes to promote heart development. It has been shown that Rbm24 represents a transcriptional target and a translational regulator of p53 in the mouse embryonic heart. In Rbm24 homozygous mutant mice, the aberrant activation of p53-dependent apoptosis partly contributes to endocardial cushion defects, growth retardation, and embryonic lethality at E12.5–13.5 [61]. Vertebrate Rbm24 proteins display a highly conserved short amino acid stretch (residues 175–199 in human RBM24), which interacts with eukaryotic initiation factor 4E (eIF4E) to prevent its association with the 5′-cap of p53 mRNA and assembly of the translation initiation complex, thereby repressing p53 mRNA translation [61]. However, phosphorylation of the serine residue in the eIF4E-binding motif of Rbm24 releases its interaction with eIF4E, thus rendering it as a translational activator [61]. This suggests that Rbm24 regulates heart development, function, and homeostasis through interaction with its partners and that the eIF4E-binding motif may represent a potential therapeutic target for modulating Rbm24 activity. The regulation of mRNA translation is consistent with the dynamic subcellular localization of Rbm24 during muscle differentiation. At least in skeletal muscle, Rbm24 is localized in the cytoplasm of fate-committed myoblasts but is translocated in the nucleus of differentiated myofibers [67]. Whether and how Rbm24 undergoes cytoplasm-to-nuclear translocation during cardiomyocyte differentiation merits future investigation.
Although the involvement of RBM24 in human heart disease remains to be determined, there is evidence showing the up-regulation of RBM24 in heart failure, which is associated with fetal-specific gene expression and protein isoform switching [20]. Given the strikingly conserved cardiac-specific expression of RBM24 during early development, it will be of interest to decipher the origin and mechanism of cardiac defects due to RBM24 deficiency, such as the impaired differentiation of progenitor cells, the disrupted morphogenesis of heart chambers or endocardial cushions, or the direct dysregulation of cardiac structural and functional genes. There is a possibility that Rbm24 displays dynamic functions dependent on its subcellular localization and its interaction with protein partners.
2.3. Loss of RBPMS and RBPMS2 Causes DCM or HCM
RNA Binding Protein with Multiple Splicing (RBPMS) variants and RBPMS2 cluster into two related families of RBPs with an RRM near the N-terminal region [22][68]. The RRM binds tandem CAC motifs with a variable spacer in mRNA targets [69]. RBPMS and RBPMS2 are expressed in the myocardium during development and in the adult of different vertebrate species [68]. Recent studies in vertebrate models have demonstrated their potential involvement in different types of cardiomyopathies. Rbpms plays an important role in post-natal cardiac function. The constitutive knockout of Rbpms in mice leads to excessive trabeculation associated with reduced thickness of the ventricles, reminiscent of non-compaction cardiomyopathy [70]. The cardiac-specific deletion of Rbpms in mice causes severe defects in cardiomyocyte contraction, resulting in DCM and early lethality in the adult [71]. The loss of Rbpms in the heart also disrupts the alternative splicing of sarcomeric genes, including Titin and Pdlim5, by preventing the inclusion of exons. In conditional Rbpms knockout hearts, the skipping of exons 11 and 47 in the Titin transcript leads to the expression of truncated N2BA and N2B isoforms, which may contribute to impaired cardiomyocyte contractility. The same splicing defects are also present in RBPMS-deficient cardiomyocytes derived from human-induced pluripotent stem cells [71]. This suggests the conserved function of RBPMS in cardiac function and raises the possibility that mutations or the dysfunction of RBPMS may contribute to DCM in humans.
In zebrafish, both rbpms2a and rbpms2b are enriched in the nkx2.5-positive cell population. The simultaneous loss of rbpms2a and rbpms2b does not seem to impact cardiac morphogenesis but causes cardiac edema and early cardiac dysfunction with reduced ejection fraction [72]. RNA-seq analysis indicates that Rbpms2 regulates the alternative splicing of several genes involved in cardiac function, such as mybpc3 and myom2a, which encode sarcomeric proteins. Consistently, rbpms2 mutant zebrafish display disorganized myofibrillar arrays and defective calcium handling [72]. Importantly, MYBPC3 mutations are most frequently detected in patients with HCM [73], and MYOM2 mutations are also identified in patients with HCM, as well as Tetralogy of Fallot [74]. It is of note that RBPMS2-deficient human cardiomyocytes also display the same defective myofibril structure and calcium handling as zebrafish rbpms2 mutants [72], suggesting the conserved molecular and cellular function of this RBP in heart development and disease.
2.4. Rbfox1 and Rbfox2 Are Involved in the Pathogenesis of Heart Disease
The RNA binding forkhead box homolog (RBFOX) family proteins have three members in vertebrates (Rbfox1, Rbfox2, and Rbfox3). They display an evolutionarily conserved RRM flanked by diversified N- and C-terminal regions (Figure 3). The RRM from all three Rbfox proteins binds to the UGCAUG motif in target mRNAs [75]. Functional studies indicate that Rbfox1 and Rbfox2 are involved in the regulation of heart development and homeostasis. RBFOX1 expression is markedly decreased in human DCM hearts and its deficiency causes pressure overload-induced heart failure in a mouse model. Transcriptomic profiling reveals that Rbfox1 functions as a prominent regulator of cardiac-specific alternative splicing and is required for isoform switching of the transcription factor MEF2 [76]. Consistently, recent studies further show that Rbfox1 mediates mRNA splicing to promote cardiomyocyte maturation [77].
Similarly, RBFOX2 regulates a large program of cardiac-specific alternative splicing events that are important for heart function. In diabetic patients, the disruption of RBFOX2 homeostasis in the heart leads to autoregulation of its exon 6 exclusion [78]. This exon codes for the second half of the RRM, and its in-frame deletion leads to the expression of a truncated protein isoform that acts as a dominant negative mutant to repress Rbfox-regulated alternative splicing [79]. The dysregulation of RBFOX2 activity affects the gene function associated with cytoskeleton and calcium handling, leading to cardiac complications before the onset of diabetic cardiomyopathy [78]. Importantly, the heterozygous loss of RBFOX2 function results from de novo frameshift, nonsense, or splice site mutations, and the reduced expression of RBFOX2 due to copy number loss is significantly enriched in patients with hypoplastic left heart syndrome (HLHS) displaying the left ventricle obstruction phenotype [80][81][82][83]. The frameshift mutation of RBFOX2 is predicted to introduce a premature stop codon, whereas the splice site mutation is expected to include intron 10 in the mRNA. Nonsense mutation truncates a portion of the C-terminal region that includes a sequence coded by an alternative exon 11 (Figure 3). This domain contains several tyrosine residues involved in RBFOX2 subcellular localization [82] and the assembly of a higher-order complex of proteins to enhance RBFOX2 splicing activity [84]. Consistent with the requirement for heart function in humans, the knockout of Rbfox2 in mice causes DCM, followed by heart failure [85]. Recently, it has been shown that Rbfox2 is involved in regulating the alternative splicing events that influence the communication between cardiomyocytes and the extracellular matrix. The loss of Rbfox2 in mice produces cardiovascular development defects similar to those observed in HLHS [86].
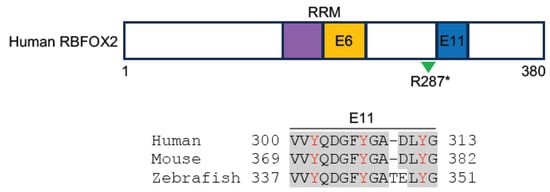
Figure 3. Schematic representation of human RBFOX2 protein structure and the alignment of sequences involved in RBFOX2 subcellular localization and formation of the multiprotein complex (human, NM_001031695.2; mouse, NP_444334.3; zebrafish, XP_021332501.1). These sequences are coded by exon 11 (blue box) in human and mouse and by exon 9 in zebrafish. Conserved residues are shadowed in grey; Tyrosine residues essential for RBFOX2 subcellular localization and complex formation are indicated in red [83][84]. The RRM is indicated in purple- and orange-colored boxes. Splicing-regulated exon 6 (E6), which codes for the second half of the RRM, and the R287* nonsense mutation identified in HLHS patients (green arrowhead) are also shown.
Zebrafish have five rbfox genes (rbfox1, rbfox1l, rbfox2, rbfox3a, and rbfox3b). The knockdown of rbfox1 alters the cardiac-specific isoform expression of several key proteins regulating cardiomyocyte structure and function [87]. Further confirming the causal relationship of RBFOX2 loss of function with HLHS, zebrafish mutants lacking rbfox1l and rbfox2 develop cardiovascular defects similar to those observed in HLHS patients, including the reduced size and disorganized architecture of ventricular cardiomyocytes, an obstructed aorta, and the absence of endocardial cushions [88]. Mechanistically, the loss of rbfox1l and rbfox2 affects the expression and alternative splicing of genes involved in sarcomere assembly and mitochondrial respiration. It is of note that cardiac defects in rbfox1l and rbfox2 mutants can be rescued by human RBFOX2 but not by HLHS-related RBFOX2 mutant variants [88]. Thus, these observations demonstrate the conserved functions of RBFOX proteins in heart development and cardiovascular disease.
2.5. RNA-Binding Proteins Associated with RNA Methylation in Heart Disease
N6-methyladenosine (m6A) is thought to be the most abundant modification in eukaryotic mRNAs, and this modification plays a central role in the post-transcriptional regulation of gene expression [89][90]. It is a dynamic and reversible process coordinated by methyltransferases, such as METTL3/14/16, and demethylases, including FTO and ALKBH5. Several RBP readers, including IGF2BP1/2/3, YTHDC1/2, YTHDF1/2/3, and hnRNPA2B1/C, function to recognize m6A-modified target transcripts and contribute to their alternative splicing, translocation, stability, and translation [90]. Evidence is accumulating that the regulation of mRNA modification is closely associated with cardiovascular disease [91].
IGF2BP2 (insulin-like growth factor 2 mRNA-binding protein 2) shows higher expression in the hearts of DCM patients [92]. The transgenic expression of human IGF2BP2 in mouse hearts causes DCM by inhibiting the expression of sarcomeric and mitochondrial proteins, leading to disorganized sarcomeres and fragmented mitochondria [92]. Nevertheless, the overexpression of IGF2BP2 has no effect on the expression of Titin and Mybpc3, suggesting that it regulates distinct sets of sarcomeric genes. There is a possibility that IGF2BP2-induced cardiac remodeling represents an adaptive response to cardiac stress. Yet, it is still unclear whether this is mediated by interactions with the methylation machinery. Thus, the post-transcriptional mechanism underlying IGF2BP2 function in heart disease requires further investigation, and it will be interesting to examine whether modulating the expression or activity of IGF2BP2 could rescue cardiac dysfunction in DCM.
The cardiac-specific conditional knockout of YTHDC1 in mice causes DCM by inducing left ventricular chamber enlargement and disorganized sarcomere arrangement, leading to decreased cardiomyocyte contractility and severe systolic dysfunction [93]. Mechanistically, YTHDC1 binds to m6A-modified Titin mRNA, and the loss of YTHDC1 increases the expression ratio of N2BA to N2B Titin isoforms [93]. This suggests that YTHDC1 may be involved in regulating cardiac stiffness by facilitating the inclusion of exons that code for the more compliant N2BA isoform. Although it remains to be determined whether other m6A readers of the methylation machinery are involved in cardiomyopathy, several lines of recent evidence indicate that YTHDF1, YTHDF2, and YTHDF3, which either promote mRNA translation or reduce mRNA stability [89], are required for heart development and function. YTHDF1 and YTHDF3 coordinate the differentiation of cardiomyocytes from mouse embryonic stem cells [94], and YTHDF1 can suppress cardiac hypertrophy in an m6A-dependent manner [95]. The loss of YTHDF2 in the cardiomyocytes of adult mice leads to abnormal cardiac remodeling and cardiac function by disrupting the stability and translation of its m6A-modified target transcripts, suggesting that it plays a role in cardiac homeostasis [96][97]. There is also evidence showing that hnRNPC is up-regulated in the sarcomeric Z-disc and likely plays a role in the pathological remodeling of the extracellular matrix [98]. Altogether, these data suggest that the interactions between m6A readers and their mRNA targets contribute to maintain cardiomyocyte integrity and cardiac contractile function.
References
- Braunwald, E. Cardiomyopathies: An overview. Circ. Res. 2017, 121, 711–721.
- Brieler, J.; Breeden, M.A.; Tucker, J. Cardiomyopathy: An overview. Am. Fam. Physician 2017, 96, 640–646.
- Ciarambino, T.; Menna, G.; Sansone, G.; Giordano, M. Cardiomyopathies: An overview. Int. J. Mol. Sci. 2021, 22, 7722.
- Arbelo, E.; Protonotarios, A.; Gimeno, J.R.; Arbustini, E.; Barriales-Villa, R.; Basso, C.; Bezzina, C.R.; Biagini, E.; Blom, N.A.; de Boer, R.A.; et al. 2023 ESC Guidelines for the management of cardiomyopathies. Eur. Heart J. 2023, 44, 3503–3626.
- Seferović, P.M.; Polovina, M.; Bauersachs, J.; Arad, M.; Ben Gal, T.; Lund, L.H.; Felix, S.B.; Arbustini, E.; Caforio, A.L.P.; Farmakis, D.; et al. Heart failure in cardiomyopathies: A position paper from the Heart Failure Association of the European Society of Cardiology. Eur. J. Heart Fail. 2019, 21, 553–576.
- Ziaeian, B.; Fonarow, G.C. Epidemiology and aetiology of heart failure. Nat. Rev. Cardiol. 2016, 13, 368–378.
- Eldemire, R.; Tharp, C.A.; Taylor, M.R.G.; Sbaizero, O.; Mestroni, L. The sarcomeric spring protein Titin: Biophysical properties, molecular mechanisms, and genetic mutations associated with heart failure and cardiomyopathy. Curr. Cardiol. Rep. 2021, 23, 121.
- Fomin, A.; Gärtner, A.; Cyganek, L.; Tiburcy, M.; Tuleta, I.; Wellers, L.; Folsche, L.; Hobbach, A.J.; von Frieling-Salewsky, M.; Unger, A.; et al. Truncated titin proteins and titin haploinsufficiency are targets for functional recovery in human cardiomyopathy due to TTN mutations. Sci. Transl. Med. 2021, 13, eabd3079.
- Herman, D.S.; Lam, L.; Taylor, M.R.; Wang, L.; Teekakirikul, P.; Christodoulou, D.; Conner, L.; DePalma, S.R.; McDonough, B.; Sparks, E.; et al. Truncations of titin causing dilated cardiomyopathy. N. Engl. J. Med. 2012, 366, 619–628.
- Loescher, C.M.; Hobbach, A.J.; Linke, W.A. Titin (TTN): From molecule to modifications, mechanics, and medical significance. Cardiovasc. Res. 2022, 118, 2903–2918.
- McAfee, Q.; Chen, C.Y.; Yang, Y.; Caporizzo, M.A.; Morley, M.; Babu, A.; Jeong, S.; Brandimarto, J.; Bedi, K.C., Jr.; Flam, E.; et al. Truncated titin proteins in dilated cardiomyopathy. Sci. Transl. Med. 2021, 13, eabd7287.
- Vikhorev, P.G.; Vikhoreva, N.N.; Yeung, W.; Li, A.; Lal, S.; Dos Remedios, C.G.; Blair, C.A.; Guglin, M.; Campbell, K.S.; Yacoub, M.H.; et al. Titin-truncating mutations associated with dilated cardiomyopathy alter length-dependent activation and its modulation via phosphorylation. Cardiovasc. Res. 2022, 118, 241–253.
- Guo, W.; Schäfer, S.; Greaser, M.L.; Radke, M.H.; Liss, M.; Govindarajan, T.; Maatz, H.; Schulz, H.; Li, S.; Parrish, A.M.; et al. RBM20, a gene for hereditary cardiomyopathy, regulates titin splicing. Nat. Med. 2012, 18, 766–773.
- Beqqali, A. Alternative splicing in cardiomyopathy. Biophys. Rev. 2018, 10, 1061–1071.
- Hasimbegovic, E.; Schweiger, V.; Kastner, N.; Spannbauer, A.; Traxler, D.; Lukovic, D.; Gyöngyösi, M.; Mester-Tonczar, J. Alternative splicing in cardiovascular disease-A survey of recent findings. Genes 2021, 12, 1457.
- Weeland, C.J.; van den Hoogenhof, M.M.; Beqqali, A.; Creemers, E.E. Insights into alternative splicing of sarcomeric genes in the heart. J. Mol. Cell. Cardiol. 2015, 81, 107–113.
- Shi, D.L.; Grifone, R. RNA-binding proteins in the post-transcriptional control of skeletal muscle development, regeneration and disease. Front. Cell Dev. Biol. 2021, 9, 738978.
- Gebauer, F.; Schwarzl, T.; Valcárcel, J.; Hentze, M.W. RNA-binding proteins in human genetic disease. Nat. Rev. Genet. 2021, 22, 185–198.
- Glisovic, T.; Bachorik, J.L.; Yong, J.; Dreyfuss, G. RNA-binding proteins and post-transcriptional gene regulation. FEBS Lett. 2008, 582, 1977–1986.
- D’Antonio, M.; Nguyen, J.P.; Arthur, T.D.; Matsui, H.; Donovan, M.K.R.; D’Antonio-Chronowska, A.; Frazer, K.A. In heart failure reactivation of RNA-binding proteins is associated with the expression of 1,523 fetal-specific isoforms. PLoS Comput. Biol. 2022, 18, e1009918.
- Riechert, E.; Kmietczyk, V.; Stein, F.; Schwarzl, T.; Sekaran, T.; Jürgensen, L.; Kamuf-Schenk, V.; Varma, E.; Hofmann, C.; Rettel, M.; et al. Identification of dynamic RNA-binding proteins uncovers a Cpeb4-controlled regulatory cascade during pathological cell growth of cardiomyocytes. Cell Rep. 2021, 35, 109100.
- Blech-Hermoni, Y.; Ladd, A.N. RNA binding proteins in the regulation of heart development. Int. J. Biochem. Cell Biol. 2013, 45, 2467–2478.
- de Bruin, R.G.; Rabelink, T.J.; van Zonneveld, A.J.; van der Veer, E.P. Emerging roles for RNA-binding proteins as effectors and regulators of cardiovascular disease. Eur. Heart J. 2017, 38, 1380–1388.
- Kelaini, S.; Chan, C.; Cornelius, V.A.; Margariti, A. RNA-binding proteins hold key roles in function, dysfunction, and disease. Biology 2021, 10, 366.
- Ruffenach, G.; Medzikovic, L.; Sun, W.; Hong, J.; Eghbali, M. Functions of RNA-binding proteins in cardiovascular disease. Cells 2023, 12, 2794.
- Völkers, M.; Preiss, T.; Hentze, M.W. RNA-binding proteins in cardiovascular biology and disease: The beat goes on. Nat. Rev. Cardiol. 2024.
- Grosch, M.; Schraft, L.; Chan, A.; Küchenhoff, L.; Rapti, K.; Ferreira, A.M.; Kornienko, J.; Li, S.; Radke, M.H.; Krämer, C.; et al. Striated muscle-specific base editing enables correction of mutations causing dilated cardiomyopathy. Nat. Commun. 2023, 14, 3714.
- Nishiyama, T.; Zhang, Y.; Cui, M.; Li, H.; Sanchez-Ortiz, E.; McAnally, J.R.; Tan, W.; Kim, J.; Chen, K.; Xu, L.; et al. Precise genomic editing of pathogenic mutations in RBM20 rescues dilated cardiomyopathy. Sci. Transl. Med. 2022, 14, eade1633.
- Shi, D.L. RNA-binding proteins as critical post-transcriptional regulators of cardiac regeneration. Int. J. Mol. Sci. 2023, 24, 12004.
- Cornelius, V.A.; Naderi-Meshkin, H.; Kelaini, S.; Margariti, A. RNA-binding proteins: Emerging therapeutics for vascular dysfunction. Cells 2022, 11, 2494.
- Upadhyay, S.K.; Mackereth, C.D. Structural basis of UCUU RNA motif recognition by splicing factor RBM20. Nucleic Acids Res. 2020, 48, 4538–4550.
- Brauch, K.M.; Karst, M.L.; Herron, K.J.; de Andrade, M.; Pellikka, P.A.; Rodeheffer, R.J.; Michels, V.V.; Olson, T.M. Mutations in ribonucleic acid binding protein gene cause familial dilated cardiomyopathy. J. Am. Coll. Cardiol. 2009, 54, 930–941.
- Watanabe, T.; Kimura, A.; Kuroyanagi, H. Alternative splicing regulator RBM20 and cardiomyopathy. Front. Mol. Biosci. 2018, 5, 105.
- Koelemen, J.; Gotthardt, M.; Steinmetz, L.M.; Meder, B. RBM20-related cardiomyopathy: Current understanding and future options. J. Clin. Med. 2021, 10, 4101.
- Lennermann, D.; Backs, J.; van den Hoogenhof, M.M.G. New insights in RBM20 cardiomyopathy. Curr. Heart Fail. Rep. 2020, 17, 234–246.
- Eberl, H.; Rebs, S.; Hoppe, S.; Sedaghat-Hamedani, F.; Kayvanpour, E.; Meder, B.; Streckfuss-Bömeke, K. Generation of an RBM20-mutation-associated left-ventricular non-compaction cardiomyopathy iPSC line (UMGi255-A) into a DCM genetic background to investigate monogenetic cardiomyopathies. Stem Cell Res. 2024, 74, 103290.
- Ihara, K.; Sasano, T.; Hiraoka, Y.; Togo-Ohno, M.; Soejima, Y.; Sawabe, M.; Tsuchiya, M.; Ogawa, H.; Furukawa, T.; Kuroyanagi, H. A missense mutation in the RSRSP stretch of Rbm20 causes dilated cardiomyopathy and atrial fibrillation in mice. Sci. Rep. 2020, 10, 17894.
- Schneider, J.W.; Oommen, S.; Qureshi, M.Y.; Goetsch, S.C.; Pease, D.R.; Sundsbak, R.S.; Guo, W.; Sun, M.; Sun, H.; Kuroyanagi, H.; et al. Dysregulated ribonucleoprotein granules promote cardiomyopathy in RBM20 gene-edited pigs. Nat. Med. 2020, 26, 1788–1800.
- Wang, C.; Zhang, Y.; Methawasin, M.; Braz, C.U.; Gao-Hu, J.; Yang, B.; Strom, J.; Gohlke, J.; Hacker, T.; Khatib, H.; et al. RBM20S639G mutation is a high genetic risk factor for premature death through RNA-protein condensates. J. Mol. Cell. Cardiol. 2022, 165, 115–129.
- Kornienko, J.; Rodríguez-Martínez, M.; Fenzl, K.; Hinze, F.; Schraivogel, D.; Grosch, M.; Tunaj, B.; Lindenhofer, D.; Schraft, L.; Kueblbeck, M.; et al. Mislocalization of pathogenic RBM20 variants in dilated cardiomyopathy is caused by loss-of-interaction with Transportin-3. Nat. Commun. 2023, 14, 4312.
- Zhang, Y.; Gregorich, Z.R.; Wang, Y.; Braz, C.U.; Zhang, J.; Liu, Y.; Liu, P.; Shen, J.; Aori, N.; Hacker, T.A.; et al. Disruption of the nuclear localization signal in RBM20 is causative in dilated cardiomyopathy. JCI Insight 2023, 8, e170001.
- van den Hoogenhof, M.M.G.; Beqqali, A.; Amin, A.S.; van der Made, I.; Aufiero, S.; Khan, M.A.F.; Schumacher, C.A.; Jansweijer, J.A.; van Spaendonck-Zwarts, K.Y.; Remme, C.A.; et al. RBM20 mutations induce an arrhythmogenic dilated cardiomyopathy related to disturbed calcium handling. Circulation 2018, 138, 1330–1342.
- Gaertner, A.; Klauke, B.; Felski, E.; Kassner, A.; Brodehl, A.; Gerdes, D.; Stanasiuk, C.; Ebbinghaus, H.; Schulz, U.; Dubowy, K.O.; et al. Cardiomyopathy-associated mutations in the RS domain affect nuclear localization of RBM20. Hum. Mutat. 2020, 41, 1931–1943.
- Murayama, R.; Kimura-Asami, M.; Togo-Ohno, M.; Yamasaki-Kato, Y.; Naruse, T.K.; Yamamoto, T.; Hayashi, T.; Ai, T.; Spoonamore, K.G.; Kovacs, R.J.; et al. Phosphorylation of the RSRSP stretch is critical for splicing regulation by RNA-Binding Motif Protein 20 (RBM20) through nuclear localization. Sci. Rep. 2018, 8, 8970.
- Fenix, A.M.; Miyaoka, Y.; Bertero, A.; Blue, S.M.; Spindler, M.J.; Tan, K.K.B.; Perez-Bermejo, J.A.; Chan, A.H.; Mayerl, S.J.; Nguyen, T.D.; et al. Gain-of-function cardiomyopathic mutations in RBM20 rewire splicing regulation and re-distribute ribonucleoprotein granules within processing bodies. Nat. Commun. 2021, 12, 6324.
- Sedaghat-Hamedani, F.; Haas, J.; Zhu, F.; Geier, C.; Kayvanpour, E.; Liss, M.; Lai, A.; Frese, K.; Pribe-Wolferts, R.; Amr, A.; et al. Clinical genetics and outcome of left ventricular non-compaction cardiomyopathy. Eur. Heart J. 2017, 38, 3449–3460.
- Inagaki, N.; Hayashi, T.; Takei, Y.; Kosuge, H.; Suzuki, S.; Tanimoto, K.; Chikamori, T.; Kimura, A. Pathogenic variant of RBM20 in a multiplex family with hypertrophic cardiomyopathy. Hum. Genome Var. 2022, 9, 6.
- Guo, W.; Bharmal, S.J.; Esbona, K.; Greaser, M.L. Titin diversity--alternative splicing gone wild. J. Biomed. Biotechnol. 2010, 2010, 753675.
- Tharp, C.A.; Haywood, M.E.; Sbaizero, O.; Taylor, M.R.G.; Mestroni, L. The giant protein Titin’s role in cardiomyopathy: Genetic, transcriptional, and post-translational modifications of TTN and their contribution to cardiac disease. Front. Physiol. 2019, 10, 1436.
- Neagoe, C.; Opitz, C.A.; Makarenko, I.; Linke, W.A. Gigantic variety: Expression patterns of titin isoforms in striated muscles and consequences for myofibrillar passive stiffness. J. Muscle Res. Cell Motil. 2003, 24, 175–189.
- Li, S.; Guo, W.; Dewey, C.N.; Greaser, M.L. Rbm20 regulates titin alternative splicing as a splicing repressor. Nucleic Acids Res. 2013, 41, 2659–2672.
- Maatz, H.; Jens, M.; Liss, M.; Schafer, S.; Heinig, M.; Kirchner, M.; Adami, E.; Rintisch, C.; Dauksaite, V.; Radke, M.H.; et al. RNA-binding protein RBM20 represses splicing to orchestrate cardiac pre-mRNA processing. J. Clin. Investig. 2014, 124, 3419–3430.
- Dai, J.; Li, Z.; Huang, W.; Chen, P.; Sun, Y.; Wang, H.; Wu, D.; Chen, Y.; Li, C.; Xiao, L.; et al. RBM20 Is a candidate gene for hypertrophic cardiomyopathy. Can. J. Cardiol. 2021, 37, 1751–1759.
- Grifone, R.; Shao, M.; Saquet, A.; Shi, D.L. RNA-binding protein Rbm24 as a multifaceted post-transcriptional regulator of embryonic lineage differentiation and cellular homeostasis. Cells 2020, 9, 1891.
- Ray, D.; Kazan, H.; Chan, E.T.; Peña-Castillo, L.; Chaudhry, S.; Talukder, S.; Blencowe, B.J.; Morris, Q.; Hughes, T.R. Rapid and systematic analysis of the RNA recognition specificities of RNA-binding proteins. Nat. Biotechnol. 2009, 27, 667–670.
- Fetka, I.; Radeghieri, A.; Bouwmeester, T. Expression of the RNA recognition motif-containing protein SEB-4 during Xenopus embryonic development. Mech. Dev. 2000, 94, 283–286.
- Grifone, R.; Xie, X.; Bourgeois, A.; Saquet, A.; Duprez, D.; Shi, D.L. The RNA-binding protein Rbm24 is transiently expressed in myoblasts and is required for myogenic differentiation during vertebrate development. Mech. Dev. 2014, 134, 1–15.
- Maragh, S.; Miller, R.A.; Bessling, S.L.; McGaughey, D.M.; Wessels, M.W.; de Graaf, B.; Stone, E.A.; Bertoli-Avella, A.M.; Gearhart, J.D.; Fisher, S.; et al. Identification of RNA binding motif proteins essential for cardiovascular development. BMC Dev. Biol. 2011, 11, 62.
- Miller, R.A.; Christoforou, N.; Pevsner, J.; McCallion, A.S.; Gearhart, J.D. Efficient array-based identification of novel cardiac genes through differentiation of mouse ESCs. PLoS ONE. 2008, 3, e2176.
- Poon, K.L.; Tan, K.T.; Wei, Y.Y.; Ng, C.P.; Colman, A.; Korzh, V.; Xu, X.Q. RNA-binding protein RBM24 is required for sarcomere assembly and heart contractility. Cardiovasc. Res. 2012, 94, 418–427.
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; De Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130.
- Shao, M.; Lu, T.; Zhang, C.; Zhang, Y.Z.; Kong, S.H.; Shi, D.L. Rbm24 controls poly(A) tail length and translation efficiency of crystallin mRNAs in the lens via cytoplasmic polyadenylation. Proc. Natl. Acad. Sci. USA 2020, 117, 7245–7254.
- Yang, J.; Hung, L.H.; Licht, T.; Kostin, S.; Looso, M.; Khrameeva, E.E.; Bindereif, A.; Schneider, A.; Braun, T. RBM24 Is a major regulator of muscle-specific alternative splicing. Dev. Cell. 2014, 31, 87–99.
- Liu, J.; Kong, X.; Zhang, M.; Yang, X.; Xu, X.Q. RNA binding protein 24 deletion disrupts global alternative splicing and causes dilated cardiomyopathy. Protein Cell. 2019, 10, 405–416.
- Lu, S.H.; Lee, K.Z.; Hsu, P.W.; Su, L.Y.; Yeh, Y.C.; Pan, C.Y.; Tsai, S.Y. Alternative splicing mediated by RNA-binding protein RBM24 facilitates cardiac myofibrillogenesis in a differentiation stage-specific manner. Circ. Res. 2022, 130, 112–129.
- Liu, J.; Wang, K.; Liu, X.; Pan, L.; Zhou, W.; Huang, J.; Liu, H.; Su, Z.; Xu, X.Q. RBM24 controls cardiac QT interval through CaMKIIδ splicing. Cell. Mol. Life Sci. 2022, 79, 613.
- Grifone, R.; Saquet, A.; Desgres, M.; Sangiorgi, C.; Gargano, C.; Li, Z.; Coletti, D.; Shi, D.L. Rbm24 displays dynamic functions required for myogenic differentiation during muscle regeneration. Sci. Rep. 2021, 11, 9423.
- Akerberg, A.A.; Burns, C.E.; Burns, C.G. Exploring the activities of RBPMS proteins in myocardial biology. Pediatr. Cardiol. 2019, 40, 1410–1418.
- Soufari, H.; Mackereth, C.D. Conserved binding of GCAC motifs by MEC-8, couch potato, and the RBPMS protein family. RNA. 2017, 23, 308–316.
- Gan, P.; Wang, Z.; Morales, M.G.; Zhang, Y.; Bassel-Duby, R.; Liu, N.; Olson, E.N. RBPMS is an RNA-binding protein that mediates cardiomyocyte binucleation and cardiovascular development. Dev. Cell. 2022, 57, 959–973.e7.
- Gan, P.; Wang, Z.; Bezprozvannaya, S.; McAnally, J.R.; Tan, W.; Li, H.; Bassel-Duby, R.; Liu, N.; Olson, E.N. RBPMS regulates cardiomyocyte contraction and cardiac function through RNA alternative splicing. Cardiovasc. Res. 2024, 120, 56–68.
- Akerberg, A.A.; Trembley, M.; Butty, V.; Schwertner, A.; Zhao, L.; Beerens, M.; Liu, X.; Mahamdeh, M.; Yuan, S.; Boyer, L.; et al. RBPMS2 Is a myocardial-enriched splicing regulator required for cardiac function. Circ. Res. 2022, 131, 980–1000.
- Tudurachi, B.S.; Zăvoi, A.; Leonte, A.; Țăpoi, L.; Ureche, C.; Bîrgoan, S.G.; Chiuariu, T.; Anghel, L.; Radu, R.; Sascău, R.A.; et al. An update on MYBPC3 gene mutation in hypertrophic cardiomyopathy. Int. J. Mol. Sci. 2023, 24, 10510.
- Auxerre-Plantié, E.; Nielsen, T.; Grunert, M.; Olejniczak, O.; Perrot, A.; Özcelik, C.; Harries, D.; Matinmehr, F.; Dos Remedios, C.; Mühlfeld, C.; et al. Identification of MYOM2 as a candidate gene in hypertrophic cardiomyopathy and Tetralogy of Fallot, and its functional evaluation in the Drosophila heart. Dis. Model Mech. 2020, 13, dmm045377.
- Conboy, J.G. Developmental regulation of RNA processing by Rbfox proteins. Wiley Interdiscip. Rev. RNA 2017, 8, e1398.
- Gao, C.; Ren, S.; Lee, J.H.; Qiu, J.; Chapski, D.J.; Rau, C.D.; Zhou, Y.; Abdellatif, M.; Nakano, A.; Vondriska, T.M.; et al. RBFox1-mediated RNA splicing regulates cardiac hypertrophy and heart failure. J. Clin. Investig. 2016, 126, 195–206.
- Huang, J.; Lee, J.Z.; Rau, C.D.; Pezhouman, A.; Yokota, T.; Miwa, H.; Feldman, M.; Kong, T.K.; Yang, Z.; Tay, W.T.; et al. Regulation of postnatal cardiomyocyte maturation by an RNA splicing regulator RBFox1. Circulation 2023, 148, 1263–1266.
- Nutter, C.A.; Jaworski, E.A.; Verma, S.K.; Deshmukh, V.; Wang, Q.; Botvinnik, O.B.; Lozano, M.J.; Abass, I.J.; Ijaz, T.; Brasier, A.R.; et al. Dysregulation of RBFOX2 Is an early event in cardiac pathogenesis of diabetes. Cell Rep. 2016, 15, 2200–2213.
- Damianov, A.; Black, D.L. Autoregulation of Fox protein expression to produce dominant negative splicing factors. RNA 2010, 16, 405–416.
- Glessner, J.T.; Bick, A.G.; Ito, K.; Homsy, J.; Rodriguez-Murillo, L.; Fromer, M.; Mazaika, E.; Vardarajan, B.; Italia, M.; Leipzig, J.; et al. Increased frequency of de novo copy number variants in congenital heart disease by integrative analysis of single nucleotide polymorphism array and exome sequence data. Circ. Res. 2014, 115, 884–896.
- Homsy, J.; Zaidi, S.; Shen, Y.; Ware, J.S.; Samocha, K.E.; Karczewski, K.J.; DePalma, S.R.; McKean, D.; Wakimoto, H.; Gorham, J.; et al. De novo mutations in congenital heart disease with neurodevelopmental and other congenital anomalies. Science 2015, 350, 1262–1266.
- McKean, D.M.; Homsy, J.; Wakimoto, H.; Patel, N.; Gorham, J.; DePalma, S.R.; Ware, J.S.; Zaidi, S.; Ma, W.; Patel, N.; et al. Loss of RNA expression and allele-specific expression associated with congenital heart disease. Nat. Commun. 2016, 7, 12824.
- Verma, S.K.; Deshmukh, V.; Nutter, C.A.; Jaworski, E.; Jin, W.; Wadhwa, L.; Abata, J.; Ricci, M.; Lincoln, J.; Martin, J.F.; et al. Rbfox2 function in RNA metabolism is impaired in hypoplastic left heart syndrome patient hearts. Sci. Rep. 2016, 6, 30896.
- Ying, Y.; Wang, X.J.; Vuong, C.K.; Lin, C.H.; Damianov, A.; Black, D.L. Splicing activation by Rbfox requires self-aggregation through its tyrosine-rich domain. Cell 2017, 170, 312–323.e10.
- Wei, C.; Qiu, J.; Zhou, Y.; Xue, Y.; Hu, J.; Ouyang, K.; Banerjee, I.; Zhang, C.; Chen, B.; Li, H.; et al. Repression of the central splicing regulator RBFox2 is functionally linked to pressure overload-induced heart failure. Cell Rep. 2015, 10, 1521–1533.
- Verma, S.K.; Deshmukh, V.; Thatcher, K.; Belanger, K.K.; Rhyner, A.M.; Meng, S.; Holcomb, R.J.; Bressan, M.; Martin, J.F.; Cooke, J.P.; et al. RBFOX2 is required for establishing RNA regulatory networks essential for heart development. Nucleic Acids Res. 2022, 50, 2270–2286.
- Frese, K.S.; Meder, B.; Keller, A.; Just, S.; Haas, J.; Vogel, B.; Fischer, S.; Backes, C.; Matzas, M.; Köhler, D.; et al. RNA splicing regulated by RBFOX1 is essential for cardiac function in zebrafish. J. Cell Sci. 2015, 128, 3030–3040.
- Huang, M.; Akerberg, A.A.; Zhang, X.; Yoon, H.; Joshi, S.; Hallinan, C.; Nguyen, C.; Pu, W.T.; Haigis, M.C.; Burns, C.G.; et al. Intrinsic myocardial defects underlie an Rbfox-deficient zebrafish model of hypoplastic left heart syndrome. Nat. Commun. 2022, 13, 5877.
- Jiang, X.; Liu, B.; Nie, Z.; Duan, L.; Xiong, Q.; Jin, Z.; Yang, C.; Chen, Y. The role of m6A modification in the biological functions and diseases. Signal Transduct. Target. Ther. 2021, 6, 74.
- Wang, S.; Lv, W.; Li, T.; Zhang, S.; Wang, H.; Li, X.; Wang, L.; Ma, D.; Zang, Y.; Shen, J.; et al. Dynamic regulation and functions of mRNA m6A modification. Cancer Cell Int. 2022, 22, 48.
- Sikorski, V.; Vento, A.; Kankuri, E.; IHD-EPITRAN Consortium. Emerging roles of the RNA modifications N6-methyladenosine and adenosine-to-inosine in cardiovascular diseases. Mol. Ther. Nucleic Acids 2022, 29, 426–461.
- Krumbein, M.; Oberman, F.; Cinnamon, Y.; Golomb, M.; May, D.; Vainer, G.; Belzer, V.; Meir, K.; Fridman, I.; Haybaeck, J.; et al. RNA binding protein IGF2BP2 expression is induced by stress in the heart and mediates dilated cardiomyopathy. Commun. Biol. 2023, 6, 1229.
- Gao, S.; Sun, H.; Chen, K.; Gu, X.; Chen, H.; Jiang, L.; Chen, L.; Zhang, S.; Liu, Y.; Shi, D.; et al. Depletion of m(6) A reader protein YTHDC1 induces dilated cardiomyopathy by abnormal splicing of Titin. J. Cell Mol. Med. 2021, 25, 10879–10891.
- Wang, S.; Zhang, J.; Wu, X.; Lin, X.; Liu, X.M.; Zhou, J. Differential roles of YTHDF1 and YTHDF3 in embryonic stem cell-derived cardiomyocyte differentiation. RNA Biol. 2021, 18, 1354–1363.
- Xu, H.; Wang, Z.; Chen, M.; Zhao, W.; Tao, T.; Ma, L.; Ni, Y.; Li, W. YTHDF2 alleviates cardiac hypertrophy via regulating Myh7 mRNA decoy. Cell Biosci. 2021, 11, 132.
- Golubeva, V.A.; Dorn, L.E.; Gilbert, C.J.; Rabolli, C.P.; Das, A.S.; Wanasinghe, V.S.; Veress, R.; Terentyev, D.; Accornero, F. Loss of YTHDF2 alters the expression of m6A-modified Myzap and causes adverse cardiac remodeling. JACC Basic Transl. Sci. 2023, 8, 1180–1194.
- Kmietczyk, V.; Oelschläger, J.; Gupta, P.; Varma, E.; Hartl, S.; Furkel, J.; Konstandin, M.; Marx, A.; Loewenthal, Z.; Kamuf-Schenk, V.; et al. Ythdf2 regulates cardiac remodeling through its mRNA target transcripts. J. Mol. Cell. Cardiol. 2023, 181, 57–66.
- Martino, F.; Varadarajan, N.M.; Perestrelo, A.R.; Hejret, V.; Durikova, H.; Vukic, D.; Horvath, V.; Cavalieri, F.; Caruso, F.; Albihlal, W.S.; et al. The mechanical regulation of RNA binding protein hnRNPC in the failing heart. Sci. Transl. Med. 2022, 14, eabo5715.
More
Information
Subjects:
Physiology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
777
Revisions:
2 times
(View History)
Update Date:
22 Mar 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No