Your browser does not fully support modern features. Please upgrade for a smoother experience.
Please note this is a comparison between Version 1 by Dimitar Djilianov and Version 2 by Fanny Huang.
Resurrection plant species are a group of higher plants whose vegetative tissues are able to withstand long periods of almost full desiccation and recover quickly upon rewatering. Apart from being a model system for studying desiccation tolerance, resurrection plant species appear to be a valuable source of metabolites, with various areas of application. A significant number of papers have been published with respect to the extraction and application of bioactive compounds from higher resurrection plant species in various test systems. Promising results have been obtained with respect to antioxidative and antiaging effects in various test systems, particularly regarding valuable anticancer effects in human cell lines.
- resurrection plants
- bioactive compounds
- myconoside
- Haberlea rhodopensis
1. Introduction
Plants have been the main source of food, feed, and energy since the very establishment of human civilizations. At the same time, the use of plants’ parts, derivatives, or infusions for other aspects of human welfare also dates back to the roots of human history. Despite the enormous recent progress in natural science, there is still not enough knowledge about extracted plant metabolites and their mode of action when applied to various biological objects. However, positive correlations between the antioxidant properties of numerous plant extracts and infusions and their beneficial potential for human welfare, including medicine, cosmetics, food additives, etc., could be easily made [1]. It is widely accepted that plants’ survival under unfavorable environments is based on, or at least involves, tolerance to oxidative stress [2].
Studies on the mechanisms of environmental stress tolerance of plants could be a good prerequisite and a reason for parallel or consecutive testing of their metabolites as potential bioactive compounds for human benefits. In this respect, the so-called resurrection plants appear to be a very useful model.
Resurrection plants or plants with vegetative desiccation tolerance are a group of higher species able to withstand the drastic decrease in their vegetative tissues’ water content to an almost dry state, and, after long periods of dryness, to recover fast (within hours or 1–2 days) and fully when water is available again [3]. Despite the fact that angiosperm resurrection plants account for less than 0.1% of all higher plant species worldwide, they belong to several botanical families and can be found on every continent, except Antarctica, with habitats in different climate zones and at various altitudes. As could be expected, most of them live under desert or semi-desert conditions. However, there are also species that belong in humid tropical regions in Africa and South America or survive winters with freezing temperatures in Europe [3][4][5][6][7][8][9][10][11][3,4,5,6,7,8,9,10,11]. Their strategies to withstand desiccation are predetermined, constitutive (e.g., in-advance high-abundance of protective compounds, including metabolites), and/or inducible, leading to reprogramming at the transcriptome and metabolome levels upon stress establishment [4][12][13][14][15][16][17][4,12,13,14,15,16,17].
To survive and recover after extreme water deficit, resurrection plants have evolved complex strategies, including dynamic changes of primary and secondary metabolites [18]. In parallel, the metabolism of these plants attracts additional attention to them as potential sources of compounds with various applications [19][20][21][19,20,21]. Moreover, in comparison with typical medicinal plants, the resurrection species accumulate and/or maintain valuable bioactive compounds in the highest concentrations under desiccation. The accumulated metabolites persist for long periods, which is a good prerequisite for using long-term stored dried samples as sources for further extractions [22].
There is no sound scientific evidence for the ethnobotanical word-of-mouth data related to the potential use of resurrection plants in folk medicine in Eastern Europe or the Pyrenees against human bronchitis, diarrhea, liver diseases, pneumonia, and infectious diseases, or foot-and-mouth disease in livestock [23][24][25][23,24,25].
It appears that probably the only direct connection between indigenous people’s knowledge about resurrection behavior of some local plant species and well-established traditions for using them in folk medicine and religious rituals and current attempts for utilization occurs in southern Africa [26][27][26,27]. There are examples of the traditional use of Myrothamnus flabellifolius aqueous extracts or tea infusions against life-threatening conditions or cases of depression and mental disorders [28], as well as in the treatment of a wide range of various other diseases [26][29][26,29].
2. Metabolite Profiling and Application of Resurrection Plant Extracts as Bioactive Compounds
Metabolic profiling of resurrection plants is performed by applying contemporary extraction procedures and analytical methods and machinery [21]. With respect to primary metabolism, it is widely accepted that sugars are among the major players in the plant desiccation tolerance complex [18][21][18,21]. The dynamics of both sucrose and raffinose family oligosaccharides (RFOs) is well-outlined [30][31][31,32], and the specific key role in some resurrection species of relatively rare molecules—e.g., octolose, stachyose, and trehalose—is also discussed [32][33][34][33,34,35]. The high amounts of sugars explain the proposed inclusion of Myrothamnus flabellifolius, the most studied African resurrection species, as a still underestimated but potential source of nutraceuticals [26]. Due to the high amounts of carbohydrates, particularly trehalose, raffinose, and stachyose, along with sucrose, the inclusion of crude extracts in poultry diets has been suggested. Future studies should be performed with a focus on the doses of crude extracts that are added to the feed, since negative effects, such as DNA and chromosomal damage, cell damage, and mutagenic activity, may possibly be observed [35][36][36,37].
The data on secondary metabolism in resurrection plants are much more limited, at least in part due to the fact that these plants are still not regarded as sources of valuable natural compounds [21]. The few exceptions available—M. flabellifolius, and the members of the Gesneriaceae (Haberlea rhodopensis, Ramonda ssp., Boea hydrometrica) and of the Linderniaceae (Craterostigma plantagineum, Lindernia brevidens) [31][37][38][39][40][41][42][43][32,38,39,40,41,42,43,44]—outline the high abundance and diversity of phenolics, flavonoids, etc.
Unique polyphenols from resurrection plant extracts were identified in high abundance (Figure 1). The 3,4,5-tri-O-galloylquinic acid was identified as a predominant polyphenol in M. flabellifolius. It is accumulated in almost twice the concentration in dry leaves in comparison with fresh [37][38]. The caffeoyl phenylethanoid glycoside myconoside [β-(3,4-dihydroxyphenyl)-ethyl-3,6-di-O-β-d-apifuranosyl-4-O-α,β-dihydrocaffeoyl-O-β-d-glucopyranoside] was isolated as the most abundant polyphenol in all members of the Gesneriaceae found in Europe, being at the same time resurrection plant species [44][45][45,46]. Hispidulin 8-C-(6-O-acetyl-2-O-syringoyl-β-glucopyranoside) was isolated for the first time recently [46][47] from leaves of H. rhodopensis as acylated hispidoline C-glicoside, possessing some unique features like 2-O-syringoyl and 6-O-acetyl moieties.
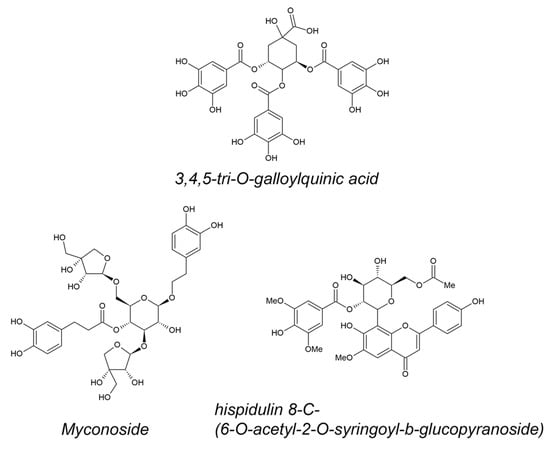
Figure 1. Structural formulas of 3,4,5-tri-O-galloylquinic acid (most abundant in M. flabellifolius), myconoside, and hispidulin 8-C-(6-O-acetyl-2-O-syringoyl-β-glucopyranoside) (most abundant in H. rhodopensis).
The secondary compounds play a significant role in ROS scavenging, thus underlying the desiccation tolerance of the plants [21][47][21,30]. The strong antioxidant activity of phenolic compounds also predetermines the interest in using the extracts of resurrection plants for various applications.
Studies on potential utilization of resurrection plants have predominantly been performed with crude total extracts or with their polar/apolar fractions obtained via various solvents. Such experiments are sometimes performed in parallel or hand-in-hand with characterization of the metabolic compounds and evaluation of their antioxidant activities, and efforts have been made to establish a positive correlation with the respective data available.
Metabolic profiling and antioxidant activity studies confirm the potential of Xerophyta spp. as a source of crude extracts for traditional ethnomedicine [48] and pharmacological application [49]. M. flabellifolius is also rich in polyphenols, especially flavonoids, and its extracts effectively suppress the growth of the leukemic cell line HL-60, but not the non-leukemic lymphocytes of the TK6 line [50]. Fractionation of the same plant species’ extracts resulted in active growth suppressing of the triple negative breast cancer cells (TNBCs) from two cell lines, BT-549 and MDA-MB-231, compared to the normal MCF10-A cell line. The main component of the efficient fraction was identified as a derivative of galloyl glucose hexahydroxydiphenic acid called strictinin (chemical name: 3-O-galloyl-4,6-[(S)-hexahydroxydiphenoyl]-b-glucopyranose). Later, strictinin was shown to suppress the activity of Receptor Tyrosine Kinase Orphan-like 1 (ROR1), which is highly active during embryonic development but is not found in growing tissues except in some tumors. In this case, the likely antitumor action is associated with reduced phosphorylation of the AKT kinase and increased apoptosis [51][52][66,74]. In another study, the main polyphenol isolated from M. flabellifolius, 3,4,5 tri-O-galloylquinic acid, was found to inhibit HIV-1 and M-MLV reverse transcriptases and could be used as a potent antiviral drug that blocks viral replication [53][71].
So far, researchers' review of the available literature shows that the potential applications of bioactive compounds isolated from resurrection plants have been predominantly studied in the Balkan endemic plant H. rhodopensis. Data for the biological activity of total extracts or purified compounds are summarized in Figure 2.
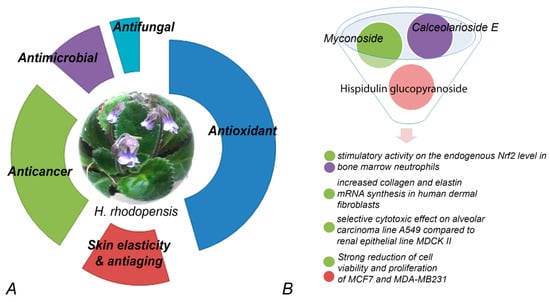
Figure 2. Bioactivity of extracts (A) and compounds (B) purified from H. rhodopensis. (A) Percentage of relative contribution of published studies on the various biological activities of extracts. (B) Myconoside, hispidulin 8-C, and calceolarioside E have been enriched or purified from different fractions. Fractions containing pure myconoside or myconoside in combination with the other two compounds have significant effects on various cell lines.
There was very weak or even no biological activity found in some experimental systems, e.g., no direct virus inactivating effect was found in HSV (Herpes simplex virus) [54][59], and there was a lack of anticancer effect in some cell lines [55][65] or very poor antimicrobial activity [56][63]. On the other hand, promising results were reported when H. rhodopensis extracts were tested as protectors to minimize the harmful effects of radiotherapy [57][75]. The use of ionizing radiation is one of the widely used approaches in treating various cancers. However, quite often, negative side effects appear during such treatments as a result of the oxidative stress that irradiation has on genomic DNA, lipids, proteins, enzymes, and membranes of living organisms. In this respect, the search for nontoxic and efficient radioprotectors, particularly of plant origin, is very intensive, since a high positive correlation was reported between phenolic compound contents and their antioxidant capacity. An interesting and still not sufficiently investigated issue is the potential use of extracts from resurrection plants’ tissues to manipulate the reaction of higher living organisms to ionizing radiation [57][75]. Several investigations have been performed in the last 15 years where New Zealand rabbit lines were used as a platform to study the radioprotective properties of H. rhodopensis leaf extracts. Pre-treatment of lymphocyte cultures with such extracts reduced the numbers of aberrant cells and chromosome aberrations in a dose-dependent manner [58][59][60][61][52,53,54,55] and resulted in a reduction in induced cellular DNA damage [62][56]. The pre-treatment significantly increased the activity of some antioxidant enzymes and had an anti-lipid peroxidative effect by reducing MDA levels in the blood. Furthermore, a significant reduction in MN events in peripheral lymphocytes was observed. Recently [63][57], the preincubation of cells with H. rhodopensis extracts was shown to modulate HeLa cancer cells’ early response to gamma IR (γ-IR) and oxidative stress. The response modulation appeared almost immediately after exposure in a dose-dependent manner, thus reducing the severity of genotoxic and oxidative stress. A strong antioxidant effect of methanol extracts of H. rhodopensis was shown in both non-neoplastic and prostate cancer cells, where the extracts reduced H2O2-generated oxidative stress [64][61]. The pretreatment of non-malignant cell line HEK 293 was apoptosis-protective and cell death-reducing when H2O2-induced oxidative stress was applied. NFκB was activated in p53+/+ cells and suppressed in p53−/− cells. Leaf water and ethanol extracts were applied to a range of other human cancer cell lines [65][62]. Water extracts were reported not to be antiproliferative, while ethanol extracts were particularly effective to hepatocellular carcinoma (HepG2) and non-small cell lung adenocarcinoma (A549) cell lines and were found to exert significant antimigratory concentration-dependent effects in both cell lines.
Total methanol extracts and polar and apolar fractions were tested in a completely different system—important plant pathogens to search for potential sustainable and eco-friendly plant protection approaches [66][67][60,64]. No fungitoxic effect on Alternaria alternata and Fusarium oxysporum was found. Strong inhibition of Botrytis cinerea was achieved, in particular by apolar fractions. The same fractions stimulated the growth of Phytophthora citricola and can be potentially used as an effector. Other Phythophtora spp. isolates were stimulated significantly to grow under in vitro conditions, which could be a good prerequisite for the development of culture media for further tests on these obligate pathogens.
The significant amount of myconoside in plant extracts from European Gesneriads along with the strong scavenging activity gave ground for potential application in various human welfare areas. There is a significant interest in the application of myconoside as a single compound or in well-characterized combination with other secondary metabolites in various medicinal test systems.
An extract rich in myconoside isolated from H. rhodopensis was reported to increase mRNA synthesis of collagen and elastin genes in human dermal fibroblasts stressed with H2O2 [68][70]. The extracts were proven to protect against UV-induced dermis oxidation and even increased skin elasticity of human volunteers. It was suggested that H. rhodopensis extracts can be used for anti-aging treatments, protecting the skin from oxidation, increasing skin elasticity, and enhancing skin radiance. The anti-aging potential of such extracts was further confirmed. Influence on cell periphery, permeabilization of the membrane, and disruption of HaCaT keratinocyte tight junctions were observed—more pronounced in actively dividing cells [69][58]. In a very different experimental system, the strong and specific proliferative, anti-aging, and protective effect was found in the model yeast Saccharomyces cerevisiae cell line to revitalize and ameliorate cellular growth as well as to balance intracellular metabolic states [70][51]. Myconoside, isolated from H. rhodopensis extracts, was shown to have strong antioxidative potential along with a significant hepatoprotective effect on isolated rat hepatocytes [71][68]. H. rhodopensis extract fractions, containing myconoside or enriched with Calceolarioside E, were found to be very effective in promoting the expression of nuclear factor erythroid 2 p45-related factor 2 (Nrf2), a transcriptional regulator of the cellular redox balance and a suppressor of the pathological manifestation of various diseases, in bone marrow neutrophils [72][69].