Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dimitar Djilianov | -- | 2242 | 2024-02-26 09:58:23 | | | |
| 2 | Fanny Huang | Meta information modification | 2242 | 2024-02-27 06:15:37 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Djilianov, D.; Moyankova, D.; Mladenov, P.; Topouzova-Hristova, T.; Kostadinova, A.; Staneva, G.; Zasheva, D.; Berkov, S.; Simova-Stoilova, L. Resurrection Plants as Source of Natural Bioactive Compounds. Encyclopedia. Available online: https://encyclopedia.pub/entry/55445 (accessed on 10 June 2026).
Djilianov D, Moyankova D, Mladenov P, Topouzova-Hristova T, Kostadinova A, Staneva G, et al. Resurrection Plants as Source of Natural Bioactive Compounds. Encyclopedia. Available at: https://encyclopedia.pub/entry/55445. Accessed June 10, 2026.
Djilianov, Dimitar, Daniela Moyankova, Petko Mladenov, Tanya Topouzova-Hristova, Aneliya Kostadinova, Galya Staneva, Diana Zasheva, Strahil Berkov, Lyudmila Simova-Stoilova. "Resurrection Plants as Source of Natural Bioactive Compounds" Encyclopedia, https://encyclopedia.pub/entry/55445 (accessed June 10, 2026).
Djilianov, D., Moyankova, D., Mladenov, P., Topouzova-Hristova, T., Kostadinova, A., Staneva, G., Zasheva, D., Berkov, S., & Simova-Stoilova, L. (2024, February 26). Resurrection Plants as Source of Natural Bioactive Compounds. In Encyclopedia. https://encyclopedia.pub/entry/55445
Djilianov, Dimitar, et al. "Resurrection Plants as Source of Natural Bioactive Compounds." Encyclopedia. Web. 26 February, 2024.
Copy Citation
Resurrection plant species are a group of higher plants whose vegetative tissues are able to withstand long periods of almost full desiccation and recover quickly upon rewatering. Apart from being a model system for studying desiccation tolerance, resurrection plant species appear to be a valuable source of metabolites, with various areas of application. A significant number of papers have been published with respect to the extraction and application of bioactive compounds from higher resurrection plant species in various test systems. Promising results have been obtained with respect to antioxidative and antiaging effects in various test systems, particularly regarding valuable anticancer effects in human cell lines.
resurrection plants
bioactive compounds
myconoside
Haberlea rhodopensis
1. Introduction
Plants have been the main source of food, feed, and energy since the very establishment of human civilizations. At the same time, the use of plants’ parts, derivatives, or infusions for other aspects of human welfare also dates back to the roots of human history. Despite the enormous recent progress in natural science, there is still not enough knowledge about extracted plant metabolites and their mode of action when applied to various biological objects. However, positive correlations between the antioxidant properties of numerous plant extracts and infusions and their beneficial potential for human welfare, including medicine, cosmetics, food additives, etc., could be easily made [1]. It is widely accepted that plants’ survival under unfavorable environments is based on, or at least involves, tolerance to oxidative stress [2].
Studies on the mechanisms of environmental stress tolerance of plants could be a good prerequisite and a reason for parallel or consecutive testing of their metabolites as potential bioactive compounds for human benefits. In this respect, the so-called resurrection plants appear to be a very useful model.
Resurrection plants or plants with vegetative desiccation tolerance are a group of higher species able to withstand the drastic decrease in their vegetative tissues’ water content to an almost dry state, and, after long periods of dryness, to recover fast (within hours or 1–2 days) and fully when water is available again [3]. Despite the fact that angiosperm resurrection plants account for less than 0.1% of all higher plant species worldwide, they belong to several botanical families and can be found on every continent, except Antarctica, with habitats in different climate zones and at various altitudes. As could be expected, most of them live under desert or semi-desert conditions. However, there are also species that belong in humid tropical regions in Africa and South America or survive winters with freezing temperatures in Europe [3][4][5][6][7][8][9][10][11]. Their strategies to withstand desiccation are predetermined, constitutive (e.g., in-advance high-abundance of protective compounds, including metabolites), and/or inducible, leading to reprogramming at the transcriptome and metabolome levels upon stress establishment [4][12][13][14][15][16][17].
To survive and recover after extreme water deficit, resurrection plants have evolved complex strategies, including dynamic changes of primary and secondary metabolites [18]. In parallel, the metabolism of these plants attracts additional attention to them as potential sources of compounds with various applications [19][20][21]. Moreover, in comparison with typical medicinal plants, the resurrection species accumulate and/or maintain valuable bioactive compounds in the highest concentrations under desiccation. The accumulated metabolites persist for long periods, which is a good prerequisite for using long-term stored dried samples as sources for further extractions [22].
There is no sound scientific evidence for the ethnobotanical word-of-mouth data related to the potential use of resurrection plants in folk medicine in Eastern Europe or the Pyrenees against human bronchitis, diarrhea, liver diseases, pneumonia, and infectious diseases, or foot-and-mouth disease in livestock [23][24][25].
It appears that probably the only direct connection between indigenous people’s knowledge about resurrection behavior of some local plant species and well-established traditions for using them in folk medicine and religious rituals and current attempts for utilization occurs in southern Africa [26][27]. There are examples of the traditional use of Myrothamnus flabellifolius aqueous extracts or tea infusions against life-threatening conditions or cases of depression and mental disorders [28], as well as in the treatment of a wide range of various other diseases [26][29].
2. Metabolite Profiling and Application of Resurrection Plant Extracts as Bioactive Compounds
Metabolic profiling of resurrection plants is performed by applying contemporary extraction procedures and analytical methods and machinery [21]. With respect to primary metabolism, it is widely accepted that sugars are among the major players in the plant desiccation tolerance complex [18][21]. The dynamics of both sucrose and raffinose family oligosaccharides (RFOs) is well-outlined [30][31], and the specific key role in some resurrection species of relatively rare molecules—e.g., octolose, stachyose, and trehalose—is also discussed [32][33][34]. The high amounts of sugars explain the proposed inclusion of Myrothamnus flabellifolius, the most studied African resurrection species, as a still underestimated but potential source of nutraceuticals [26]. Due to the high amounts of carbohydrates, particularly trehalose, raffinose, and stachyose, along with sucrose, the inclusion of crude extracts in poultry diets has been suggested. Future studies should be performed with a focus on the doses of crude extracts that are added to the feed, since negative effects, such as DNA and chromosomal damage, cell damage, and mutagenic activity, may possibly be observed [35][36].
The data on secondary metabolism in resurrection plants are much more limited, at least in part due to the fact that these plants are still not regarded as sources of valuable natural compounds [21]. The few exceptions available—M. flabellifolius, and the members of the Gesneriaceae (Haberlea rhodopensis, Ramonda ssp., Boea hydrometrica) and of the Linderniaceae (Craterostigma plantagineum, Lindernia brevidens) [31][37][38][39][40][41][42][43]—outline the high abundance and diversity of phenolics, flavonoids, etc.
Unique polyphenols from resurrection plant extracts were identified in high abundance (Figure 1). The 3,4,5-tri-O-galloylquinic acid was identified as a predominant polyphenol in M. flabellifolius. It is accumulated in almost twice the concentration in dry leaves in comparison with fresh [37]. The caffeoyl phenylethanoid glycoside myconoside [β-(3,4-dihydroxyphenyl)-ethyl-3,6-di-O-β-d-apifuranosyl-4-O-α,β-dihydrocaffeoyl-O-β-d-glucopyranoside] was isolated as the most abundant polyphenol in all members of the Gesneriaceae found in Europe, being at the same time resurrection plant species [44][45]. Hispidulin 8-C-(6-O-acetyl-2-O-syringoyl-β-glucopyranoside) was isolated for the first time recently [46] from leaves of H. rhodopensis as acylated hispidoline C-glicoside, possessing some unique features like 2-O-syringoyl and 6-O-acetyl moieties.
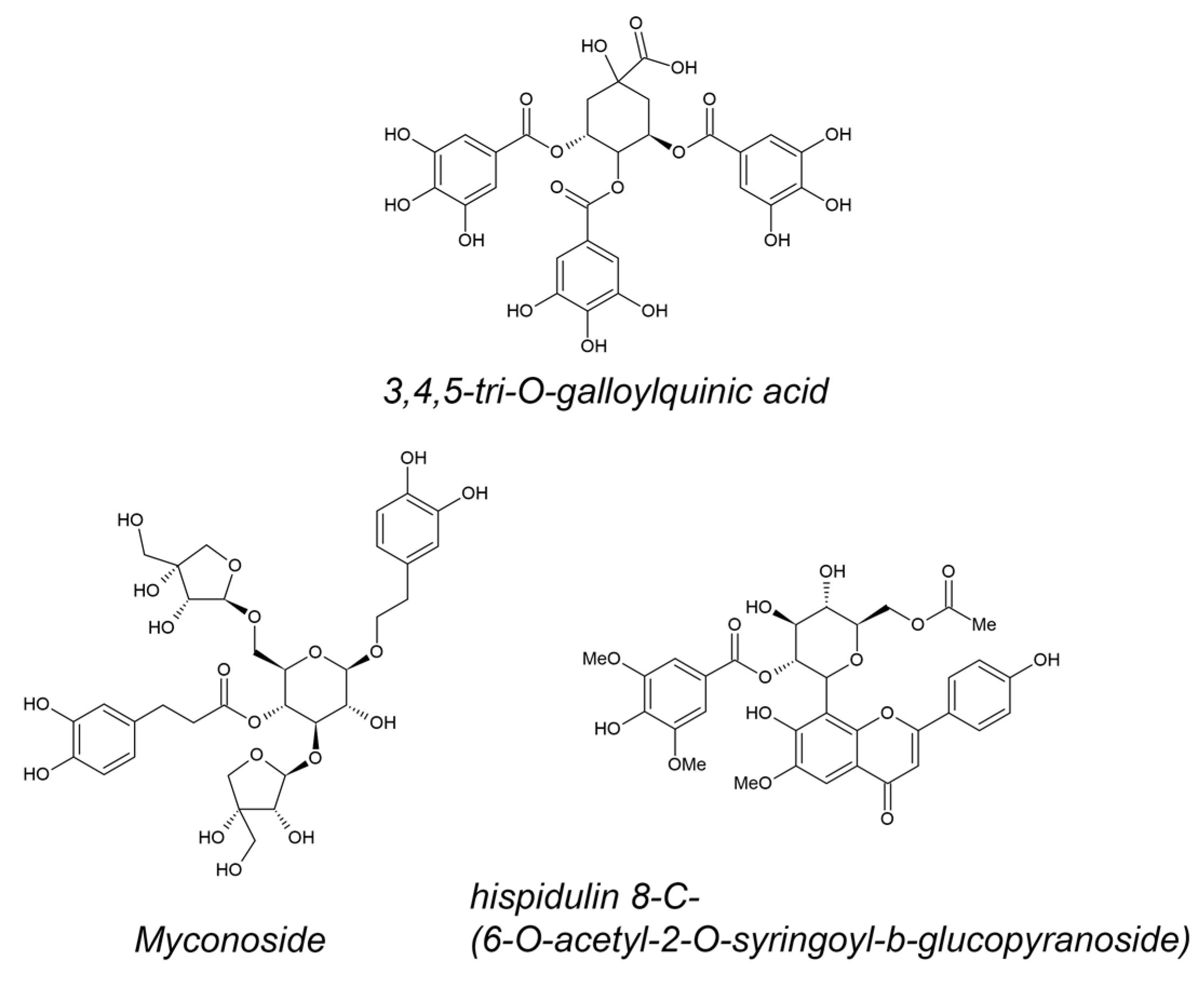
Figure 1. Structural formulas of 3,4,5-tri-O-galloylquinic acid (most abundant in M. flabellifolius), myconoside, and hispidulin 8-C-(6-O-acetyl-2-O-syringoyl-β-glucopyranoside) (most abundant in H. rhodopensis).
The secondary compounds play a significant role in ROS scavenging, thus underlying the desiccation tolerance of the plants [21][47]. The strong antioxidant activity of phenolic compounds also predetermines the interest in using the extracts of resurrection plants for various applications.
Studies on potential utilization of resurrection plants have predominantly been performed with crude total extracts or with their polar/apolar fractions obtained via various solvents. Such experiments are sometimes performed in parallel or hand-in-hand with characterization of the metabolic compounds and evaluation of their antioxidant activities, and efforts have been made to establish a positive correlation with the respective data available.
Metabolic profiling and antioxidant activity studies confirm the potential of Xerophyta spp. as a source of crude extracts for traditional ethnomedicine [48] and pharmacological application [49]. M. flabellifolius is also rich in polyphenols, especially flavonoids, and its extracts effectively suppress the growth of the leukemic cell line HL-60, but not the non-leukemic lymphocytes of the TK6 line [50]. Fractionation of the same plant species’ extracts resulted in active growth suppressing of the triple negative breast cancer cells (TNBCs) from two cell lines, BT-549 and MDA-MB-231, compared to the normal MCF10-A cell line. The main component of the efficient fraction was identified as a derivative of galloyl glucose hexahydroxydiphenic acid called strictinin (chemical name: 3-O-galloyl-4,6-[(S)-hexahydroxydiphenoyl]-b-glucopyranose). Later, strictinin was shown to suppress the activity of Receptor Tyrosine Kinase Orphan-like 1 (ROR1), which is highly active during embryonic development but is not found in growing tissues except in some tumors. In this case, the likely antitumor action is associated with reduced phosphorylation of the AKT kinase and increased apoptosis [51][52]. In another study, the main polyphenol isolated from M. flabellifolius, 3,4,5 tri-O-galloylquinic acid, was found to inhibit HIV-1 and M-MLV reverse transcriptases and could be used as a potent antiviral drug that blocks viral replication [53].
So far, researchers' review of the available literature shows that the potential applications of bioactive compounds isolated from resurrection plants have been predominantly studied in the Balkan endemic plant H. rhodopensis. Data for the biological activity of total extracts or purified compounds are summarized in Figure 2.
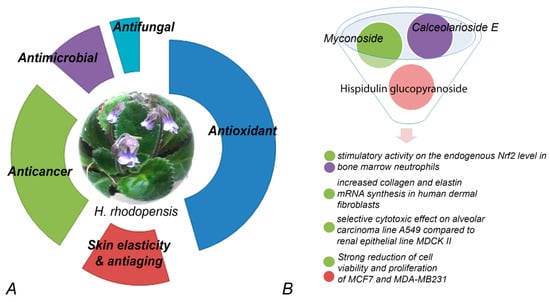
Figure 2. Bioactivity of extracts (A) and compounds (B) purified from H. rhodopensis. (A) Percentage of relative contribution of published studies on the various biological activities of extracts. (B) Myconoside, hispidulin 8-C, and calceolarioside E have been enriched or purified from different fractions. Fractions containing pure myconoside or myconoside in combination with the other two compounds have significant effects on various cell lines.
There was very weak or even no biological activity found in some experimental systems, e.g., no direct virus inactivating effect was found in HSV (Herpes simplex virus) [54], and there was a lack of anticancer effect in some cell lines [55] or very poor antimicrobial activity [56]. On the other hand, promising results were reported when H. rhodopensis extracts were tested as protectors to minimize the harmful effects of radiotherapy [57]. The use of ionizing radiation is one of the widely used approaches in treating various cancers. However, quite often, negative side effects appear during such treatments as a result of the oxidative stress that irradiation has on genomic DNA, lipids, proteins, enzymes, and membranes of living organisms. In this respect, the search for nontoxic and efficient radioprotectors, particularly of plant origin, is very intensive, since a high positive correlation was reported between phenolic compound contents and their antioxidant capacity. An interesting and still not sufficiently investigated issue is the potential use of extracts from resurrection plants’ tissues to manipulate the reaction of higher living organisms to ionizing radiation [57]. Several investigations have been performed in the last 15 years where New Zealand rabbit lines were used as a platform to study the radioprotective properties of H. rhodopensis leaf extracts. Pre-treatment of lymphocyte cultures with such extracts reduced the numbers of aberrant cells and chromosome aberrations in a dose-dependent manner [58][59][60][61] and resulted in a reduction in induced cellular DNA damage [62]. The pre-treatment significantly increased the activity of some antioxidant enzymes and had an anti-lipid peroxidative effect by reducing MDA levels in the blood. Furthermore, a significant reduction in MN events in peripheral lymphocytes was observed. Recently [63], the preincubation of cells with H. rhodopensis extracts was shown to modulate HeLa cancer cells’ early response to gamma IR (γ-IR) and oxidative stress. The response modulation appeared almost immediately after exposure in a dose-dependent manner, thus reducing the severity of genotoxic and oxidative stress. A strong antioxidant effect of methanol extracts of H. rhodopensis was shown in both non-neoplastic and prostate cancer cells, where the extracts reduced H2O2-generated oxidative stress [64]. The pretreatment of non-malignant cell line HEK 293 was apoptosis-protective and cell death-reducing when H2O2-induced oxidative stress was applied. NFκB was activated in p53+/+ cells and suppressed in p53−/− cells. Leaf water and ethanol extracts were applied to a range of other human cancer cell lines [65]. Water extracts were reported not to be antiproliferative, while ethanol extracts were particularly effective to hepatocellular carcinoma (HepG2) and non-small cell lung adenocarcinoma (A549) cell lines and were found to exert significant antimigratory concentration-dependent effects in both cell lines.
Total methanol extracts and polar and apolar fractions were tested in a completely different system—important plant pathogens to search for potential sustainable and eco-friendly plant protection approaches [66][67]. No fungitoxic effect on Alternaria alternata and Fusarium oxysporum was found. Strong inhibition of Botrytis cinerea was achieved, in particular by apolar fractions. The same fractions stimulated the growth of Phytophthora citricola and can be potentially used as an effector. Other Phythophtora spp. isolates were stimulated significantly to grow under in vitro conditions, which could be a good prerequisite for the development of culture media for further tests on these obligate pathogens.
The significant amount of myconoside in plant extracts from European Gesneriads along with the strong scavenging activity gave ground for potential application in various human welfare areas. There is a significant interest in the application of myconoside as a single compound or in well-characterized combination with other secondary metabolites in various medicinal test systems.
An extract rich in myconoside isolated from H. rhodopensis was reported to increase mRNA synthesis of collagen and elastin genes in human dermal fibroblasts stressed with H2O2 [68]. The extracts were proven to protect against UV-induced dermis oxidation and even increased skin elasticity of human volunteers. It was suggested that H. rhodopensis extracts can be used for anti-aging treatments, protecting the skin from oxidation, increasing skin elasticity, and enhancing skin radiance. The anti-aging potential of such extracts was further confirmed. Influence on cell periphery, permeabilization of the membrane, and disruption of HaCaT keratinocyte tight junctions were observed—more pronounced in actively dividing cells [69]. In a very different experimental system, the strong and specific proliferative, anti-aging, and protective effect was found in the model yeast Saccharomyces cerevisiae cell line to revitalize and ameliorate cellular growth as well as to balance intracellular metabolic states [70]. Myconoside, isolated from H. rhodopensis extracts, was shown to have strong antioxidative potential along with a significant hepatoprotective effect on isolated rat hepatocytes [71]. H. rhodopensis extract fractions, containing myconoside or enriched with Calceolarioside E, were found to be very effective in promoting the expression of nuclear factor erythroid 2 p45-related factor 2 (Nrf2), a transcriptional regulator of the cellular redox balance and a suppressor of the pathological manifestation of various diseases, in bone marrow neutrophils [72].
References
- Bistgani, Z.E.; Barker, A.V.; Hashemi, M. Physiology of Medicinal and Aromatic Plants under Drought Stress. Crop J. 2024.
- Dumanović, J.; Nepovimova, E.; Natić, M.; Kuča, K.; Jaćević, V. The Significance of Reactive Oxygen Species and Antioxidant Defense System in Plants: A Concise Overview. Front. Plant Sci. 2021, 11, 552969.
- Gaff, D.F.; Oliver, M. The Evolution of Desiccation Tolerance in Angiosperm Plants: A Rare yet Common Phenomenon. Funct. Plant Biol. 2013, 40, 315–328.
- Dinakar, C.; Bartels, D. Desiccation Tolerance in Resurrection Plants: New Insights from Transcriptome, Proteome and Metabolome Analysis. Front. Plant Sci. 2013, 4, 482.
- Suguiyama, V.; da Silva, E.; Meirelles, S.; Centeno, D.; Braga, M. Leaf Metabolite Profile of the Brazilian Resurrection Plant Barbacenia purpurea Hook. (Velloziaceae) Shows Two Time-Dependent Responses during Desiccation and Recovering. Front. Plant Sci. 2014, 5, 96.
- Legardón, A.; García-Plazaola, J.I. Gesneriads, a Source of Resurrection and Double-Tolerant Species: Proposal of New Desiccation- and Freezing-Tolerant Plants and Their Physiological Adaptations. Biology 2023, 12, 107.
- Rakic, T.; Lazarevic, M.; Jovanovic, Z.; Radovic, S.; Siljak-Yakovlev, S.; Stevanovic, B.; Stevanovic, V. Resurrection Plants of the Genus Ramonda: Prospective Survival Strategies—Unlock Further Capacity of Adaptation, or Embark on the Path of Evolution? Front. Plant Sci. 2014, 4, 550.
- Okemo, P.A.; Njaci, I.; Kim, Y.-M.; McClure, R.S.; Peterson, M.J.; Beliaev, A.S.; Hixson, K.K.; Mundree, S.; Williams, B. Tripogon loliiformis Tolerates Rapid Desiccation after Metabolic and Transcriptional Priming during Initial Drying. Sci. Rep. 2023, 13, 20613.
- Mihailova, G.; Solti, Á.; Sárvári, É.; Hunyadi-Gulyás, É.; Georgieva, K. Protein Changes in Shade and Sun Haberlea rhodopensis Leaves during Dehydration at Optimal and Low Temperatures. Plants 2023, 12, 401.
- Mihaylova, D.; Bahchevanska, S.; Toneva, V. Examination of the Antioxidant Activity of Haberlea rhodopensis Leaf Extracts and Their Phenolic Constituents. J. Food Biochem. 2013, 37, 255–261.
- Gechev, T.; Lyall, R.; Petrov, V.; Bartels, D. Systems Biology of Resurrection Plants. Cell. Mol. Life Sci. 2021, 78, 6365–6394.
- Gechev, T.S.; Benina, M.; Obata, T.; Tohge, T.; Sujeeth, N.; Minkov, I.; Hille, J.; Temanni, M.-R.; Marriott, A.S.; Bergström, E.; et al. Molecular Mechanisms of Desiccation Tolerance in the Resurrection Glacial Relic Haberlea rhodopensis. Cell. Mol. Life Sci. 2013, 70, 689–709.
- Liu, J.; Moyankova, D.; Lin, C.-T.; Mladenov, P.; Sun, R.-Z.; Djilianov, D.; Deng, X. Transcriptome Reprogramming during Severe Dehydration Contributes to Physiological and Metabolic Changes in the Resurrection Plant Haberlea rhodopensis. BMC Plant Biol. 2018, 18, 351.
- Tebele, S.M.; Marks, R.A.; Farrant, J.M. Two Decades of Desiccation Biology: A Systematic Review of the Best Studied Angiosperm Resurrection Plants. Plants 2021, 10, 2784.
- Liu, J.; Moyankova, D.; Djilianov, D.; Deng, X. Common and Specific Mechanisms of Desiccation Tolerance in Two Gesneriaceae Resurrection Plants. Multiomics Evidences. Front. Plant Sci. 2019, 10, 1067.
- Mladenov, P.; Zasheva, D.; Planchon, S.; Leclercq, C.C.; Falconet, D.; Moyet, L.; Brugière, S.; Moyankova, D.; Tchorbadjieva, M.; Ferro, M.; et al. Proteomics Evidence of a Systemic Response to Desiccation in the Resurrection Plant Haberlea rhodopensis. Int. J. Mol. Sci. 2022, 23, 8520.
- Mladenov, P.; Wang, X.; Yang, Z.; Djilianov, D.; Deng, X. Dynamics of Chromatin Accessibility and Genome Wide Control of Desiccation Tolerance in the Resurrection Plant Haberlea rhodopensis. BMC Plant Biol. 2023, 23, 654.
- Oliver, M.J.; Farrant, J.M.; Hilhorst, H.W.M.; Mundree, S.; Williams, B.; Bewley, J.D. Desiccation Tolerance: Avoiding Cellular Damage During Drying and Rehydration. Ann. Rev. Plant Biol. 2020, 71, 435–460.
- Reshi, Z.A.; Ahmad, W.; Lukatkin, A.S.; Javed, S.B. From Nature to Lab: A Review of Secondary Metabolite Biosynthetic Pathways, Environmental Influences, and In Vitro Approaches. Metabolites 2023, 13, 895.
- Blomstedt, C.K.; Griffiths, C.A.; Gaff, D.F.; Hamill, J.D.; Neale, A.D. Plant Desiccation Tolerance and Its Regulation in the Foliage of Resurrection “Flowering-Plant” Species. Agronomy 2018, 8, 146.
- Dace, H.J.W.; Adetunji, A.E.; Moore, J.P.; Farrant, J.M.; Hilhorst, H.W.M. A Review of the Role of Metabolites in Vegetative Desiccation Tolerance of Angiosperms. Curr. Opin. Plant Biol. 2023, 75, 102410.
- Moyankova, D.; Djilianov, D. Time- and Space-Saving Procedure to Obtain Extracts with Antioxidative Properties from Haberlea rhodopensis. C. R. L’académie Bulg. Sci. 2016, 69, 879–884.
- Georgiev, Y.N.; Ognyanov, M.H.; Denev, P.N. The Ancient Thracian Endemic Plant Haberlea rhodopensis Friv. and Related Species: A Review. J. Ethnopharmacol. 2020, 249, 112359.
- Rigat, M.; Bonet, M.À.; Garcia, S.; Garnatje, T.; Vallès, J. Studies on Pharmaceutical Ethnobotany in the High River Ter Valley (Pyrenees, Catalonia, Iberian Peninsula). J. Ethnopharmacol. 2007, 113, 267–277.
- Agelet, A.; Vallès, J. Studies on Pharmaceutical Ethnobotany in the Region of Pallars (Pyrenees, Catalonia, Iberian Peninsula). Part II. New or Very Rare Uses of Previously Known Medicinal Plants. J. Ethnopharmacol. 2003, 84, 211–227.
- Nantapo, C.W.T.; Marume, U. Exploring the Potential of Myrothamnus flabellifolius Welw. (Resurrection Tree) as a Phytogenic Feed Additive in Animal Nutrition. Animals 2022, 12, 1973.
- Gahamanyi, N.; Munyaneza, E.; Dukuzimana, E.; Tuyiringire, N.; Pan, C.-H.; Komba, E.V.G. Ethnobotany, Ethnopharmacology, and Phytochemistry of Medicinal Plants Used for Treating Human Diarrheal Cases in Rwanda: A Review. Antibiotics 2021, 10, 1231.
- Erhabor, J.O.; Komakech, R.; Kang, Y.; Tang, M.; Matsabisa, M.G. Ethnopharmacological Importance and Medical Applications of Myrothamnus flabellifolius Welw. (Myrothamnaceae)—A Review. J. Ethnopharmacol. 2020, 252, 112576.
- Gessner, D.K.; Ringseis, R.; Eder, K. Potential of Plant Polyphenols to Combat Oxidative Stress and Inflammatory Processes in Farm Animals. J. Anim. Physiol. Anim. Nutr. 2017, 101, 605–628.
- Peters, S.; Mundree, S.G.; Thomson, J.A.; Farrant, J.M.; Keller, F. Protection Mechanisms in the Resurrection Plant Xerophyta viscosa (Baker): Both Sucrose and Raffinose Family Oligosaccharides (RFOs) Accumulate in Leaves in Response to Water Deficit. J. Exp. Bot. 2007, 58, 1947–1956.
- Djilianov, D.; Ende, W.V.D.; Alexieva, V.; Moyankova, D. Sugar Ratios, Glutathione Redox Status and Phenols in the Resurrection Species Haberlea rhodopensis and the Closely Related Non-Resurrection Species Chirita eberhardtii. Plant Biol. 2011, 13, 767–776.
- Ghasempour, H.R.; Gaff, D.F.; Williams, R.P.W.; Gianello, R.D. Contents of Sugars in Leaves of Drying Desiccation Tolerant Flowering Plants, Particularly Grasses. Plant Growth Regul. 1998, 24, 185–191.
- Farrant, J.M.; Cooper, K.; Hilgart, A.; Abdalla, K.O.; Bentley, J.; Thomson, J.A.; Dace, H.J.W.; Peton, N.; Mundree, S.G.; Rafudeen, M.S. A Molecular Physiological Review of Vegetative Desiccation Tolerance in the Resurrection Plant Xerophyta viscosa (Baker). Planta 2015, 242, 407–426.
- Zhang, Q.; Bartels, D. Octulose: A Forgotten Metabolite? J. Exp. Bot. 2017, 68, 5689–5694.
- Cheikhyoussef, A.; Summers, R.; Kahaka, G. Qualitative and Quantitative Analysis of Phytochemical Compounds in Namibian Myrothamnus flabellifolius. Int. Sci. Technol. J. Namibia 2015, 5, 71–83.
- Abdelli, N.; Solà-Oriol, D.; Pérez, J.F. Phytogenic Feed Additives in Poultry: Achievements, Prospective and Challenges. Animals 2021, 11, 3471.
- Moore, J.P.; Westall, K.L.; Ravenscroft, N.; Farrant, J.M.; Lindsey, G.G.; Brandt, W.F. The Predominant Polyphenol in the Leaves of the Resurrection Plant Myrothamnus flabellifolius, 3,4,5 Tri-O-Galloylquinic Acid, Protects Membranes against Desiccation and Free Radical-Induced Oxidation. Biochem. J. 2004, 385, 301–308.
- Veljovic-Jovanovic, S.; Kukavica, B.; Navari-Izzo, F. Characterization of Polyphenol Oxidase Changes Induced by Desiccation of Ramonda serbica Leaves. Physiol. Plant 2008, 132, 407–416.
- Feng, W.-S.; Li, Y.-J.; Zheng, X.-K.; Wang, Y.-Z.; Su, F.-Y. Chemical Constituents of Boea hygrometrica. Chin. J. Nat. Med. 2011, 9, 406–409.
- Moyankova, D.; Mladenov, P.; Berkov, S.; Peshev, D.; Georgieva, D.; Djilianov, D. Metabolic Profiling of the Resurrection Plant Haberlea rhodopensis during Desiccation and Recovery. Physiol. Plant 2014, 152, 675–687.
- Vidović, M.; Battisti, I.; Pantelić, A.; Morina, F.; Arrigoni, G.; Masi, A.; Jovanović, S.V. Desiccation Tolerance in Ramonda serbica Panc.: An Integrative Transcriptomic, Proteomic, Metabolite and Photosynthetic Study. Plants 2022, 11, 1199.
- Gođevac, D.; Ivanović, S.; Simić, K.; Anđelković, B.; Jovanović, Ž.; Rakić, T. Metabolomics Study of the Desiccation and Recovery Process in the Resurrection Plants Ramonda serbica and R. nathaliae. Phytochem. Anal. 2022, 33, 961–970.
- Passon, M.; Weber, F.; Jung, N.U.; Bartels, D. Profiling of Phenolic Compounds in Desiccation-Tolerant and Non-Desiccation-Tolerant Linderniaceae. Phytochem. Anal. 2021, 32, 521–529.
- Cañigueral, S.; Salvía, M.J.; Vila, R.; Iglesias, J.; Virgili, A.; Parella, T. New Polyphenol Glycosides from Ramonda myconi. J. Nat. Prod. 1996, 59, 419–422.
- Jensen, S.R. Caffeoyl Phenylethanoid Glycosides in Sanango racemosum and in the Gesneriaceae. Phytochemistry 1996, 43, 777–783.
- Ebrahimi, S.N.; Gafner, F.; Dell’Acqua, G.; Schweikert, K.; Hamburger, M. Flavone 8-C-Glycosides from Haberlea rhodopensis Friv. (Gesneriaceae). Helv. Chim. Acta 2011, 94, 38–45.
- Gechev, T.S.; Hille, J.; Woerdenbag, H.J.; Benina, M.; Mehterov, N.; Toneva, V.; Fernie, A.R.; Mueller-Roeber, B. Natural Products from Resurrection Plants: Potential for Medical Applications. Biotechnol. Adv. 2014, 32, 1091–1101.
- Nyalo, P.; Omwenga, G.; Ngugi, M. Quantitative Phytochemical Profile and In Vitro Antioxidant Properties of Ethyl Acetate Extracts of Xerophyta spekei (Baker) and Grewia tembensis (Fresen). J. Evid. Based Complement. Altern. Med. 2023, 28, 2515690X231165096.
- Da Costa, D.J.; Leitão, A.; Faria, R.X.; Anholeti, M.C.; Nunes, M.A.; Oliveira, M.B.P.; Da Costa Santos, W.; De Barros Machado, T. Preliminary Phytochemical Analysis of the Ethanolic Extract of Xerophyta stenophylla Baker. Res. Soc. Dev. 2022, 11, e38211528319.
- Dhillon, J.; Miller, V.; Carter, J.; Badiab, A.; Tang, C.N.; Huynh, A.; Peethambaran, B. Apoptosis-Inducing Potential of Myrothamnus flabellifolius, an Edible Medicinal Plant, on Human Myeloid Leukemia HL-60 Cells. Int. J. Appl. Res. Nat. Prod. 2014, 7, 28–32.
- Brar, J.; Fultang, N.; Askey, K.; Tettamanzi, M.C.; Peethambaran, B. A Novel Anti-Triple Negative Breast Cancer Compound Isolated from Medicinal Herb Myrothamnus flabellifolius. J. Med. Plants Res. 2018, 12, 7–14.
- Fultang, N.; Brar, J.; Mercier, I.; Klase, Z.; Peethambaran, B. Myrothamnus flabellifolius Selectively Targets Triple Negative Breast Cancer in Vitro, Restoring Tamoxifen Sensitivity through Modulation of MiRNAs Associated with Estrogen Receptors. Int. J. Appl. Res. Nat. Prod. 2018, 11, 24–33.
- Kamng’ona, A.; Moore, J.P.; Lindsey, G.; Brandt, W. Inhibition of HIV-1 and M-MLV Reverse Transcriptases by a Major Polyphenol (3,4,5 Tri-O-Galloylquinic Acid) Present in the Leaves of the South African Resurrection Plant, Myrothamnus flabellifolia. J. Enzym. Inhib. Med. Chem. 2011, 26, 843–853.
- Moyankova, D.; Hinkov, A.; Shishkov, S.; Djilianov, D. Inhibitory Effect of Extracts from Haberlea rhodopensis Friv. against Herpes Simplex Virus. C. R. L’académie Bulg. Sci. 2014, 76, 1369–1376.
- Berkov, S.; Nikolova, M.; Hristozova, N.; Momekov, G.; Ionkova, I.; Djilianov, D. GC-MS Profiling of Bioactive Extracts from Haberlea rhodopensis: An Endemic Resurrection Plant. J. Serbian Chem. Soc. 2011, 76, 211–220.
- Radev, R.; Lazarova, G.; Nedialkov, P.; Sokolova, K.; Rukanova, D.; Tsokeva, Z. Study on Antibacterial Activity of Haberlea rhodopensis. Trakia J. Sci. 2008, 7, 34–36.
- Georgieva, S.; Gencheva, D.; Popov, B.; Grozeva, N.; Zhelyazkova, M. Radioprotective Action of Resurrection Plant Haberlea rhodopensis Friv. (Gesneriaceae) and Role of Flavonoids and Phenolic Acids. Bulg. J. Agric. Sci. 2019, 25, 158–168.
- Popov, B.; Georgieva, S.; Oblakova, M.; Bonev, G. Effects of Haberlea rhodopensis Extract on Antioxidation and Lipid Peroxidation in Rabbits after Exposure to 60Co-γ-Rays. Arch. Biol. Sci. 2013, 65, 91–97.
- Popov, B.; Radev, R.; Georgieva, S. In Vitro Incidence of Chromosome Aberrations in Gamma-Irradiated Rabbit Lymphocytes, Treated with Haberlea rhodopensis Extract and Vitamin C. Bulg. J. Vet. Med. 2010, 13, 148–153.
- Dobreva, Z.G.; Popov, B.N.; Georgieva, S.Y.; Stanilova, S.A. Immunostimulatory Activities of Haberlea rhodopensis Leaf Extract on the Specific Antibody Response: Protective Effects against γ-Radiation-Induced Immunosuppression. Food Agric. Immunol. 2015, 26, 381–393.
- Popov, B.; Georgieva, S.; Gadjeva, V. Modulatory Effects of Total Extract of Haberlea rhodopensis against the Cyclophosphamide Induced Genotoxicity in Rabbit Lymphocytes in Vivo. Trakia J. Sci. 2011, 9, 51–57.
- Georgieva, S.; Popov, B.; Bonev, G. Radioprotective Effect of Haberlea rhodopensis (Friv.) Leaf Extract on Gamma-Radiation-Induced DNA Damage, Lipid Peroxidation and Antioxidant Levels in Rabbit Blood. Indian. J. Exp. Biol. 2013, 51, 29–36.
- Staneva, D.; Dimitrova, N.; Popov, B.; Alexandrova, A.; Georgieva, M.; Miloshev, G. Haberlea rhodopensis Extract Tunes the Cellular Response to Stress by Modulating DNA Damage, Redox Components, and Gene Expression. Int. J. Mol. Sci. 2023, 24, 15964.
- Hayrabedyan, S.; Todorova, K.; Zasheva, D.; Moyankova, D.; Georgieva, D.; Todorova, J.; Djilianov, D. Haberlea rhodopensis Has Potential as a New Drug Source Based on Its Broad Biological Modalities. Biotechnol. Biotechnol. Equip. 2013, 27, 3553–3560.
- Spyridopoulou, K.; Kyriakou, S.; Nomikou, A.; Roupas, A.; Ermogenous, A.; Karamanoli, K.; Moyankova, D.; Djilianov, D.; Galanis, A.; Panayiotidis, M.I.; et al. Chemical Profiling, Antiproliferative and Antimigratory Capacity of Haberlea rhodopensis Extracts in an In Vitro Platform of Various Human Cancer Cell Lines. Antioxidants 2022, 11, 2305.
- Moyankova, D.; Lyubenova, A.; Slavov, S.; Djilianov, D. Extracts of the Endemic Resurrection Plant Haberlea rhodopensis Stimulate In Vitro Growth of Various Phytophthora Spp. Pathogens. Eur. J. Plant Pathol. 2014, 138, 149–155.
- Moyankova, D.; Georgieva, D.; Batchvarova, R.; Slavov, S.; Djilianov, D. Effect of Extracts from the Resurrection Plant Haberlea rhodopensis on in Vitro Growth of Plant Pathogens. C. R. L’académie Bulg. Sci. 2013, 66, 1269–1272.
- Dell’Acqua, G.; Schweikert, K. Skin Benefits of a Myconoside-Rich Extract from Resurrection Plant Haberlea rhodopensis. Int. J. Cosmet. Sci. 2012, 34, 132–139.
- Kostadinova, A.; Doumanov, J.; Moyankova, D.; Ivanov, S.; Mladenova, K.; Djilianov, D.; Topuzova-Hristova, T. Haberlea rhodopensis Extracts Affect Cell Periphery of Keratinocytes. C. R. L’académie Bulg. Sci. 2016, 69, 439–448.
- Georgieva, M.; Moyankova, D.; Djilianov, D.; Uzunova, K.; Miloshev, G. Methanol Extracts from the Resurrection Plant Haberlea rhodopensis Ameliorate Cellular Vitality in Chronologically Ageing Saccharomyces Cerevisiae Cells. Biogerontology 2015, 16, 461–472.
- Kondeva-Burdina, M.; Zheleva-Dimitrova, D.; Nedialkov, P.; Girreser, U.; Mitcheva, M. Cytoprotective and Antioxidant Effects of Phenolic Compounds from Haberlea rhodopensis Friv. (Gesneriaceae). Pharmacogn. Mag. 2013, 9, 294–301.
- Amirova, K.M.; Dimitrova, P.A.; Marchev, A.S.; Krustanova, S.V.; Simova, S.D.; Alipieva, K.I.; Georgiev, M.I. Biotechnologically-Produced Myconoside and Calceolarioside E Induce Nrf2 Expression in Neutrophils. Int. J. Mol. Sci. 2021, 22, 1759.
More
Information
Subjects:
Plant Sciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
621
Revisions:
2 times
(View History)
Update Date:
27 Feb 2024
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No