 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Mostafa Ahmed Abdalmageed | -- | 8050 | 2024-03-08 16:41:36 | | | |
| 2 | Jason Zhu | -103 word(s) | 7923 | 2024-03-12 03:38:21 | | |
Video Upload Options
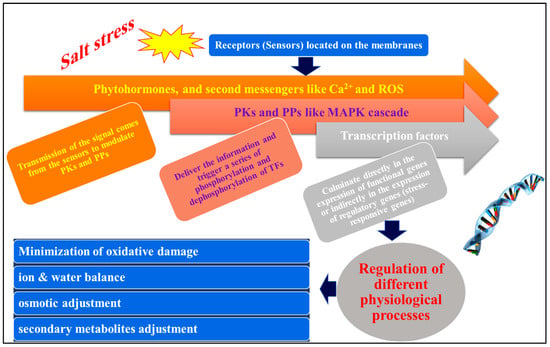
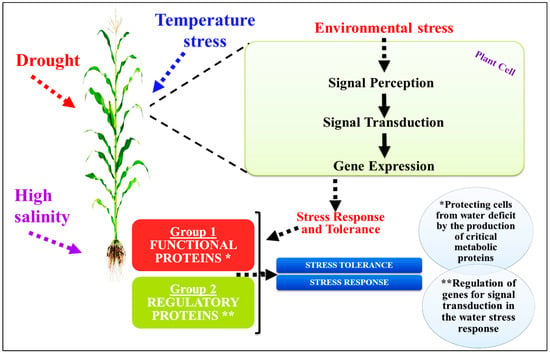
One of the most significant environmental challenges to crop growth and yield worldwide is soil salinization. Salinity lowers soil solution water potential, causes ionic disequilibrium and specific ion effects, and increases reactive oxygen species (ROS) buildup, causing several physiological and biochemical issues in plants. Plants have developed biological and molecular methods to combat salt stress. Salt-signaling mechanisms regulated by phytohormones may provide additional defense in salty conditions. That discovery helped identify the molecular pathways that underlie zinc-oxide nanoparticle (ZnO-NP)-based salt tolerance in certain plants. It emphasized the need to study processes like transcriptional regulation that govern plants’ many physiological responses to such harsh conditions. ZnO-NPs have shown the capability to reduce salinity stress by working with transcription factors (TFs) like AP2/EREBP, WRKYs, NACs, and bZIPs that are released or triggered to stimulate plant cell osmotic pressure-regulating hormones and chemicals. In addition, ZnO-NPs have been shown to reduce the expression of stress markers such as malondialdehyde (MDA) and hydrogen peroxide (H2O2) while also affecting transcriptional factors. Those systems helped maintain protein integrity, selective permeability, photosynthesis, and other physiological processes in salt-stressed plants.
1. Introduction
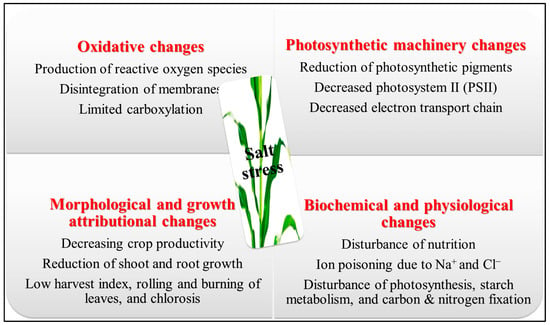
2. The Presence of High Salinity Levels Has Been Observed to Result in a Decrease in Biomass Production and Subsequent Yield Losses
3. The Negative Effects of Soil Salinity on Crops
3.1. The Effects of Salinity on the Agricultural Value Measuring Properties of Plant Species
3.2. Physiological Processes Whose Characteristics Are Influenced by Salinity Stress
3.3. The Variant Effects of Salinity on Enzymatic and Non-Enzymatic Antioxidants in Plants
4. Different Pathways of Salt Tolerance in Plants
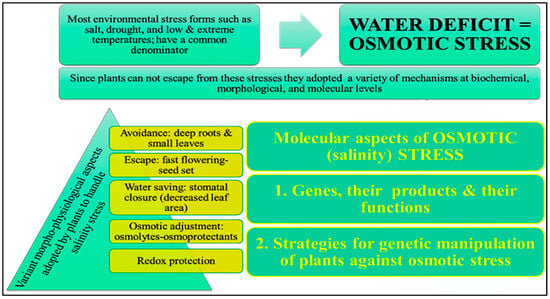
4.1. Osmotic Adjustment
4.2. The Exclusion, Redistribution, or Inclusion/Sequestration of Salt Facing the Ion Toxicity
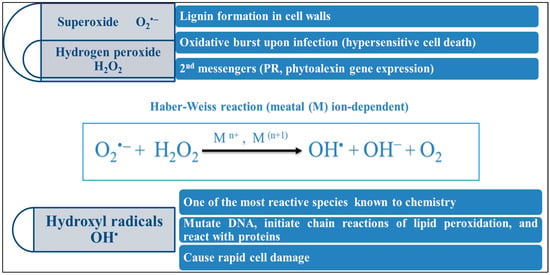
4.3. The Activation of Redox Responses
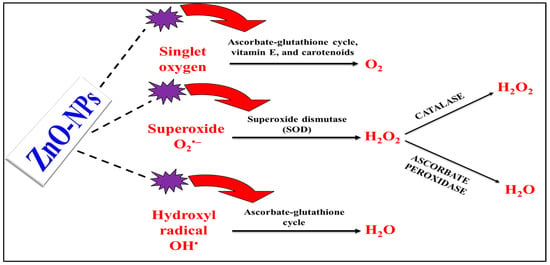
5. Exploring Nanoparticle Kinds and Their Ability to Reduce Abiotic Stress
6. Various Nanoparticles and Their Impact on the Genes Responsible for Plant Salt Tolerance: Enzymatic Expression
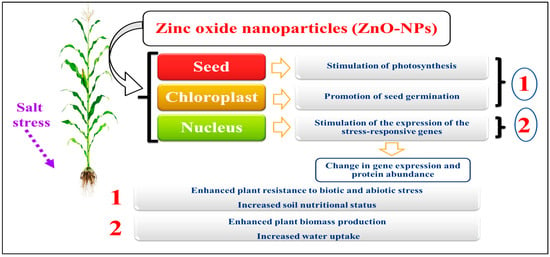
7. Transcriptional Factors Affecting Salt Tolerance

7.1. AP2/EREBP (Apetala2/Ethylene-Responsive Element-Binding Protein) Transcription Factors
7.2. WRKY TFs
7.3. NAC TFs
7.4. bZIP TFs
8. Gene Expression and Zinc Oxide Nanoparticles
9. Conclusions
References
- Ingrao, C.; Strippoli, R.; Lagioia, G.; Huisingh, D. Water Scarcity in Agriculture: An Overview of Causes, Impacts and Approaches for Reducing the Risks. Heliyon 2023, 9, e18507.
- Shrivastava, P.; Kumar, R. Soil Salinity: A Serious Environmental Issue and Plant Growth Promoting Bacteria as One of the Tools for Its Alleviation. Saudi J. Biol. Sci. 2015, 22, 123–131.
- Grigore, M.N.; Toma, C.; Grigore, M.N.; Toma, C. Saline environments; Springer International Publishing: New York, NY, USA, 2017.
- Negacz, K.; Malek, Ž.; de Vos, A.; Vellinga, P. Saline Soils Worldwide: Identifying the Most Promising Areas for Saline Agriculture. J. Arid. Environ. 2022, 203, 104775.
- Nachshon, U. Cropland Soil Salinization and Associated Hydrology: Trends, Processes and Examples. Water 2018, 10, 1030.
- Butcher, K.; Wick, A.; DeSutter, T.; Chatterjee, A.; Harmon, J. Soil Salinity: A Threat to Global Food Security. Agron. J. 2016, 108, 2189–2200.
- Nabati, J.; Kafi, M.; Nezami, A.; Moghaddam, P.R.; Ali, M.; Mehrjerdi, M.Z. Effect of Salinity on Biomass Production and Activities of Some Key Enzymatic Antioxidants in Kochia (Kochia Scoparia). Pak. J. Bot. 2011, 43, 539–548.
- Ahanger, M.A.; Agarwal, R.M.; Tomar, N.S.; Shrivastava, M. Potassium Induces Positive Changes in Nitrogen Metabolism and Antioxidant System of Oat (Avena sativa L. Cultivar Kent). J. Plant Interact. 2015, 10, 211–223.
- Munns, R.; Tester, M. Mechanisms of Salinity Tolerance. Annu. Rev. Plant Biol. 2008, 59, 651–681.
- Jiang, C.; Zu, C.; Lu, D.; Zheng, Q.; Shen, J.; Wang, H.; Li, D. Effect of Exogenous Selenium Supply on Photosynthesis, Na+ Accumulation and Antioxidative Capacity of Maize (Zea mays L.) under Salinity Stress. Sci. Rep. 2017, 7, 42039.
- Mittal, S.; Kumari, N.; Sharma, V. Differential Response of Salt Stress on Brassica Juncea: Photosynthetic Performance, Pigment, Proline, D1 and Antioxidant Enzymes. Plant Physiol. Biochem. 2012, 54, 17–26.
- Gengmao, Z.; Shihui, L.; Xing, S.; Yizhou, W.; Zipan, C. The Role of Silicon in Physiology of the Medicinal Plant (Lonicera japonica L.) under Salt Stress. Sci. Rep. 2015, 5, 12696.
- Quintero, J.M.; Fournier, J.M.; Benlloch, M. Na+ Accumulation in Shoot Is Related to Water Transport in K+-Starved Sunflower Plants but Not in Plants with a Normal K+ Status. J. Plant Physiol. 2007, 164, 60–67.
- Abbasi, G.H.; Akhtar, J.; Ahmad, R.; Jamil, M.; Anwar-Ul-Haq, M.; Ali, S.; Ijaz, M. Potassium Application Mitigates Salt Stress Differentially at Different Growth Stages in Tolerant and Sensitive Maize Hybrids. Plant Growth Regul. 2015, 76, 111–125.
- Noctor, G.; Foyer, C.H. Ascorbate and Glutathione: Keeping Active Oxygen Under Control. Ann. Rev. Plant Physiol. Plant Mol. Biol. 1998, 49, 249–279.
- Pitzschke, A.; Forzani, C.; Hirt, H. Reactive Oxygen Species Signaling in Plants. Antioxid. Redox Signal 2006, 8, 1757–1764.
- Maas, E.V. Crop Salt Tolerance. In Agricultural salinity assessment and management, ASCE Manuals and Reports on Engineering Practice; Tanji, K.K., Ed.; American Society of Civil Engineers: Reston, VA, USA, 1990.
- Banerjee, A.; Roychoudhury, A. Abscisic-acid-dependent basic leucine zipper (bZIP) transcription factors in plant abiotic stress. Protoplasma 2017, 254, 3–16.
- Joshi, R.; Wani, S.H.; Singh, B. Transcription Factors and Plants Response to Drought Stress: Current Understanding and Future Directions. Front. Plant Sci. 2016, 7, 1029.
- Golldack, D.; Li, C.; Mohan, H.; Probst, N. Tolerance to Drought and Salt Stress in Plants: Unraveling the Signaling Networks. Front. Plant Sci. 2014, 5, 151.
- Handford, C.E.; Dean, M.; Henchion, M.; Spence, M.; Elliott, C.T.; Campbell, K. Implications of Nanotechnology for the Agri-Food Industry: Opportunities, Benefits and Risks. Trends Food Sci. Technol. 2014, 40, 226–241.
- Acharya, A.; Pal, P.K. Agriculture Nanotechnology: Translating Research Outcome to Field Applications by Influencing Environmental Sustainability. NanoImpact 2020, 19, 100232.
- Foley, J.A.; Ramankutty, N.; Brauman, K.A.; Cassidy, E.S.; Gerber, J.S.; Johnston, M. Solutions for a Cultivated Planet. Nature 2011, 478, 337–342.
- Tuomisto, H.L.; Scheelbeek, P.F.D.; Chalabi, Z.; Green, R.; Smith, R.D.; Haines, A.; Dangour, A.D. Effects of Environmental Change on Agriculture, Nutrition and Health: A Framework with a Focus on Fruits and Vegetables. Wellcome Open Res. 2017, 2, 21.
- Raza, A.; Razzaq, A.; Mehmood, S.S.; Zou, X.; Zhang, X.; Lv, Y.; Xu, J. Impact of Climate Change on Crops Adaptation and Strategies to Tackle Its Outcome: A Review. Plants 2019, 8, 34.
- Dresselhaus, T.; Hückelhoven, R. Biotic and Abiotic Stress Responses in Crop Plants. Agronomy 2018, 8, 267.
- Sekhon, B.S. Nanotechnology in Agri-Food Production: An Overview. Nanotechnol. Sci. Appl. 2014, 7, 31–53.
- Tipu, M.M.H.; Baroi, A.; Rana, J.; Islam, S.; Jahan, R.; Miah, M.S. Potential Applications of Nanotechnology in Agriculture: A Smart Tool for Sustainable Agriculture. In Agricultural Development in Asia-Potential Use of Nano-Materials and Nano-Technology; IntechOpen: London, UK, 2021.
- Kumar, A.A.; Mishra, P.; Kumari, K.; Panigrahi, K. Environmental Stress Influencing Plant Development and Flowering. Front. Biosci. 2012, 4, 1315–1324.
- Yang, L.; Wen, K.-S.; Ruan, X.; Zhao, Y.-X.; Wei, F.; Wang, Q. Response of plant secondary metabolites to environmental factors. Molecules 2018, 23, 762.
- Das, K.; Roychoudhury, A. Reactive Oxygen Species (ROS) and Response of Antioxidants as ROS-Scavengers during Environmental Stress in Plants. Front. Environ. Sci. 2014, 2, 53.
- Rai, P.K.; Kumar, V.; Lee, S.; Raza, N.; Kim, K.-H.; Ok, Y.S. Nanoparticleplant Interaction: Implications in Energy, Environment, and Agriculture. Environ. Int. 2018, 119, 1–19.
- Abdel Latef, A.A.H.; Abu Alhmad, M.F.; Abdelfattah, K.E. The Possible Roles of Priming with ZnO Nanoparticles in Mitigation of Salinity Stress in Lupine (Lupinus Termis. Plants. J. Plant Growth Regul. 2017, 36, 60–70.
- Taran, N.; Storozhenko, V.; Svietlova, N.; Batsmanova, L.; Shvartau, V.; Kovalenko, M. Effect of Zinc and Copper Nanoparticles on Drought Resistance of Wheat Seedlings. Nanoscale Res. Lett. 2017, 12, 60.
- Goharrizi, K.J.; Baghizadeh, A.; Kalantar, M.; Fatehi, F. Combined Effects of Salinity and Drought on Physiological and Biochemical Characteristics of Pistachio Rootstocks. Sci. Hortic. 2020, 261, 108970.
- Hussain, S.; Cao, X.; Zhong, C.; Zhu, L.; Khaskheli, M.A.; Fiaz, S.; Zhang, J.; Jin, Q. Sodium Chloride Stress during Early Growth Stages Altered Physiological and Growth Characteristics of Rice. Chil. J. Agric. Res. 2018, 78, 183–197.
- Ahmad, M.; Zahir, Z.A.; Naeem Asghar, H.; Asghar, M. Inducing Salt Tolerance in Mung Bean through Coinoculation with Rhizobia and Plant-Growth-Promoting Rhizobacteria Containing 1- Aminocyclopropane-1-Carboxylate Deaminase. Can. J. Microbiol. 2011, 57, 578–589.
- Munns, R.; James, R.A.; Läuchli, A. Approaches to Increasing the Salt Tolerance of Wheat and Other Cereals. J. Exp. Bot. 2006, 57, 1025–1043.
- Tavakkoli, E.; Rengasamy, P.; McDonald, G.K. High Concentrations of Na+ and Cl– Ions in Soil Solution Have Simultaneous Detrimental Effects on Growth of Faba Bean under Salinity Stress. J. Exp. Bot. 2010, 61, 4449–4459.
- Geilfus, C.-M.; Tenhaken, R.; Carpentier, S.C. Transient Alkalinization of the Leaf Apoplast Stiffens the Cell Wall during Onset of Chloride Salinity in Corn Leaves. J. Biol. Chem. 2017, 292, 18800–18813.
- Ma, Y.; Dias, M.C.; Freitas, H. Drought and Salinity Stress Responses and Microbe-Induced Tolerance in Plants. Front. Plant Sci. 2020, 11, 591911.
- Munns, R.; Gilliham, M. Salinity Tolerance of Crops—What Is the Cost? New Phytol. 2015, 208, 668–673.
- Amthor, J. The McCree–de Wit–Penning de Vries–Thornley Respiration Paradigms: 30 Years Later. Ann. Bot. 2000, 86, 1–20.
- Zhao, S.; Zhang, Q.; Liu, M.; Zhou, H.; Ma, C.; Wang, P. Regulation of Plant Responses to Salt Stress. Int. J. Mol. Sci. 2021, 22, 4609.
- Childs, B.G.; Durik, M.; Baker, D.J.; van Deursen, J.M. Cellular Senescence in Aging and Age-Related Disease: From Mechanisms to Therapy. Nat. Med. 2015, 21, 1424–1435.
- Werf, A.; Kooijman, A.; Welschen, R.; Lambers, H. Respiratory Energy Costs for the Maintenance of Biomass, for Growth and for Ion Uptake in Roots ofCarex diandraandCarex Acutiformis. Physiol. Plant. 1988, 72, 483–491.
- Katerji, N.; van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M. Salt Tolerance of Crops According to Three Classification Methods and Examination of Some Hypothesis about Salt Tolerance. Agric. Water Manag. 2001, 47, 1–8.
- Al-shareef, N.O.; Tester, M. Plant Salinity Tolerance. In Encyclopedia of Life Sciences; Wiley: Hoboken, NJ, USA, 2019; pp. 1–6.
- Richard, B.; Qi, A.; Fitt, B.D.L. Control of Crop Diseases through Integrated Crop Management to Deliver Climate-smart Farming Systems for Low- and High-input Crop Production. Plant Pathol. 2022, 71, 187–206.
- De-la-Cruz Chacón, I.; Riley-Saldaña, C.A.; González-Esquinca, A.R. Secondary Metabolites during Early Development in Plants. Phytochem. Rev. 2013, 12, 47–64.
- Kamran, M.; Parveen, A.; Ahmar, S.; Malik, Z.; Hussain, S.; Chattha, M.S.; Saleem, M.H.; Adil, M.; Heidari, P.; Chen, J.-T. An Overview of Hazardous Impacts of Soil Salinity in Crops, Tolerance Mechanisms, and Amelioration through Selenium Supplementation. Int. J. Mol. Sci. 2019, 21, 148.
- Munns, R. Physiological Processes Limiting Plant Growth in Saline Soils: Some Dogmas and Hypotheses. Plant Cell Environ. 1993, 16, 15–24.
- Aghighi Shahverdi, M.; Omidi, H.; Tabatabaei, S.J. Plant Growth and Steviol Glycosides as Affected by Foliar Application of Selenium, Boron, and Iron under NaCl Stress in Stevia Rebaudiana Bertoni. Ind. Crops Prod. 2018, 125, 408–415.
- Shahrajabian, M.H.; Khoshkharam, M.; Sun, W.; Cheng, Q. Germination and Seedlings Growth of Corn (Zea mays L.) to Allelopathic Effects of Rice (Oryza sativa L.). Trop. Plant Res. 2019, 6, 152–156.
- Lee, D.G.; Park, K.W.; An, J.Y.; Sohn, Y.G.; Ha, J.K.; Kim, H.Y.; Bae, D.W.; Lee, K.H.; Kang, N.J.; Lee, B.H. Proteomics Analysis of Salt-Induced Leaf Proteins in Two Rice Germplasms with Different Salt Sensitivity. Can. J. Plant Sci. 2011, 91, 337–349.
- Ibrahim, E.A. Seed Priming to Alleviate Salinity Stress in Germinating Seeds. J. Plant Physiol. 2016, 192, 38–46.
- Ashraf, M. Relationships between Growth and Gas Exchange Characteristics in Some Salt-Tolerant Amphidiploid Brassica Species in Relation to Their Diploid Parents. Environ. Exp. Bot. 2001, 45, 155–163.
- Qados, A.M.S.A. Effect of salt stress on plant growth and metabolism of bean plant Vicia faba (L.). J. Saudi Soc. Agric. Sci. 2011, 10, 7–15.
- Ahmadi, F.I.; Karimi, K.; Struik, P.C. Effect of Exogenous Application of Methyl Jasmonate on Physiological and Biochemical Characteristics of Brassica Napus L. Cv. Talaye under Salinity Stress. S. Afr. J. Bot. 2018, 115, 5–11.
- Hajiboland, R.; Norouzi, F.; Poschenrieder, C. Growth, Physiological, Biochemical and Ionic Responses of Pistachio Seedlings to Mild and High Salinity. Trees Struct. Funct. 2014, 28, 1065–1078.
- Mallahi, T.; Saharkhiz, M.J.; Javanmardi, J. Salicylic Acid Changes Morpho-Physiological Attributes of Feverfew (Tanacetum parthenium L.) under Salinity Stress. Acta Ecol. Sin. 2018, 38, 351–355.
- Shi, Q.; Ding, F.; Wang, X.; Wei, M. Exogenous Nitric Oxide Protects Cucumber Roots against Oxidative Stress Induced by Salt Stress. Plant Physiol. Biochem. 2007, 45, 542–550.
- Hameed, A.; Ahmed, M.Z.; Hussain, T.; Aziz, I.; Ahmad, N.; Gul, B.; Nielsen, B.L. Effects of Salinity Stress on Chloroplast Structure and Function. Cells 2021, 10, 2023.
- Netondo, G.W.; Onyango, J.C.; Beck, E.S.; salinity, I. Gas Exchange and Chlorophyll Fluorescence of Sorghum under Salt Stress. Crop Sci. 2004, 44, 806.
- Sapre, S.; Gontia-Mishra, I.; Tiwari, S. Klebsiella Sp. Confers Enhanced Tolerance to Salinity and Plant Growth Promotion in Oat Seedlings (Avena sativa). Microbiol. Res. 2018, 206, 25–32.
- Rozeff, N. Sugarcane and Salinity—A Review Paper. Sugar Cane 1995, 5, 8–19.
- Cantabella, D.; Piqueras, A.; Acosta-Motos, J.R.; Bernal-Vicente, A.; Hernández, J.A.; Díaz-Vivancos, P. Salt-Tolerance Mechanisms Induced in Stevia Rebaudiana Bertoni: Effects on Mineral Nutrition, Antioxidative Metabolism and Steviol Glycoside Content. Plant Physiol. Biochem. 2017, 115, 484–496.
- Elkelish, A.A.; Soliman, M.H.; Alhaithloul, H.A.; El-Esawi, M.A. Selenium Protects Wheat Seedlings against Salt Stress-Mediated Oxidative Damage by up-Regulating Antioxidants and Osmolytes Metabolism. Plant Physiol. Biochem. 2019, 137, 144–153.
- Khan, M.I.R.; Asgher, M.; Khan, N.A. Alleviation of Salt-Induced Photosynthesis and Growth Inhibition by Salicylic Acid Involves Glycinebetaine and Ethylene in Mungbean (Vigna radiata L.). Plant Physiol. Biochem. 2014, 80, 67–74.
- Munns, R. Genes and Salt Tolerance: Bringing Them Together. New Phytol. 2005, 167, 645–663.
- Hussain, S.; Zhang, J.H.; Zhong, C.; Zhu, L.F.; Cao, X.C.; YU, S.M.; Allen Bohr, J.; Hu, J.J.; Jin, Q.Y. Effects of Salt Stress on Rice Growth, Development Characteristics, and the Regulating Ways: A Review. J. Integr. Agric. 2017, 16, 2357–2374.
- Hussain, S.; Zhong, C.; Bai, Z.; Cao, X.; Zhu, L.; Hussain, A.; Zhu, C.; Fahad, S.; James, A.B.; Zhang, J. Effects of 1-Methylcyclopropene on Rice Growth Characteristics and Superior and Inferior Spikelet Development Under Salt Stress. J. Plant Growth Regul. 2018, 37, 1368–1384.
- Wang, W.; Vinocur, B.; Altman, A. Plant Responses to Drought, Salinity and Extreme Temperatures: Towards Genetic Engineering for Stress Tolerance. Planta 2003, 218, 1–14.
- Zahedi, S.M.; Abdelrahman, M.; Hosseini, M.S.; Hoveizeh, N.F.; Tran, L.P. Alleviation of the Effect of Salinity on Growth and Yield of Strawberry by Foliar Spray of Selenium-Nanoparticles. In Environmental Pollution; Elsevier: Amsterdam, The Netherlands, 2019.
- Gururani, M.A.; Venkatesh, J.; Tran, L.S.P. Regulation of Photosynthesis during Abiotic Stress-Induced Photoinhibition. Mol. Plant 2015, 8, 1304–1320.
- James, R.A.; Blake, C.; Byrt, C.S.; Munns, R. Major Genes for Na+ Exclusion, Nax1 and Nax2 (Wheat HKT1;4 and HKT1;5), Decrease Na+ Accumulation in Bread Wheat Leaves under Saline and Waterlogged Conditions. J. Exp. Bot. 2011, 62, 2939–2947.
- Evelin, H.; Kapoor, R.; Giri, B. Arbuscular Mycorrhizal Fungi in Alleviation of Salt Stress: A Review. Ann. Bot. 2009, 104, 1263–1280.
- Hussain, S.; Bai, Z.; Huang, J.; Cao, X.; Zhu, L.; Zhu, C.; Khaskheli, M.A.; Zhong, C.; Jin, Q.; Zhang, J. 1-Methylcyclopropene Modulates Physiological, Biochemical, and Antioxidant Responses of Rice to Different Salt Stress Levels. Front. Plant Sci. 2019, 10, 124.
- Singam, K.; Juntawong, N.; Cha-Um, S.; Kirdmanee, C. Salt Stress Induced Ion Accumulation, Ion Homeostasis, Membrane Injury and Sugar Contents in Salt-Sensitive Rice (Oryza sativa L. spp. Indica) Roots under Isoosmotic Conditions. Afr. J. Biotechnol. 2011, 10, 1340–1346.
- Demiral, T.; Türkan, I. Comparative Lipid Peroxidation, Antioxidant Defense Systems and Proline Content in Roots of Two Rice Cultivars Differing in Salt Tolerance. Environ. Exp. Bot. 2005, 53, 247–257.
- Levitt, J. Responses of Plants to Environmental Stresses. J. Range Manag. 1985, 38, 480.
- Pattanayak, G.K.; Tripathy, B.C. Overexpression of Protochlorophyllide Oxidoreductase c Regulates Oxidative Stress in Arabidopsis. PLoS ONE 2011, 6, 26532.
- Santos, M.G.; Ribeiro, R.V.; Machado, E.C.; Pimentel, C. Photosynthetic Parameters and Leaf Water Potential of Five Common Bean Genotypes under Mild Water Deficit. Biol. Plant 2009, 53, 229–236.
- Subramanyam, K.; Laing, G.; Damme, E.J.M. Sodium Selenate Treatment Using a Combination of Seed Priming and Foliar Spray Alleviates Salinity Stress in Rice. Front. Plant Sci. 2019, 10, 116.
- Foyer, C.H.; Noctor, G. Redox Sensing and Signalling Associated with Reactive Oxygen in Chloroplasts, Peroxisomes and Mitochondria. Physiol. Plant 2003, 119, 355–364.
- Kamran, M.; Malik, Z.; Parveen, A.; Huang, L.; Riaz, M.; Bashir, S.; Mustafa, A.; Abbasi, G.H.; Xue, B.; Ali, U. Ameliorative Effects of Biochar on Rapeseed (Brassica napus L.) Growth and Heavy Metal Immobilization in Soil Irrigated with Untreated Wastewater. J. Plant Growth Regul. 2020, 39, 266–281.
- Adly, A.A.M. Oxidative stress and disease: An updated review. Res. J. Immunol. 2010, 3, 129–145.
- Houot, V.; Etienne, P.; Petitot, A.S.; Barbier, S.; Blein, J.P.; Suty, L. Hydrogen peroxide induces programmed cell death features in cultured tobacco BY-2 cells, in a dose-dependent manner. J. Exp. Bot. 2001, 52, 1721–1730.
- Farmer, E.E.; Mueller, M.J. ROS-Mediated Lipid Peroxidation and RES-Activated Signaling. Ann. Rev. Plant Biol. 2013, 64, 429–450.
- Asada, K. Causes of Photooxidative Stress and Amelioration of Defense Systems in Plants. In Production and Action of Active Oxygen Species in Photosynthetic Tissues; CRC Press: Boca Raton, FL, USA, 1994; pp. 77–104.
- Ramachandra Reddy, A.; Chaitanya, K.V.; Jutur, P.P.; Sumithra, K. Differential Antioxidative Responses to Water Stress among Five Mulberry (Morus alba L.) Cultivars. Environ. Exp. Bot. 2004, 52, 33–42.
- Li, M.; Guo, S.; Xu, Y.; Meng, Q.; Li, G.; Yang, X. Glycine Betaine-Mediated Potentiation of HSP Gene Expression Involves Calcium Signaling Pathways in Tobacco Exposed to NaCl Stress. Physiol. Plant 2014, 150, 63–75.
- Farhangi-Abriz, S.; Ghassemi-Golezani, K. How Can Salicylic Acid and Jasmonic Acid Mitigate Salt Toxicity in Soybean Plants? Ecotoxicol. Environ. Saf. 2018, 154, 1010–1016.
- Krishnaiah, D.; Sarbatly, R.; Nithyanandam, R. A Review of the Antioxidant Potential of Medicinal Plant Species. Food Bioprod. Process 2011, 89, 217–233.
- Buchanan, B.B.; Gruissem, W.; Jones, R.L. (Eds.) Biochemistry and Molecular Biology of Plants, 2nd ed.; Wiley-Blackwell:: Hoboken, NJ, USA, 2015.
- Ashraf, M.; Foolad, M.R. Roles of Glycine Betaine and Proline in Improving Plant Abiotic Stress Resistance. Environ. Exp. Bot. 2007, 59, 206–216.
- Mansour, M.M.F.; Ali, E.F. Evaluation of Proline Functions in Saline Conditions. Phytochemistry 2017, 140, 52–68.
- Balasubramaniam, T.; Shen, G.; Esmaeili, N.; Zhang, H. Plants’ Response Mechanisms to Salinity Stress. Plants 2023, 12, 2253.
- Zhu, J.K.; Liu, J.P.; Xiong, L.M. Genetic Analysis of Salt Tolerance in Arabidopsis: Evidence for a Critical Role of Potassium Nutrition. Plant Cell 1998, 10, 1181–1191.
- Hauser, F.; Horie, T. A Conserved Primary Salt Tolerance Mechanism Mediated by HKT Transporters: A Mechanism for Sodium Exclusion and Maintenance of High K/Na Ratio in Leaves during Salinity Stress. Plant Cell Environ. 2010, 33, 552–565.
- Galva, C.; Artigas, P.; Gatto, C. Nuclear Na+/K+-ATPase Plays an Active Role in Nucleoplasmic Ca2+ Homeostasis. J. Cell Sci. 2012, 125, 6137–6147.
- Lunde, C.; Drew, D.P.; Jacobs, A.K.; Tester, M. Exclusion of Na+ via Sodium ATPase (PpENA1) Ensures Normal Growth of Physcomitrella Patens under Moderate Salt Stress. Plant 2007, 144, 1786–1796.
- Wu, H.; Shabala, L.; Azzarello, E.; Huang, Y.; Pandolfi, C.; Su, N.; Wu, Q.; Cai, S.; Bazihizina, N.; Wang, L.; et al. Na+ Extrusion from the Cytosol and Tissue-Specific Na+ Sequestration in Roots Confer Differential Salt Stress Tolerance between Durum and Bread Wheat. J. Exp. Bot. 2018, 69, 3987–4001.
- Cuin, T.A.; Bose, J.; Stefano, G.; Jha, D.; Tester, M.; Mancuso, S.; Shabala, S. Assessing the Role of Root Plasma Membrane and Tonoplast Na+/H+ Exchangers in Salinity Tolerance in Wheat: In Planta Quantification Methods: Cytosolic Na+ Exclusion in Wheat. Plant Cell Environ. 2011, 34, 947–961.
- Byrt, C.S.; Platten, J.D.; Spielmeyer, W.; James, R.A.; Lagudah, E.S.; Dennis, E.S.; Tester, M.; Munns, R. HKT1;5-like Cation Transporters Linked to Na+ Exclusion Loci in Wheat, Nax2 and Kna1. Plant Physiol. 2007, 143, 1918–1928.
- Munns, R.; James, R.A.; Xu, B. Wheat Grain Yield on Saline Soils Is Improved by an Ancestral Na+ Transporter Gene. Nature 2012, 30, 360–364.
- Jaime-Pérez, N.; Pineda, B.; García-Sogo, B.; Atares, A.; Athman, A.; Byrt, C.S.; Olías, R.; Asins, M.J.; Gilliham, M.; Moreno, V.; et al. The Sodium Transporter Encoded by the HKT1;2 Gene Modulates Sodium/Potassium Homeostasis in Tomato Shoots under Salinity. Plant Cell Environ. 2017, 40, 658–671.
- Hsu, S.Y.; Kao, C.H. Differential Effect of Sorbitol and Polyethylene Glycol on Antioxidant Enzymes in Rice Leaves. Plant Growth Regul. 2003, 39, 83–90.
- Ghannoum, O. C4 Photosynthesis and Water Stress. Ann. Bot. 2009, 103, 635–644.
- Azeem, M.; Pirjan, K.; Qasim, M. Salinity stress improves antioxidant potential by modulating physio-biochemical responses in Moringa oleifera Lam. Sci. Rep. 2023, 13, 2895.
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of Nanoparticle Toxicity on Their Physical and Chemical Properties. Nanoscale Res. Lett. 2018, 13, 1–21.
- Dziergowska, K.; Michalak, I. Chapter 9—The Role of Nanoparticles in Sustainable Agriculture. In Smart Agrochemicals for Sustainable Agriculture; Chojnacka, K., Saeid, A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 225–278. ISBN 978-0-12-817036-6.
- Solaiman, M.A.; Ali, M.A.; Abdel-Moein, N.M.; Mahmoud, E.A. Synthesis of Ag-NPs Developed by Green-Chemically Method and Evaluation of Antioxidant Activities and Anti-Inflammatory of Synthesized Nanoparticles against LPS-Induced NO in RAW 264.7 Macrophages. Biocatal. Agric. Biotechnol. 2020, 29, 101832.
- Rastogi, A.; Zivcak, M.; Sytar, O.; Kalaji, H.M.; He, X.; Mbarki, S.; Brestic, M. Impact of Metal and Metal Oxide Nanoparticles on Plant: A Critical Review. Front. Chem. 2017, 5, 78.
- Ambrosone, A.; Mattera, L.; Marchesano, V.; Quarta, A.; Susha, A.S.; Tino, A.; Rogach, A.L.; Tortiglione, C. Mechanisms Underlying Toxicity Induced by CdTe Quantum Dots Determined in an Invertebrate Model Organism. Biomaterials 2012, 33, 1991–2000.
- Nguyen, K.C.; Rippstein, P.; Tayabali, A.F.; Willmore, W.G. Mitochondrial Toxicity of Cadmium Telluride Quantum Dot Nanoparticles in Mammalian Hepatocytes. Toxicol. Sci. 2015, 146, 31–42.
- Singh, B.R.; Singh, B.N.; Khan, W.; Singh, H.B.; Naqvi, A.H. ROS-Mediated Apoptotic Cell Death in Prostate Cancer LNCaP Cells Induced by Biosurfactant Stabilized CdS Quantum Dots. Biomaterials 2012, 33, 5753–5767.
- Paramo, L.A.; Feregrino-Pérez, A.A.; Guevara, R.; Mendoza, S.; Esquivel, K. Nanoparticles in Agroindustry: Applications, Toxicity, Challenges, and Trends. Nanomaterials 2020, 10, 1654.
- Alabdallah, N.M.; Alzahrani, H.S. The Potential Mitigation Effect of ZnO Nanoparticles on Metabolism under Salt Stress Conditions. Saudi J. Biol. Sci. 2020, 27, 3132–3137.
- Sarraf, M.; Vishwakarma, K.; Kumar, V.; Arif, N.; Das, S.; Johnson, R. Metal/Metalloid-Based Nanomaterials for Plant Abiotic Stress Tolerance: An Overview of the Mechanisms. Plants 2022, 11, 316.
- Khattak, S.G.; Dominy, P.J.; Ahmad, W. Effect of Zn as Soil Addition and Foliar Application on Yield and Protein Content of Wheat in Alkaline Soil. J. Natl. Sci. Found. Sri Lanka 2015, 43, 303.
- Ghiyasi, M.; Rezaee Danesh, Y.; Amirnia, R.; Najafi, S.; Mulet, J.M.; Porcel, R. Foliar Applications of ZnO and Its Nanoparticles Increase Safflower (Carthamus tinctorius L.) Growth and Yield under Water Stress. Agronomy 2023, 13, 192.
- Aljutheri, H.; Habeeb, K.; Al-Taey, D.; Rahman, A.; Al Tawaha, A.R.; Kadhim, F. Effect of Foliar Application of Different Sources of Nano-Fertilizers on Growth and Yield of wheatEffect of Foliar Application of Different Sources of Nano-Fertilizers on Growth and Yield of Wheat. Biosci. Res. 2018, 4, 3976–3985.
- Rossi, L.; Fedenia, L.N.; Sharifan, H.; Ma, X.; Lombardini, L. Effects of Foliar Application of Zinc Sulfate and Zinc Nanoparticles in Coffee (Coffea arabica L.) Plants. Plant Physiol. Biochem. 2019, 135, 160–166.
- Rajput, V.D.; Minkina, T.M.; Behal, A.; Sushkova, S.N.; Mandzhieva, S.; Singh, R.; Gorovtsov, A.; Tsitsuashvili, V.S.; Purvis, W.O.; Ghazaryan, K.A.; et al. Effects of Zinc-Oxide Nanoparticles on Soil, Plants, Animals and Soil Organisms: A Review. Environ. Nanotechnol. Monit. Manag. 2018, 9, 76–84.
- Rao, S.; Shekhawat, G.S. Phytotoxicity and Oxidative Stress Perspective of Two Selected Nanoparticles in Brassica Juncea. 3 Biotech 2016, 6, 244.
- Zohra, E.; Ikram, M.; Omar, A.A.; Hussain, M.; Satti, S.H.; Raja, N.I.; Ehsan, M. Potential Applications of Biogenic Selenium Nanoparticles in Alleviating Biotic and Abiotic Stresses in Plants: A Comprehensive Insight on the Mechanistic Approach and Future Perspectives. Green. Process. Synth. 2021, 10, 456–475.
- Munir, N.; Hanif, M.; Dias, D.A.; Abideen, A. The Role of Halophytic Nanoparticles towards the Remediation of Degraded and Saline Agricultural Lands. Environ. Sci. Pollut. Res. 2021, 28, 60383–60405.
- Sharma, P.; Bhatt, D.; Zaidi, M.; Saradhi, P.P.; Khanna, P.; Arora, S. Silver Nanoparticle-Mediated Enhancement in Growth and Antioxidant Status of Brassica Juncea. Appl. Biochem. Biotechnol. 2012, 167, 2225–2233.
- Gunjan, B.; Zaidi, M. Impact of Gold Nanoparticles on Physiological and Biochemical Characteristics of Brassica Juncea. J. Plant Biochem. Physiol. 2014, 2, 67–73.
- Nair, P.M.G.; Chung, I.M. Assessment of silver nanoparticle-induced physiological and molecular changes in Arabidopsis thaliana. Environ. Sci. Pollut. Res. 2014, 21, 8858–8869.
- Mirzajani, F.; Askari, H.; Hamzelou, S.; Schober, Y.; Römpp, A.; Ghassempour, A.; Spengler, B. Proteomics study of silver nanoparticles toxicity on Oryza sativa L. Ecotoxicol. Environ. Saf. 2014, 108, 335–339.
- Khodakovskaya, M.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon Nanotubes Are Able to Penetrate Plant Seed Coat and Dramatically Affect Seed Germination and Plant Growth. ACS Nano 2009, 3, 3221–3227.
- Ahmed, M.; Decsi, K.; Tóth, Z. Different Tactics of Synthesized Zinc Oxide Nanoparticles, Homeostasis Ions, and Phytohormones as Regulators and Adaptatively Parameters to Alleviate the Adverse Effects of Salinity Stress on Plants. Life 2022, 13, 73.
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 Mediates Osmotic-Stress-Evoked Ca2+ Increases Vital for Osmosensing in Arabidopsis. Nature 2014, 514, 367–371.
- Khan, S.-A.; Li, M.-Z.; Wang, S.-M.; Yin, H.-J. Revisiting the Role of Plant Transcription Factors in the Battle against Abiotic Stress. Int. J. Mol. Sci. 2018, 19, 1634.
- Asai, T.; Tena, G.; Plotnikova, J.; Willmann, M.R.; Chiu, W.-L.; Gomez-Gomez, L.; Boller, T.; Ausubel, F.M.; Sheen, J. MAP Kinase Signalling Cascade in Arabidopsis Innate Immunity. Nature 2002, 415, 977–983.
- Hu, W.; Huang, C.; Deng, X.; Zhou, S.; Chen, L.; Li, Y.; Wang, C.; Ma, Z.; Yuan, Q.; Wang, Y.; et al. TaASR1, a Transcription Factor Gene in Wheat, Confers Drought Stress Tolerance in Transgenic Tobacco: The Role of TaASR1 in Drought Stress. Plant Cell Environ. 2013, 36, 1449–1464.
- Kacperska, A. Sensor Types in Signal Transduction Pathways in Plant Cells Responding to Abiotic Stressors: Do They Depend on Stress Intensity? Physiol. Plant. 2004, 122, 159–168.
- Liu, W.; Tai, H.; Li, S.; Gao, W.; Zhao, M.; Xie, C.; Li, W.-X. bHLH122 Is Important for Drought and Osmotic Stress Resistance in Arabidopsis and in the Repression of ABA Catabolism. New Phytol. 2014, 201, 1192–1204.
- Ma, Y.; Dai, X.; Xu, Y.; Luo, W.; Zheng, X.; Zeng, D.; Pan, Y.; Lin, X.; Liu, H.; Zhang, D.; et al. COLD1 confers chilling tolerance in rice. Cell 2015, 160, 1209–1221.
- Saibo, N.J.M.; Lourenço, T.; Oliveira, M.M. Transcription Factors and Regulation of Photosynthetic and Related Metabolism under Environmental Stresses. Ann. Bot. 2009, 103, 609–623.
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channels. Plant Physiol. 2013, 163, 514–522.
- Torres, M.A.; Dangl, J.L. Functions of the Respiratory Burst Oxidase in Biotic Interactions, Abiotic Stress and Development. Curr. Opin. Plant Biol. 2005, 8, 397–403.
- Sharoni, A.M.; Nuruzzaman, M.; Satoh, K. Gene Structures, Classification and Expression Models of the AP2/EREBP Transcription Factor Family in Rice. Plant Cell Physiol. 2011, 52, 344–360.
- Riechmann, J.L.; Meyerowitz, E.M. The AP2/EREBP Family of Plant Transcription Factors. Biol. Chem. 1998, 379, 633–646.
- Augustine, S.M.; Ashwin Narayan, J.; Syamaladevi, D.P. Overexpression of EaDREB2 and Pyramiding of EaDREB2 with the Pea DNA Helicase Gene (PDH45) Enhance Drought and Salinity Tolerance in Sugarcane (Saccharum spp. Hybrid). Plant Cell Rep. 2015, 34, 247–263.
- Bouaziz, D.; Pirrello, J.; Charfeddine, M. Overexpression of StDREB1 Transcription Factor Increases Tolerance to Salt in Transgenic Potato Plants. Mol. Biotechnol. 2013, 54, 803–817.
- Cai, H.; Tian, S.; Dong, H.; Guo, C. Pleiotropic Effects of TaMYB3R1 on Plant Development and Response to Osmotic Stress in Transgenic Arabidopsis. Gene 2015, 558, 227–234.
- Cao, Z.H.; Zhang, S.Z.; Wang, R.K. Genome Wide Analysis of the Apple MYB Transcription Factor Family Allows the Identification of MdoMYB121 Gene Confering Abiotic Stress Tolerance in Plants. PLoS ONE 2013, 8, e69955.
- Banerjee, A.; Roychoudhury, A. WRKY Proteins: Signaling and Regulation of Expression during Abiotic Stress Responses. Sci. World J. 2015, 2015, 807560.
- Rushton, P.J.; Somssich, I.E.; Ringler, P.; Shen, Q.J. WRKY Transcription Factors. Trends Plant Sci. 2010, 15, 247–258.
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent Advances in Utilizing Transcription Factors to Improve Plant Abiotic Stress Tolerance by Transgenic Technology. Front. Plant Sci. 2016, 7, 67.
- Agarwal, P.; Dabi, M.; Sapara, K.K. Ectopic Expression of JcWRKY Transcription Factor Confers Salinity Tolerance via Salicylic Acid Signaling. Front. Plant Sci. 2016, 7, 1541.
- Le, D.T.; Nishiyama, R.; Watanabe, Y.A. Genome-Wide Survey and Expression Analysis of the Plant-Specific NAC Transcription Factor Family in Soybean during Development and Dehydration Stress. DNA Res. 2011, 18, 263–276.
- Nuruzzaman, M.; Manimekalai, R.; Sharoni, A.M. Genome-Wide Analysis of NAC Transcription Factor Family in Rice. Gene 2010, 465, 30–44.
- Puranik, S.; Sahu, P.P.; Mandal, S.N. Comprehensive Genome-Wide Survey, Genomic Constitution and Expression Profiling of the NAC Transcription Factor Family in Foxtail Millet (Setaria italica L.). PLoS ONE 2013, 8, e64594.
- Puranik, S.; Sahu, P.P.; Srivastava, P.S.; Prasad, M. NAC Proteins: Regulation and Role in Stress Tolerance. Trends Plant Sci. 2012, 17, 369–381.
- Hong, Y.; Zhang, H.; Huang, L. Overexpression of a Stress-Responsive NAC Transcription Factor Gene ONAC022 Improves Drought and Salt Tolerance in Rice. Front. Plant Sci. 2016, 7, 4.
- Arif, M.; Jan, T.; Riaz, M.; Fahad, S.; Shakoor, M.B.; Amanullah; Rasul, F. Advances in Rice Research for Abiotic Stress Tolerance; Woodhead Publishing: Duxford, UK, 2019.
- Liang, C.; Liu, Y.; Li, Y.; Meng, Z.; Yan, R.; Zhu, T.; Wang, Y.; Kang, S.; Ali Abid, M.; Malik, W.; et al. Activation of ABA Receptors Gene GhPYL9-11A Is Positively Correlated with Cotton Drought Tolerance in Transgenic Arabidopsis. Front. Plant Sci. 2017, 8, 1453.
- Roychoudhury, A.; Chakraborty, M. Biochemical and Molecular Basis of Varietal Difference in Plant Salt Tolerance. Annu. Rev. Res. Biol. 2013, 3, 422–454.
- Jakoby, M.; Weisshaar, B.; Dröge-Laser, W. bZIP Transcription Factors in Arabidopsis. Trends Plant Sci. 2002, 7, 106–111.
- Nijhawan, A.; Jain, M.; Tyagi, A.K.; Khurana, J.P. Genomic Survey and Gene Expression Analysis of the Basic Leucine Zipper Transcription Factor Family in Rice. Plant Physiol. 2008, 146, 333.
- Wei, K.A.; Chen, J.U.A.N.; Wang, Y.A. Genome-Wide Analysis of bZIP-Encoding Genes in Maize. DNA Res. 2012, 19, 463–476.
- Hwang, I.; Manoharan, R.K.; Kang, J. Genome-Wide Identification and Characterization of bZIP Transcription Factors in Brassica Oleracea under Cold Stress. Biomed. Res. Int. 2016, 2016, 4376598.
- Liu, X.; Chu, Z. Genome-Wide Evolutionary Characterization and Analysis of bZIP Transcription Factors and Their Expression Profiles in Response to Multiple Abiotic Stresses in Brachypodium Distachyon. BMC Genom. 2015, 16, 1–15.
- Hezaveh, T.A.; Pourakbar, L.; Rahmani, F.; Alipour, H. Interactive Effects of Salinity and ZnO Nanoparticles on Physiological and Molecular Parameters of Rapeseed (Brassica napus L.). Commun. Soil. Sci. Plant Anal. 2019, 50, 698–715.
- Chen, Z.; Yuan, Y.; Fu, D.; Shen, C.; Yang, Y. Identification and Expression Profiling of the Auxin Response Factors in Dendrobium Officinale under Abiotic Stresses. Int. J. Mol. Sci. 2017, 18, 927.
- Hamzah Saleem, M.; Usman, K.; Rizwan, M.; Al Jabri, H.; Alsafran, M. Functions and Strategies for Enhancing Zinc Availability in Plants for Sustainable Agriculture. Front. Plant Sci. 2022, 13, 1033092.
- Zafar, H.; Ali, A.; Ali, J.S.; Haq, I.U.; Zia, M. Effect of ZnO nanoparticles on Brassica nigra seedlings and stem explants: Growth dynamics and antioxidative response. Front. Plant Sci. 2016, 7, 535.
- Fageria, V.D. Nutrient Interactions in Crop Plants. J. Plant Nutr. 2001, 24, 1269–1290.

