Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Shigehiko Yumura | -- | 3418 | 2024-02-21 23:37:27 | | | |
| 2 | Sirius Huang | -1 word(s) | 3417 | 2024-02-22 03:54:15 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Yumura, S. Wound Repair of the Cell Membrane. Encyclopedia. Available online: https://encyclopedia.pub/entry/55328 (accessed on 24 June 2026).
Yumura S. Wound Repair of the Cell Membrane. Encyclopedia. Available at: https://encyclopedia.pub/entry/55328. Accessed June 24, 2026.
Yumura, Shigehiko. "Wound Repair of the Cell Membrane" Encyclopedia, https://encyclopedia.pub/entry/55328 (accessed June 24, 2026).
Yumura, S. (2024, February 21). Wound Repair of the Cell Membrane. In Encyclopedia. https://encyclopedia.pub/entry/55328
Yumura, Shigehiko. "Wound Repair of the Cell Membrane." Encyclopedia. Web. 21 February, 2024.
Copy Citation
The cell membrane is frequently subjected to damage, either through physical or chemical means. The swift restoration of the cell membrane’s integrity is crucial to prevent the leakage of intracellular materials and the uncontrolled influx of extracellular ions. Consequently, wound repair plays a vital role in cell survival, akin to the importance of DNA repair.
actin
cell membrane
calcium ion
membrane-remodeling
myosin
wound repair
1. Introduction
The cell membrane serves as a crucial barrier between the extracellular and intracellular spaces, yet it is consistently vulnerable to physical or chemical damage. Such injuries compromise the membrane’s integrity, leading to an influx of undesirable substances into the cell and cytoplasmic loss. Local wounds on the cell membrane also impact cell polarity during migration [1] and influence the division axis and symmetrical division in cell division [2]. Mechanically active tissues, like skeletal and cardiac muscles, frequently experience membrane wounds due to repeated contractions [3][4][5]. Ischemia-reperfusion injury followed by heart attack and stroke also damages the cell membrane [6]. Infection by pathogenic funguses, bacteria, and viruses and their pore-forming toxins can also result in membrane wounds at the cell membrane [7][8][9][10]. Loss of the wound repair function is observed in various diseases, including diabetes [11], muscular dystrophies [12][13], acute kidney injury [14], and vitamin deficiencies [15]. Recent studies have identified defects in wound repair as common in Parkinson’s and Alzheimer’s diseases [16][17][18]. Plant cells, affected by freezing damage in cold seasons, also possess the capability to repair damaged membranes [19][20][21]. Similar to DNA repair, wound repair is a physiologically vital phenomenon for living cells. Moreover, many methods for introducing extracellular substances into cells, such as microinjection and electroporation, rely on cellular wound repair. See also recent good reviews [6][22][23][24][25][26].
The mechanisms of wound repair have been extensively studied across various model organisms, including mammalian cells [27][28], amphibian oocytes [29][30][31][32], echinoderm oocytes [33][34][35], fruit flies [36][37][38], nematodes [12][39][40], amoebae [41][42], yeast [43][44], ciliate [45], plant cells [19][46][47], and Dictyostelium cells [48]. A common feature among these mechanisms is the essential role of Ca2+ influx from an external medium in the wound repair process. While the “membrane patch hypothesis” suggests that cytosolic membrane vesicles accumulate at the wound site to form an impermanent “patch” for emergency wound pore plugging [34][49][50][51], alternative hypotheses that do not involve patching have also been proposed [25][27][52]. However, there is no universally accepted general model for the mechanisms driving the repair process.
2. The Cell Can Repair a Wounded Cell Membrane
The presence of a cell membrane wound repair mechanism must be noticed during the initial phases of single-cell microsurgery and microinjection experiments [30][31][53][54][55][56]. For example, when a Dictyostelium cell is divided into two fragments using a microneedle, the nucleate fragment exhibits normal migration, whereas the anucleate fragment is incapable of doing so [57]. This experiment underscores the nucleus’s indispensable role in cell migration and simultaneously emphasizes the prompt repair of the cell membrane, which was wounded during microsurgery—a pioneering observation in Dictyostelium cells.
The majority of wound experiments have focused on a limited life stage of cells. The life cycle of Dictyostelium discoideum is broadly categorized into four stages: vegetative, aggregation, multicellular, and culmination. After the starvation of vegetative cells, individual cells aggregate to form streams towards the aggregation center. Aggregation is mediated by the chemotaxis of cells toward cAMP excreted from the aggregation centers. This process results in the formation of a multicellular organism and eventually leads to the development of fruiting bodies consisting of spores and stalks. Wound repair is observed at all stages in Dictyostelium cells (Figure 1A), including spore cells, which are dormant cells with a rigid cell wall. Furthermore, wound repair is noted at different stages of the cell cycle, such as interphase and the mitotic stage, in Dictyostelium cells [2].
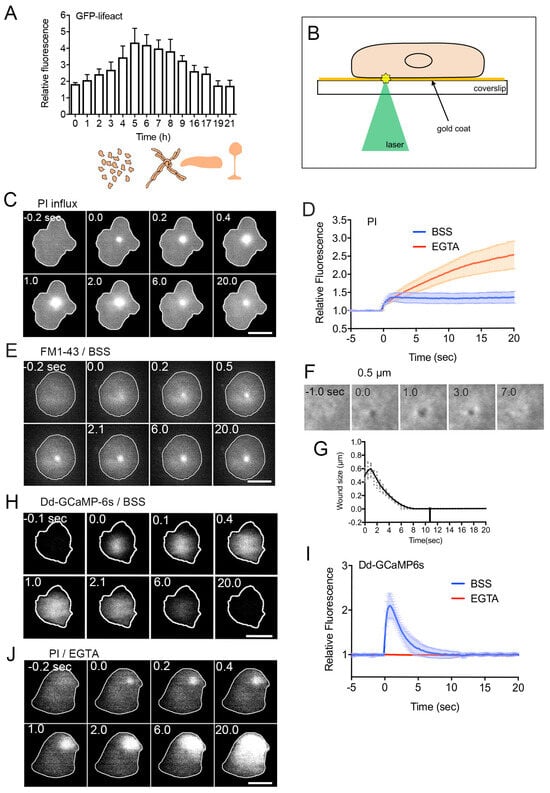
Figure 1. Wound repair of the cell membrane. (A) Relative amplitudes of actin accumulation at the wound site over time following starvation (0 h). As illustrated in the lower drawings, upon initiation of vegetative cell starvation, individual cells aggregate, forming streams direct toward the aggregation center. This process leads to the creation of a multicellular structure, culminating in the development of a fruiting body. Importantly, wound repair is observed at every stage of the lifecycle in Dictyostelium discoideum. (B) Schematic representation of the enhanced laserporation with gold coating. The wound diameter is usually set at 0.5 μm for Dictyostelium cells. (C) A representative sequence of fluorescence images capturing PI influx after laserporation. (D) Temporal profiles of PI influx in the presence (BSS, control) and absence (EGTA) of external Ca2+. The wound laser beam was applied at 0 sec. (E) A typical sequence of fluorescence images illustrating FM dye influx after laserporation. (F) Laserporation of a cell expressing GFP-cAR1 resulted in the appearance of a black spot on the cell membrane. The black spot transiently expanded, then contracted, and finally closed. (G) The time course of the black spot diameter. (H) A sequence of fluorescence images featuring a cell expressing GCAMP6s after laserporation. (I) Temporal profiles of GCAMP6s fluorescence intensities in the presence (BSS) and absence (EGTA) of external Ca2+. (J) A typical sequence of fluorescence images illustrating PI influx after laserporation in the absence (EGTA) of external Ca2+. Scale bars, 10 µm. Figures are posted from [48][51][58][59] with CC BY licence.
3. Monitoring of Wound Repair
Various methods have been employed to investigate the wound repair mechanism. In early experiments, the cell membrane was wounded mainly by microneedle poking in large cells such as protozoan amebae [41][56][60], amphibian eggs [30][53][55], and echinoderm eggs [50]. For small cells like yeast, animal cultured cells, and Dictyostelium cells, laser ablation has been predominantly used due to the technical challenges and time constraints associated with microneedle poking in such small cells. Recent research also utilizes laser ablation for large cells, offering precise-sized wounds and accurate timing. However, both laser ablation and previous methods not only damage the cell membrane but also impact intracellular structures, including cortical actin networks, microtubules, and organelles.
To address this, researchers have developed an improved laser ablation method that selectively injures only the cell membrane. As depicted in Figure 1B, after placing cells on a carbon or gold-coated coverslip, a laser beam is focused on the coat underneath the cells. The laser energy absorbed by the coat generates heat and/or plasmon [48][61], selectively injuring the cell membrane attached to the coat [48]. This method has been originally invented for the introduction of extracellular substances into cells [62]. Instead of wounding individual cells, for biochemical analysis, a large number of cells can be wounded by treating with pore-forming agents or detergents [63][64][65].
For monitoring the wounding process, propidium iodide (PI) or FM1-43 has been widely used. PI, a cell-impermeant dye emitting fluorescence upon binding to RNA or DNA, and FM1-43, a cell-impermeable fluorescent lipid analog emitting fluorescence upon insertion into the membrane, are placed in the external medium. Their entry into the cytosol is monitored by the increase in fluorescence upon wounding. As shown in Figure 1C, PI fluorescence begins to increase at the wound site upon injury, spreading over the cytosol, suggesting PI entry through the wound pores. Figure 1D (BSS) illustrates the time course of PI fluorescence intensity in the cytosol of wounded cells, indicating that PI influx ceases within 2–3 s after injury, terminating urgent wound repair within this timeframe. Figure 1E demonstrates the influx of FM1-43 dye upon wounding, also showing that the dye enters from the wound pore and spreads across the cytoplasm.
To visualize the wound pore in Dictyostelium cells, cells expressing GFP-cAR1 (cAMP receptor) as a membrane protein marker are wounded. Immediately after wounding, a black spot appears at the laser application site (Figure 1F). This black spot is not generated by photobleaching, as it transiently expands slightly, then shrinks, and eventually closes (Figure 1G). This closure is not uniform but occurs from the wound edge to the center.
4. Ca2+ Influx as the First Signal
The initial signal common to all examined cells across various species is the influx of Ca2+ from the wound pore [30][32][41][50][66]. Monitoring this influx is feasible using a Ca2+ indicating fluorescent dye or a GFP-based Ca2+ indicator. Figure 1H presents a time series of fluorescence images of Dictyostelium cells expressing GCAMP6s, a GFP-based fluorescent Ca2+ indicator. In Figure 1I (BSS), the time course of fluorescence intensities in the cytosol is depicted. Intracellular Ca2+ concentration (Cai2+) promptly rises upon wounding, returning to resting levels within approximately 7 s. In the absence of external Ca2+, Cai2+ remains unchanged upon wounding (Figure 1I, EGTA), indicating that the influx of Ca2+ triggers the increase in Cai2+. Without external Ca2+, PI influx persists, leading to the eventual death of wounded cells (Figure 1D, EGTA, and Figure 1J). Additionally, the black spot observed in experiments using GFP-cAR1 does not close without Ca2+ influx. A concentration higher than 0.1 mM of Ca2+ in the external medium is necessary for wound repair in Dictyostelium cells [51].
Ca2+ influx induces the release of Ca2+ from intracellular stores through the calcium-induced calcium release (CICR) mechanism [67][68]. Deleting CICR reduces the amplitude of Cai2+ but does not impact wound repair, indicating that a local increase in Cai2+ is crucial, not a global one [58]. On the other hand, MCOLN1, an endosomal and lysosomal Ca2+-channel, is crucial for cell membrane repair in muscle cells, emphasizing the significance of Ca2+ release from intracellular stores in wound repair [69].
Various intracellular targets of Ca2+ for wound repair include dysfelin, mitsugmin 53 (MG53), neuroblast differentiation-associated protein (AHNAK), calpain, calmodulin, annexins, the endosomal sorting complex required for transport (ESCRT), protein kinase C, and actin-related proteins. These will be discussed in detail later.
5. Closing Wound Pores
5.1. Spontaneous Self-Sealing
Ruptured artificial lipid bilayer membranes exhibit spontaneous resealing [70]. Similarly, extremely small wound pores, such as those generated by electroporation in live cells at the nanometer scale, are thought to undergo spontaneous resealing due to thermodynamically unfavorable lipid disorder (Figure 2A). However, even with such small pores, there may be a necessity for an active wound repair mechanism [71]. Additionally, membrane pores created by pore-forming toxins, despite being nanometer-sized, require an active wound repair mechanism [7][23][72].
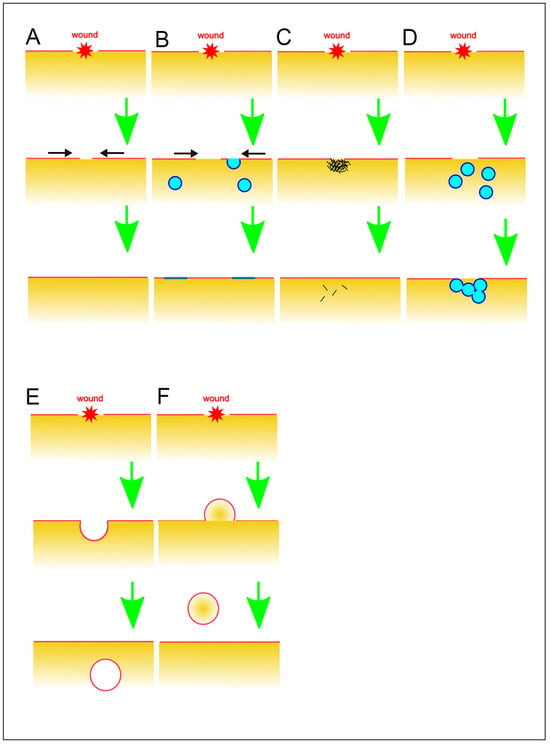
Figure 2. Various models for wound repair mechanisms. (A) Spontaneous self-sealing. (B) Self-sealing by regulation of surface tension. Black arrows indicate the direction of the membrane flow. (C) Sealing by protein aggregation. (D) Sealing by membrane patch. (E) Endocytosis of damaged membrane. (F) Vesicle budding and shedding to the outside. These illustrations are simplified for a better understanding of basic concepts.
5.2. Self-Sealing by Regulation of Surface Tension
Given the tension on the cell surface, larger pores cannot spontaneously reseal against the cell surface tension. The wound-induced influx of Ca2+ triggers the fusion of exocytic vesicles with the cell membrane, extending beyond the wound site and enlarging the plasma membrane (Figure 2B). This process results in a reduction of cell surface tension, facilitating spontaneous resealing and closure of the wounded pore [73]. In large cells, although the actomyosin ring exerts force to close the wound pore against the opening force of the cell surface tension, self-sealing alone is insufficient, and a membrane patch is also required for sealing, as described later.
5.3. Sealing by Protein Aggregation
The wounded pores have been suggested to be clogged by the aggregation of proteins, including annexins and actin (Figure 2C). Annexin A5 self-assembles into two-dimensional arrays on the membrane upon Ca2+ activation, a crucial aspect of its role in plasma membrane repair in mammalian cells [74]. Similar clogging phenomena have been reported for other annexins, which collaborate with wound repair-related proteins such as actin, dysferin, and MG53 [75][76][77][78]. Actin accumulates at the wound site, potentially serving a clogging function through actin gelation. However, this accumulation does not happen immediately upon wounding but occurs after the cessation of PI influx.
5.4. Sealing by Membrane Patch
Mammalian red blood cells, lacking endomembranes, including nuclei, take a longer time to reseal wound membranes or fail to repair in physiological conditions, suggesting that endomembranes are necessary for wound repair [79]. The membrane patch hypothesis was initially proposed in large cells like echinoderm and frog oocytes (Figure 2D). A local increase in Ca2+ induces the fusion of small cytoplasmic vesicles with each other, creating a continuous membrane plug at the wound site along with the plasma membrane [34][35]. More recently, cortical granules in Xenopus oocytes have been identified as such intracellular compartments, and their fusion was visualized in live cells [80][81]. This wound repair process is succeeded by the constriction of an actomyosin ring, akin to the contractile ring involved in cytokinesis.
Various sources for the membrane patch, including lysosomes [82][83], endosomes [84][85], MG53-rich vesicles [66], dysferlin-containing vesicles [86][87], or AHNAK-positive “enlargeosomes” [88][89], have been proposed. However, these vesicles and organelles might not meet the spatiotemporal requirements for rapid and efficient wound repair, considering the possibility of multiple wounds with very short intervals [58].
Recently, researchers proposed that the vesicles for the membrane plug are newly generated at the wound site in Dictyostelium cells [51]. In influx experiments using FM dye, most of the FM fluorescence diffuses in the cytosol, but a portion of FM dye remains at the wounded site and increases in size (Figure 1E), indicating membrane accumulation at the wound site. In the PI influx experiment (Figure 1C), a portion of PI fluorescence also persists at the wound site, suggesting that cytoplasm, including PI dye, is entrapped in the newly enclosed vesicles. It is improbable that the membrane plug originates from the broken cell membrane due to the limited amount of the broken cell membrane. Additionally, vesicles are unlikely to be transported from other locations since pharmacological disruption of microtubules and actin did not impede membrane accumulation. Therefore, researchers propose that the vesicles for the membrane plug are generated de novo at the wound site, although the mechanism for this generation remains unclear.
As described earlier, experiments using GFP-cAR1 show that the wound pore is not repaired from the wound edge to the center. Therefore, the wound edge grows toward the center through vesicles repeatedly fusing with the edge of the cell membrane, rather than forming a fused large patch to plug the wound pore.
5.5. Endocytosis of Damaged Membrane
Ca2+-triggered endocytosis is suggested to eliminate damaged membrane (Figure 2E). The membrane, including the damaged portion, invaginates inward, and the resulting bud is removed by releasing it into the cell, dependent on Ca2+ [65][90]. Upon wounding, acid sphingomyelinase is secreted into the extracellular space through Ca2+-dependent lysosomal exocytosis. This enzyme hydrolyzes sphingomyelin in the cell membrane into ceramide, facilitating membrane invagination and vesiculation [91]. Ceramide formation by sphingomyelinase also induces caveolae-mediated endocytosis, internalizing the wounded membrane [90][92][93][94][95][96]. It has been also reported that clathrin- and dynamin-mediated endocytosis facilitates removing the wounds by pore-forming proteins or toxins [97][98].
In Dictyostelium cells, neither endocytosis nor exocytosis appears to contribute to membrane accumulation for wound repair [51], despite the rapid turnover of the cell membrane through endocytosis–exocytosis coupling [99][100][101][102]. Notably, caveolin proteins are not present in Dictyostelium [103] and inhibitors of sphingomyelinase do not affect the wound repair in Dictyostelium cells (our preliminary observations). Additionally, clathrin- and dynamin-mediated endocytosis does not contribute to wound repair in Dictyostelium cells [48].
5.6. Vesicle Budding and Shedding to the Outside
Rather than endocytosis involving the inward budding and scission of the damaged membrane, vesicle budding or blebbing toward the outside of the cell, followed by scission, facilitates the removal and shedding of damaged membrane or pore-forming reagents (Figure 2F). Membrane-binding proteins for wound repair, such as the endosomal sorting complex required for transport (ESCRT) and annexins, facilitate this type of shedding in a Ca2+-dependent manner in mammal cells [10][44][75][90][104][105].
In Dictyostelium cells, FM dye that accumulates at the wound site remains there for the duration of observation. The wounded sites do not move relative to the substrate and eventually shed onto the substrate as cells migrate [1].
6. Membrane-Binding Proteins in Wound Repair
For wound repair, various membrane-binding proteins, including annexins, the ESCRT complex, synaptotagmin, and dysferin, have been proposed. These proteins also act as sensors, detecting damage to the cell membrane due to their calcium-dependency [106].
6.1. Annexins
Annexins, a highly conserved and ubiquitous family of Ca2+- and phospholipid-binding proteins, play a crucial role in wound repair [107][108][109][110][111][112]. In vertebrates, 12 annexin subfamilies (A1–A11 and A13) have been identified. Annexins such as A1, A2, A5, and A6 accumulate at the wound site by binding to the inner cell membrane, particularly acidic phospholipids like phosphatidylserine, in response to a Ca2+ influx. Through membrane binding and interactions with other proteins, such as S100 family proteins, annexin prevents further expansion of the wound pore, reduces membrane tension, and prepares the membrane for resealing [113][114][115].
Annexins induce curvature in the free-edge membranes and generate constriction force to close the wound pore through annexin crosslinking [116][117][118]. Annexins can also be cross-linked by transglutamilases in a Ca2+-dependent manner [119], which have also been implicated in plasma membrane repair [120].
Moreover, annexins have been proposed to assemble into multimeric lattice structures, recruiting M53-laden vesicles and mini-dysferlin72, effectively clogging the wound pore [13][74][111][121]. Some annexins, such as annexin 1 and 2, can bind to actin and stabilize actin filaments at the wound site [76][110][122][123][124].
Dictyostelium possesses two annexin genes, annexin C1 (annexin VII or synexin) and annexin C2 (annexin I) [125]. Both can bind phosphatidylserine in a Ca2+-dependent manner [126][127]. Only annexin C1 accumulates at the wound site immediately after wounding. Wounded annexin C1-null cells exhibit irregular curves with multiple peaks in PI influx, Ca2+ influx, and actin dynamics. Additionally, annexin C1-null cells have a significantly reduced survival rate following injury, suggesting that annexin C1 partially contributes to wound repair in Dictyostelium cells [48][58].
6.2. ESCRT Complexes
The endosomal sorting complex required for transport (ESCRT) is categorized into five protein complexes (ESCRT-0, ESCRT-I, ESCRT-II, ESCRT-III, and Vps4). These complexes play integral roles in various cellular processes, including endosomal budding transport, virus budding, and cytokinesis. The ESCRT complex constricts and severs narrow necks during membrane budding processes [44][128][129][130][131][132][133][134][135][136]. Additionally, ESCRT complexes have been implicated in shedding wounded membranes as extracellular vesicles [44][105][136]. Upon injury, ESCRT complexes promptly accumulate at the wound site, protrude the wounded membrane as a bud or bleb, and subsequently cut it off to release extracellular vesicles. This ESCRT-mediated abscission of the wounded membrane appears to limit smaller-sized wounds (<100 nm in diameter) [137][138].
Interestingly, ESCRT complexes also participate in repairing damaged membranes of intracellular organelles, such as the nuclear envelope and lysosomes [139][140][141]. Furthermore, ESCRT complexes mediate the sealing of holes in the nascent nuclear envelope and nascent autophagosome [142].
While ESCRT complexes themselves are not sensitive to Ca2+, Ca2+-sensitive proteins like ALG-2 and calmodulin confer Ca2+ sensitivity on ESCRT complexes [10][136][143]. Recently, it has been reported that annexin A6 also plays a similar role in the secretion of exosomes [144].
In Dictyostelium cells, components of the ESCRT complexes accumulate at the wound site immediately upon injury, depending on the influx of Ca2+ [51]. However, in ESCRT null cells, PI influx ceases normally, and actin dynamics are observed, similar to wild-type cells. This suggests that ESCRT complexes are not essential for wound repair in Dictyostelium cells [48].
During cytokinesis in animal cells, ESCRT complexes and annexins accumulate at the cleavage furrow and/or midbody and are considered to play a role in cytokinesis [145][146][147][148][149][150]. Dictyostelium cells lack a midbody and undergo division through physical cutting via the constriction of the contractile ring and the traction force of the two daughter fragments migrating in opposite directions [151][152]. This suggests that this abscission might be a form of ‘physiological wound’. However, preliminary observations indicate that neither ESCRT components nor annexins accumulate at the torn edges, suggesting the existence of a novel mechanism for cytokinetic abscission in Dictyostelium cells.
6.3. Synaptotagmin
Synaptotagmin comprises a family of Ca2+-binding and membrane-trafficking proteins, with particular emphasis on its well-characterized role in the release of synaptic vesicles in neurons, where it regulates Ca2+-dependent exocytosis [153]. Synaptotagmin 7, specifically, participates in the repair of wounded membranes through Ca2+-dependent lysosome exocytosis [83][154]. Knockdown mice lacking synaptotagmin 7 exhibit defects in wound repair [155]. While Dictyostelium possesses a synaptotagmin-like protein, there is currently no available information regarding its role in wound repair.
6.4. Dysferlin
Dysferlin, a membrane protein within the Ferlin family involved in vesicle fusion, is notably abundant in skeletal and cardiac muscle [156]. Dysferlin binds to vesicles containing acid phospholipids, such as phosphatidylserine, through the dysferlin C2 domain, relying on Ca2+ [157]. Mice deficient in dysferlin display defects in wound repair in muscle cells and develop muscular dystrophy [12]. Dysferlin is proposed to generate a membrane patch by recruiting and fusing vesicles in wounded skeletal and cardiac muscle cells [87][158]. Dysferlin organizes vesicle fusion with the assistance of binding partners like S100A10, annexin A2, AHNAK, caveolin-3, and TRIM72 (MG53) [13][66][93][106][159]. TRIM72, also known as MG53, which is highly expressed in muscle cells, assembles into a higher-ordered structure on the phosphatidylserine-enriched membranes. This assembly and association with the membrane depend on its oligomeric assembly and ubiquitination activity, facilitating vesicle transport to the wound site [160]. Notably, Dictyostelium cells lack a dysferlin homolog.
References
- Pervin, M.S.; Yumura, S. Manipulation of cell migration by laserporation-induced local wounding. Sci. Rep. 2019, 9, 4291.
- Tanvir, M.I.O.; Yumura, S. Effects of wounds in the cell membrane on cell division. Sci. Rep. 2023, 13, 1941.
- McNeil, P.L.; Ito, S. Gastrointestinal cell plasma membrane wounding and resealing in vivo. Gastroenterology 1989, 96, 1238–1248.
- McNeil, P.L.; Khakee, R. Disruptions of muscle fiber plasma membranes. Role in exercise-induced damage. Am. J. Pathol. 1992, 140, 1097–1109.
- McNeil, P.L. Cellular and molecular adaptations to injurious mechanical stress. Trends Cell Biol. 1993, 3, 302–307.
- Dias, C.; Nylandsted, J. Plasma membrane integrity in health and disease: Significance and therapeutic potential. Cell Discov. 2021, 7, 4.
- Etxaniz, A.; Gonzalez-Bullon, D.; Martin, C.; Ostolaza, H. Membrane Repair Mechanisms against Permeabilization by Pore-Forming Toxins. Toxins 2018, 10, 234.
- Luisoni, S.; Suomalainen, M.; Boucke, K.; Tanner, L.B.; Wenk, M.R.; Guan, X.L.; Grzybek, M.; Coskun, U.; Greber, U.F. Co-option of Membrane Wounding Enables Virus Penetration into Cells. Cell Host Microbe 2015, 18, 75–85.
- Stow, J.L.; Condon, N.D. The cell surface environment for pathogen recognition and entry. Clin. Trans. Immunol. 2016, 5, e71.
- Westman, J.; Plumb, J.; Licht, A.; Yang, M.; Allert, S.; Naglik, J.R.; Hube, B.; Grinstein, S.; Maxson, M.E. Calcium-dependent ESCRT recruitment and lysosome exocytosis maintain epithelial integrity during Candida albicans invasion. Cell Rep. 2022, 38, 110187.
- Howard, A.C.; McNeil, A.K.; Xiong, F.; Xiong, W.C.; McNeil, P.L. A novel cellular defect in diabetes: Membrane repair failure. Diabetes 2011, 60, 3034–3043.
- Bansal, D.; Miyake, K.; Vogel, S.S.; Groh, S.; Chen, C.C.; Williamson, R.; McNeil, P.L.; Campbell, K.P. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature 2003, 423, 168–172.
- Waddell, L.B.; Lemckert, F.A.; Zheng, X.F.; Tran, J.; Evesson, F.J.; Hawkes, J.M.; Lek, A.; Street, N.E.; Lin, P.; Clarke, N.F.; et al. Dysferlin, annexin A1, and mitsugumin 53 are upregulated in muscular dystrophy and localize to longitudinal tubules of the T-system with stretch. J. Neuropathol. Exp. Neurol. 2011, 70, 302–313.
- Duann, P.; Li, H.; Lin, P.; Tan, T.; Wang, Z.; Chen, K.; Zhou, X.; Gumpper, K.; Zhu, H.; Ludwig, T.; et al. MG53-mediated cell membrane repair protects against acute kidney injury. Sci. Transl. Med. 2015, 7, 279ra36.
- Labazi, M.; McNeil, A.K.; Kurtz, T.; Lee, T.C.; Pegg, R.B.; Angeli, J.P.F.; Conrad, M.; McNeil, P.L. The antioxidant requirement for plasma membrane repair in skeletal muscle. Free. Radic. Biol. Med. 2015, 84, 246–253.
- Agnihotri, A.; Aruoma, O.I. Alzheimer’s Disease and Parkinson’s Disease: A Nutritional Toxicology Perspective of the Impact of Oxidative Stress, Mitochondrial Dysfunction, Nutrigenomics and Environmental Chemicals. J. Am. Coll. Nutr. 2020, 39, 16–27.
- Bulgart, H.R.; Goncalves, I.; Weisleder, N. Leveraging Plasma Membrane Repair Therapeutics for Treating Neurodegenerative Diseases. Cells 2023, 12, 1660.
- Maiti, P.; Manna, J.; Dunbar, G.L. Current understanding of the molecular mechanisms in Parkinson’s disease: Targets for potential treatments. Transl. Neurodegener. 2017, 6, 28.
- Schapire, A.L.; Valpuesta, V.; Botella, M.A. Plasma membrane repair in plants. Trends Plant Sci. 2009, 14, 645–652.
- Yamazaki, T.; Takata, N.; Uemura, M.; Kawamura, Y. Arabidopsis synaptotagmin SYT1, a type I signal-anchor protein, requires tandem C2 domains for delivery to the plasma membrane. J. Biol. Chem. 2010, 285, 23165–23176.
- Vyse, K.; Penzlin, J.; Sergeant, K.; Hincha, D.K.; Arora, R.; Zuther, E. Repair of sub-lethal freezing damage in leaves of Arabidopsis thaliana. BMC Plant Biol. 2020, 20, 35.
- Ammendolia, D.A.; Bement, W.M.; Brumell, J.H. Plasma membrane integrity: Implications for health and disease. BMC Biol. 2021, 19, 71.
- Barisch, C.; Holthuis, J.C.M.; Cosentino, K. Membrane damage and repair: A thin line between life and death. Biol. Chem. 2023, 404, 467–490.
- Horn, A.; Jaiswal, J.K. Structural and signaling role of lipids in plasma membrane repair. Curr. Top. Membr. 2019, 84, 67–98.
- Nakamura, M.; Dominguez, A.N.M.; Decker, J.R.; Hull, A.J.; Verboon, J.M.; Parkhurst, S.M. Into the breach: How cells cope with wounds. Open Biol. 2018, 8, 180135.
- Xu, S.; Yang, T.J.; Xu, S.; Gong, Y.N. Plasma membrane repair empowers the necrotic survivors as innate immune modulators. Semin. Cell Dev. Biol. 2023, 156, 93–106.
- Andrews, N.W.; Corrotte, M. Plasma membrane repair. Curr. Biol. 2018, 28, R392–R397.
- Togo, T.; Alderton, J.M.; Bi, G.Q.; Steinhardt, R.A. The mechanism of facilitated cell membrane resealing. J. Cell Sci. 1999, 112, 719–731.
- Bement, W.M.; Capco, D.G. Analysis of inducible contractile rings suggests a role for protein kinase C in embryonic cytokinesis and wound healing. Cell Motil. Cytoskelet. 1991, 20, 145–157.
- Gingell, D. Contractile responses at the surface of an amphibian egg. J. Embryol. Exp. Morphol. 1970, 23, 583–609.
- Holtfreter, J. Properties and functions of the surface coat in amphibian embryos. J. Exp. Zool. 1943, 93, 251–323.
- Stanisstreet, M. Calcium and wound healing in Xenopus early embryos. J. Embryol. Exp. Morphol. 1982, 67, 195–205.
- Bi, G.Q.; Alderton, J.M.; Steinhardt, R.A. Calcium-regulated exocytosis is required for cell membrane resealing. J. Cell Biol. 1995, 131, 1747–1758.
- McNeil, P.L.; Vogel, S.S.; Miyake, K.; Terasaki, M. Patching plasma membrane disruptions with cytoplasmic membrane. J. Cell Sci. 2000, 113, 1891–1902.
- Terasaki, M.; Miyake, K.; McNeil, P.L. Large plasma membrane disruptions are rapidly resealed by Ca2+-dependent vesicle-vesicle fusion events. J. Cell Biol. 1997, 139, 63–74.
- Abreu-Blanco, M.T.; Verboon, J.M.; Parkhurst, S.M. Cell wound repair in Drosophila occurs through three distinct phases of membrane and cytoskeletal remodeling. J. Cell. Biol. 2011, 193, 455–464.
- Abreu-Blanco, M.T.; Verboon, J.M.; Parkhurst, S.M. Coordination of Rho family GTPase activities to orchestrate cytoskeleton responses during cell wound repair. Curr. Biol. 2014, 24, 144–155.
- Razzell, W.; Wood, W.; Martin, P. Swatting flies: Modelling wound healing and inflammation in Drosophila. Dis. Model. Mech. 2011, 4, 569–574.
- Ma, Y.; Xie, J.; Wijaya, C.S.; Xu, S. From wound response to repair—Lessons from C. elegans. Cell Regen. 2021, 10, 5.
- Xu, S.; Chisholm, A.D. C. elegans epidermal wounding induces a mitochondrial ROS burst that promotes wound repair. Dev. Cell 2014, 31, 48–60.
- Jeon, K.W.; Jeon, M.S. Cytoplasmic filaments and cellular wound healing in Amoeba proteus. J. Cell Biol. 1975, 67, 243–249.
- Szubinska, B. Closure of the plasma membrane around microneedle in Amoeba proteus. An ultrastructural study. Exp. Cell Res. 1978, 111, 105–115.
- Kono, K.; Saeki, Y.; Yoshida, S.; Tanaka, K.; Pellman, D. Proteasomal degradation resolves competition between cell polarization and cellular wound healing. Cell 2012, 150, 151–164.
- Radulovic, M.; Stenmark, H. ESCRTs in membrane sealing. Biochem. Soc. Trans. 2018, 46, 773–778.
- Zhang, K.S.; Blauch, L.R.; Huang, W.; Marshall, W.F.; Tang, S.K.Y. Microfluidic guillotine reveals multiple timescales and mechanical modes of wound response in Stentor coeruleus. BMC Biol. 2021, 19, 63.
- Foissner, I.; Wasteneys, G.O. The characean internodal cell as a model system for studying wound healing. J. Microsc. 2012, 247, 10–22.
- Schapire, A.L.; Voigt, B.; Jasik, J.; Rosado, A.; Lopez-Cobollo, R.; Menzel, D.; Salinas, J.; Mancuso, S.; Valpuesta, V.; Baluska, F.; et al. Arabidopsis synaptotagmin 1 is required for the maintenance of plasma membrane integrity and cell viability. Plant Cell 2008, 20, 3374–3388.
- Yumura, S.; Talukder, M.S.U.; Pervin, M.S.; Tanvir, M.I.O.; Matsumura, T.; Fujimoto, K.; Tanaka, M.; Itoh, G. Dynamics of Actin Cytoskeleton and Their Signaling Pathways during Cellular Wound Repair. Cells 2022, 11, 3166.
- McNeil, P.L.; Terasaki, M. Coping with the inevitable: How cells repair a torn surface membrane. Nat. Cell Biol. 2001, 3, E124–E129.
- Steinhardt, R.A.; Bi, G.; Alderton, J.M. Cell membrane resealing by a vesicular mechanism similar to neurotransmitter release. Science 1994, 263, 390–393.
- Talukder, M.S.U.; Pervin, M.S.; Tanvir, M.I.O.; Fujimoto, K.; Tanaka, M.; Itoh, G.; Yumura, S. Ca2+-calmodulin dependent wound repair in Dictyostelium cell membrane. Cells 2020, 9, 1058.
- Moe, A.M.; Golding, A.E.; Bement, W.M. Cell healing: Calcium, repair and regeneration. Semin. Cell Dev. Biol. 2015, 45, 18–23.
- Bluemink, J.G. Cortical wound healing in the amphibian egg: An electron microscopical study. J. Ultrastruct. Res. 1972, 41, 95–114.
- Jeon, K.W.; Danielli, J.F. Micrurgical studies with large free-living amebas. Int. Rev. Cytol. 1971, 30, 49–89.
- Luckenbill, L.M. Dense material associated with wound closure in the axolotl egg (A. mexicanum). Exp. Cell Res. 1971, 66, 263–267.
- Szubinska, B. “New membrane” formation in Amoeba proteus upon injury of individual cells. Electron microscope observations. J. Cell Biol. 1971, 49, 747–772.
- Swanson, J.A.; Taylor, D.L. Local and spatially coordinated movements in Dictyostelium discoideum amoebae during chemotaxis. Cell 1982, 28, 225–232.
- Pervin, M.S.; Itoh, G.; Talukder, M.S.U.; Fujimoto, K.; Morimoto, Y.V.; Tanaka, M.; Ueda, M.; Yumura, S. A study of wound repair in Dictyostelium cells by using novel laserporation. Sci. Rep. 2018, 8, 7969.
- Tanvir, M.I.O.; Itoh, G.; Adachi, H.; Yumura, S. Dynamics of Myosin II Filaments during Wound Repair in Dividing Cells. Cells 2021, 10, 1229.
- Taylor, D.L.; Wang, Y.L.; Heiple, J.M. Contractile basis of ameboid movement. VII. The distribution of fluorescently labeled actin in living amebas. J. Cell Biol. 1980, 86, 590–598.
- Nakatoh, T.; Osaki, T.; Tanimoto, S.; Jahan, M.G.S.; Kawakami, T.; Chihara, K.; Sakai, N.; Yumura, S. Cell behaviors within a confined adhesive area fabricated using novel micropatterning methods. PLoS ONE 2022, 17, e0262632.
- Yumura, S. A novel low-power laser-mediated transfer of foreign molecules into cells. Sci. Rep. 2016, 6, 22055.
- Bischofberger, M.; Gonzalez, M.R.; van der Goot, F.G. Membrane injury by pore-forming proteins. Curr. Opin. Cell Biol. 2009, 21, 589–595.
- Calvello, R.; Mitolo, V.; Acquafredda, A.; Cianciulli, A.; Panaro, M.A. Plasma membrane damage sensing and repairing. Role of heterotrimeric G-proteins and the cytoskeleton. Toxicol. Vitr. 2011, 25, 1067–1074.
- Idone, V.; Tam, C.; Goss, J.W.; Toomre, D.; Pypaert, M.; Andrews, N.W. Repair of injured plasma membrane by rapid Ca2+-dependent endocytosis. J. Cell Biol. 2008, 180, 905–914.
- Cai, C.; Masumiya, H.; Weisleder, N.; Matsuda, N.; Nishi, M.; Hwang, M.; Ko, J.K.; Lin, P.; Thornton, A.; Zhao, X.; et al. MG53 nucleates assembly of cell membrane repair machinery. Nat. Cell Biol. 2009, 11, 56–64.
- Fisher, P.R.; Wilczynska, Z. Contribution of endoplasmic reticulum to Ca(2+) signals in Dictyostelium depends on extracellular Ca(2+). FEMS Microbiol. Lett. 2006, 257, 268–277.
- Malchow, D.; Lusche, D.F.; De Lozanne, A.; Schlatterer, C. A fast Ca2+-induced Ca2+-release mechanism in Dictyostelium discoideum. Cell Calcium 2008, 43, 521–530.
- Cheng, X.; Zhang, X.; Gao, Q.; Ali Samie, M.; Azar, M.; Tsang, W.L.; Dong, L.; Sahoo, N.; Li, X.; Zhuo, Y.; et al. The intracellular Ca(2)(+) channel MCOLN1 is required for sarcolemma repair to prevent muscular dystrophy. Nat. Med. 2014, 20, 1187–1192.
- Gozen, I.; Dommersnes, P. Pore dynamics in lipid membranes. Eur. Phys. J. Spec. Top. 2014, 223, 1813–1829.
- Hai, A.; Spira, M.E. On-chip electroporation, membrane repair dynamics and transient in-cell recordings by arrays of gold mushroom-shaped microelectrodes. Lab. Chip 2012, 12, 2865–2873.
- Lata, K.; Singh, M.; Chatterjee, S.; Chattopadhyay, K. Membrane Dynamics and Remodelling in Response to the Action of the Membrane-Damaging Pore-Forming Toxins. J. Membr. Biol. 2022, 255, 161–173.
- Togo, T.; Krasieva, T.B.; Steinhardt, R.A. A decrease in membrane tension precedes successful cell-membrane repair. Mol. Biol. Cell 2000, 11, 4339–4346.
- Bouter, A.; Gounou, C.; Berat, R.; Tan, S.; Gallois, B.; Granier, T.; d’Estaintot, B.L.; Poschl, E.; Brachvogel, B.; Brisson, A.R. Annexin-A5 assembled into two-dimensional arrays promotes cell membrane repair. Nat. Commun. 2011, 2, 270.
- Babiychuk, E.B.; Monastyrskaya, K.; Potez, S.; Draeger, A. Blebbing confers resistance against cell lysis. Cell Death Differ. 2011, 18, 80–89.
- Demonbreun, A.R.; Quattrocelli, M.; Barefield, D.Y.; Allen, M.V.; Swanson, K.E.; McNally, E.M. An actin-dependent annexin complex mediates plasma membrane repair in muscle. J. Cell Biol. 2016, 213, 705–718.
- Roostalu, U.; Strahle, U. In vivo imaging of molecular interactions at damaged sarcolemma. Dev. Cell 2012, 22, 515–529.
- Swaggart, K.A.; Demonbreun, A.R.; Vo, A.H.; Swanson, K.E.; Kim, E.Y.; Fahrenbach, J.P.; Holley-Cuthrell, J.; Eskin, A.; Chen, Z.; Squire, K.; et al. Annexin A6 modifies muscular dystrophy by mediating sarcolemmal repair. Proc. Natl. Acad. Sci. USA 2014, 111, 6004–6009.
- McNeil, P.L.; Miyake, K.; Vogel, S.S. The endomembrane requirement for cell surface repair. Proc. Natl. Acad. Sci. USA 2003, 100, 4592–4597.
- Davenport, N.R.; Bement, W.M. Cell repair: Revisiting the patch hypothesis. Commun. Integr. Biol. 2016, 9, e1253643.
- Davenport, N.R.; Sonnemann, K.J.; Eliceiri, K.W.; Bement, W.M. Membrane dynamics during cellular wound repair. Mol. Biol. Cell 2016, 27, 2272–2285.
- McNeil, P.L. Repairing a torn cell surface: Make way, lysosomes to the rescue. J. Cell Sci. 2002, 115, 873–879.
- Reddy, A.; Caler, E.V.; Andrews, N.W. Plasma membrane repair is mediated by Ca2+-regulated exocytosis of lysosomes. Cell 2001, 106, 157–169.
- Eddleman, C.S.; Ballinger, M.L.; Smyers, M.E.; Fishman, H.M.; Bittner, G.D. Endocytotic formation of vesicles and other membranous structures induced by Ca2+ and axolemmal injury. J. Neurosci. 1998, 18, 4029–4041.
- Raj, N.; Greune, L.; Kahms, M.; Mildner, K.; Franzkoch, R.; Psathaki, O.E.; Zobel, T.; Zeuschner, D.; Klingauf, J.; Gerke, V. Early Endosomes Act as Local Exocytosis Hubs to Repair Endothelial Membrane Damage. Adv. Sci. 2023, 10, e2300244.
- Lek, A.; Evesson, F.J.; Lemckert, F.A.; Redpath, G.M.; Lueders, A.K.; Turnbull, L.; Whitchurch, C.B.; North, K.N.; Cooper, S.T. Calpains, cleaved mini-dysferlinC72, and L-type channels underpin calcium-dependent muscle membrane repair. J. Neurosci. 2013, 33, 5085–5094.
- McDade, J.R.; Michele, D.E. Membrane damage-induced vesicle-vesicle fusion of dysferlin-containing vesicles in muscle cells requires microtubules and kinesin. Hum. Mol. Genet. 2014, 23, 1677–1686.
- Cocucci, E.; Racchetti, G.; Podini, P.; Rupnik, M.; Meldolesi, J. Enlargeosome, an exocytic vesicle resistant to nonionic detergents, undergoes endocytosis via a nonacidic route. Mol. Biol. Cell 2004, 15, 5356–5368.
- Meldolesi, J. Surface wound healing: A new, general function of eukaryotic cells. J. Cell Mol. Med. 2003, 7, 197–203.
- Andrews, N.W.; Almeida, P.E.; Corrotte, M. Damage control: Cellular mechanisms of plasma membrane repair. Trends Cell Biol. 2014, 24, 734–742.
- Holopainen, J.M.; Angelova, M.I.; Kinnunen, P.K. Vectorial budding of vesicles by asymmetrical enzymatic formation of ceramide in giant liposomes. Biophys. J. 2000, 78, 830–838.
- Castro-Gomes, T.; Corrotte, M.; Tam, C.; Andrews, N.W. Plasma Membrane Repair Is Regulated Extracellularly by Proteases Released from Lysosomes. PLoS ONE 2016, 11, e0152583.
- Corrotte, M.; Almeida, P.E.; Tam, C.; Castro-Gomes, T.; Fernandes, M.C.; Millis, B.A.; Cortez, M.; Miller, H.; Song, W.; Maugel, T.K.; et al. Caveolae internalization repairs wounded cells and muscle fibers. eLife 2013, 2, e00926.
- Draeger, A.; Babiychuk, E.B. Ceramide in plasma membrane repair. Handb. Exp. Pharmacol. 2013, 341–353.
- Idone, V.; Tam, C.; Andrews, N.W. Two-way traffic on the road to plasma membrane repair. Trends Cell Biol. 2008, 18, 552–559.
- Tam, C.; Idone, V.; Devlin, C.; Fernandes, M.C.; Flannery, A.; He, X.; Schuchman, E.; Tabas, I.; Andrews, N.W. Exocytosis of acid sphingomyelinase by wounded cells promotes endocytosis and plasma membrane repair. J. Cell Biol. 2010, 189, 1027–1038.
- Corrotte, M.; Fernandes, M.C.; Tam, C.; Andrews, N.W. Toxin pores endocytosed during plasma membrane repair traffic into the lumen of MVBs for degradation. Traffic 2012, 13, 483–494.
- Thiery, J.; Keefe, D.; Saffarian, S.; Martinvalet, D.; Walch, M.; Boucrot, E.; Kirchhausen, T.; Lieberman, J. Perforin activates clathrin- and dynamin-dependent endocytosis, which is required for plasma membrane repair and delivery of granzyme B for granzyme-mediated apoptosis. Blood 2010, 115, 1582–1593.
- Aguado-Velasco, C.; Bretscher, M.S. Circulation of the plasma membrane in Dictyostelium. Mol. Biol. Cell 1999, 10, 4419–4427.
- Tanaka, M.; Kikuchi, T.; Uno, H.; Okita, K.; Kitanishi-Yumura, T.; Yumura, S. Turnover and flow of the cell membrane for cell migration. Sci. Rep. 2017, 7, 12970.
- Tanaka, M.; Fujimoto, K.; Yumura, S. Regulation of the Total Cell Surface Area in Dividing Dictyostelium Cells. Front. Cell Dev. Biol. 2020, 8, 238.
- Wu, L.G.; Hamid, E.; Shin, W.; Chiang, H.C. Exocytosis and endocytosis: Modes, functions, and coupling mechanisms. Annu. Rev. Physiol. 2014, 76, 301–331.
- Kirkham, M.; Nixon, S.J.; Howes, M.T.; Abi-Rached, L.; Wakeham, D.E.; Hanzal-Bayer, M.; Ferguson, C.; Hill, M.M.; Fernandez-Rojo, M.; Brown, D.A.; et al. Evolutionary analysis and molecular dissection of caveola biogenesis. J. Cell Sci. 2008, 121, 2075–2086.
- Babiychuk, E.B.; Monastyrskaya, K.; Potez, S.; Draeger, A. Intracellular Ca2+ operates a switch between repair and lysis of streptolysin O-perforated cells. Cell Death Differ. 2009, 16, 1126–1134.
- Jimenez, A.J.; Maiuri, P.; Lafaurie-Janvore, J.; Divoux, S.; Piel, M.; Perez, F. ESCRT machinery is required for plasma membrane repair. Science 2014, 343, 1247136.
- Li, Z.; Shaw, G.S. Role of calcium-sensor proteins in cell membrane repair. Biosci. Rep. 2023, 43, BSR20220765.
- Blackwood, R.A.; Ernst, J.D. Characterization of Ca2(+)-dependent phospholipid binding, vesicle aggregation and membrane fusion by annexins. Biochem. J. 1990, 266, 195–200.
- Boye, T.L.; Nylandsted, J. Annexins in plasma membrane repair. Biol. Chem. 2016, 397, 961–969.
- Koerdt, S.N.; Ashraf, A.P.K.; Gerke, V. Annexins and plasma membrane repair. Curr. Top. Membr. 2019, 84, 43–65.
- Lauritzen, S.P.; Boye, T.L.; Nylandsted, J. Annexins are instrumental for efficient plasma membrane repair in cancer cells. Semin. Cell Dev. Biol. 2015, 45, 32–38.
- Lennon, N.J.; Kho, A.; Bacskai, B.J.; Perlmutter, S.L.; Hyman, B.T.; Brown, R.H.J. Dysferlin interacts with annexins A1 and A2 and mediates sarcolemmal wound-healing. J. Biol. Chem. 2003, 278, 50466–50473.
- McNeil, A.K.; Rescher, U.; Gerke, V.; McNeil, P.L. Requirement for annexin A1 in plasma membrane repair. J. Biol. Chem. 2006, 281, 35202–35207.
- Jaiswal, J.K.; Lauritzen, S.P.; Scheffer, L.; Sakaguchi, M.; Bunkenborg, J.; Simon, S.M.; Kallunki, T.; Jaattela, M.; Nylandsted, J. S100A11 is required for efficient plasma membrane repair and survival of invasive cancer cells. Nat. Commun. 2014, 5, 3795.
- Ashraf, A.P.K.; Gerke, V. Plasma membrane wound repair is characterized by extensive membrane lipid and protein rearrangements in vascular endothelial cells. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118991.
- Ashraf, A.P.K.; Gerke, V. The resealing factor S100A11 interacts with annexins and extended synaptotagmin-1 in the course of plasma membrane wound repair. Front. Cell Dev. Biol. 2022, 10, 968164.
- Boye, T.L.; Maeda, K.; Pezeshkian, W.; Sonder, S.L.; Haeger, S.C.; Gerke, V.; Simonsen, A.C.; Nylandsted, J. Annexin A4 and A6 induce membrane curvature and constriction during cell membrane repair. Nat. Commun. 2017, 8, 1623.
- Boye, T.L.; Jeppesen, J.C.; Maeda, K.; Pezeshkian, W.; Solovyeva, V.; Nylandsted, J.; Simonsen, A.C. Annexins induce curvature on free-edge membranes displaying distinct morphologies. Sci. Rep. 2018, 8, 10309.
- Mularski, A.; S√∏nder, S.L.; Heitmann, A.S.B.; Pandey, M.P.; Khandelia, H.; Nylandsted, J.; Simonsen, A.C. Interplay of membrane crosslinking and curvature induction by annexins. Sci. Rep. 2022, 12, 22568.
- Ando, Y.; Imamura, S.; Owada, M.K.; Kannagi, R. Calcium-induced intracellular cross-linking of lipocortin I by tissue transglutaminase in A431 cells. Augmentation by membrane phospholipids. J. Biol. Chem. 1991, 266, 1101–1108.
- Kawai, Y.; Wada, F.; Sugimura, Y.; Maki, M.; Hitomi, K. Transglutaminase 2 activity promotes membrane resealing after mechanical damage in the lung cancer cell line A549. Cell Biol. Int. 2008, 32, 928–934.
- Matsuda, C.; Miyake, K.; Kameyama, K.; Keduka, E.; Takeshima, H.; Imamura, T.; Araki, N.; Nishino, I.; Hayashi, Y. The C2A domain in dysferlin is important for association with MG53 (TRIM72). PLoS Curr. 2012, 4, e5035add8caff4.
- Hayes, M.J.; Rescher, U.; Gerke, V.; Moss, S.E. Annexin-actin interactions. Traffic 2004, 5, 571–576.
- Nakamura, M.; Verboon, J.M.; Parkhurst, S.M. Prepatterning by RhoGEFs governs Rho GTPase spatiotemporal dynamics during wound repair. J. Cell Biol. 2017, 216, 3959–3969.
- Monastyrskaya, K.; Babiychuk, E.B.; Hostettler, A.; Wood, P.; Grewal, T.; Draeger, A. Plasma membrane-associated annexin A6 reduces Ca2+ entry by stabilizing the cortical actin cytoskeleton. J. Biol. Chem. 2009, 284, 17227–17242.
- Marko, M.; Prabhu, Y.; Muller, R.; Blau-Wasser, R.; Schleicher, M.; Noegel, A.A. The annexins of Dictyostelium. Eur. J. Cell Biol. 2006, 85, 1011–1022.
- Bonfils, C.; Greenwood, M.; Tsang, A. Expression and characterization of a Dictyostelium discoideum annexin. Mol. Cell Biochem. 1994, 139, 159–166.
- Doring, V.; Veretout, F.; Albrecht, R.; Muhlbauer, B.; Schlatterer, C.; Schleicher, M.; Noegel, A.A. The in vivo role of annexin VII (synexin): Characterization of an annexin VII-deficient Dictyostelium mutant indicates an involvement in Ca(2+)-regulated processes. J. Cell Sci. 1995, 108, 2065–2076.
- Babst, M. A protein’s final ESCRT. Traffic 2005, 6, 2–9.
- Carlton, J. The ESCRT machinery: A cellular apparatus for sorting and scission. Biochem. Soc. Trans. 2010, 38, 1397–1412.
- Christ, L.; Raiborg, C.; Wenzel, E.M.; Campsteijn, C.; Stenmark, H. Cellular Functions and Molecular Mechanisms of the ESCRT Membrane-Scission Machinery. Trends Biochem. Sci. 2017, 42, 42–56.
- Franquelim, H.G.; Schwille, P. Revolving around constriction by ESCRT-III. Nat. Cell Biol. 2017, 19, 754–756.
- Henne, W.M.; Stenmark, H.; Emr, S.D. Molecular mechanisms of the membrane sculpting ESCRT pathway. Cold Spring Harb. Perspect. Biol. 2013, 5, a016766.
- Hurley, J.H. ESCRTs are everywhere. EMBO J. 2015, 34, 2398–2407.
- McCullough, J.; Clippinger, A.K.; Talledge, N.; Skowyra, M.L.; Saunders, M.G.; Naismith, T.V.; Colf, L.A.; Afonine, P.; Arthur, C.; Sundquist, W.I.; et al. Structure and membrane remodeling activity of ESCRT-III helical polymers. Science 2015, 350, 1548–1551.
- Peel, S.; Macheboeuf, P.; Martinelli, N.; Weissenhorn, W. Divergent pathways lead to ESCRT-III-catalyzed membrane fission. Trends Biochem. Sci. 2011, 36, 199–210.
- Scheffer, L.L.; Sreetama, S.C.; Sharma, N.; Medikayala, S.; Brown, K.J.; Defour, A.; Jaiswal, J.K. Mechanism of Ca2+-triggered ESCRT assembly and regulation of cell membrane repair. Nat. Commun. 2014, 5, 5646.
- Jimenez, A.J.; Maiuri, P.; Lafaurie-Janvore, J.; Perez, F.; Piel, M. Laser induced wounding of the plasma membrane and methods to study the repair process. Methods Cell Biol. 2015, 125, 391–408.
- Jimenez, A.J.; Perez, F. Plasma membrane repair: The adaptable cell life-insurance. Curr. Opin. Cell Biol. 2017, 47, 99–107.
- Bohannon, K.P.; Hanson, P.I. ESCRT puts its thumb on the nanoscale: Fixing tiny holes in endolysosomes. Curr. Opin. Cell Biol. 2020, 65, 122–130.
- Denais, C.M.; Gilbert, R.M.; Isermann, P.; McGregor, A.L.; te Lindert, M.; Weigelin, B.; Davidson, P.M.; Friedl, P.; Wolf, K.; Lammerding, J. Nuclear envelope rupture and repair during cancer cell migration. Science 2016, 352, 353–358.
- Radulovic, M.; Schink, K.O.; Wenzel, E.M.; Nahse, V.; Bongiovanni, A.; Lafont, F.; Stenmark, H. ESCRT-mediated lysosome repair precedes lysophagy and promotes cell survival. EMBO J. 2018, 37, e99753.
- Zhen, Y.; Radulovic, M.; Vietri, M.; Stenmark, H. Sealing holes in cellular membranes. EMBO J. 2021, 40, e106922.
- Maki, M.; Suzuki, H.; Shibata, H. Structure and function of ALG-2, a penta-EF-hand calcium-dependent adaptor protein. Sci. China Life Sci. 2011, 54, 770–779.
- Williams, J.K.; Ngo, J.M.; Lehman, I.M.; Schekman, R. Annexin A6 mediates calcium-dependent exosome secretion during plasma membrane repair. Elife 2023, 12, e86556.
- Benaud, C.; Le Dez, G.; Mironov, S.; Galli, F.; Reboutier, D.; Prigent, C. Annexin A2 is required for the early steps of cytokinesis. EMBO Rep. 2015, 16, 481–489.
- Elia, N.; Sougrat, R.; Spurlin, T.A.; Hurley, J.H.; Lippincott-Schwartz, J. Dynamics of endosomal sorting complex required for transport (ESCRT) machinery during cytokinesis and its role in abscission. Proc. Natl. Acad. Sci. USA 2011, 108, 4846–4851.
- Gulluni, F.; Martini, M.; Hirsch, E. Cytokinetic Abscission: Phosphoinositides and ESCRTs Direct the Final Cut. J. Cell Biochem. 2017, 118, 3561–3568.
- Merigliano, C.; Burla, R.; La Torre, M.; Del Giudice, S.; Teo, H.; Liew, C.W.; Chojnowski, A.; Goh, W.I.; Olmos, Y.; Maccaroni, K.; et al. AKTIP interacts with ESCRT I and is needed for the recruitment of ESCRT III subunits to the midbody. PLoS Genet. 2021, 17, e1009757.
- Morita, E.; Sandrin, V.; Chung, H.Y.; Morham, S.G.; Gygi, S.P.; Rodesch, C.K.; Sundquist, W.I. Human ESCRT and ALIX proteins interact with proteins of the midbody and function in cytokinesis. EMBO J. 2007, 26, 4215–4227.
- Tomas, A.; Futter, C.; Moss, S.E. Annexin 11 is required for midbody formation and completion of the terminal phase of cytokinesis. J. Cell Biol. 2004, 165, 813–822.
- Jahan, M.G.S.; Yumura, S. Traction force and its regulation during cytokinesis in Dictyostelium cells. Eur. J. Cell Biol. 2017, 96, 515–528.
- Taira, R.; Yumura, S. A novel mode of cytokinesis without cell-substratum adhesion. Sci. Rep. 2017, 7, 17694.
- DeBello, W.M.; Betz, H.; Augustine, G.J. Synaptotagmin and neurotransmitter release. Cell 1993, 74, 947–950.
- Martinez, I.; Chakrabarti, S.; Hellevik, T.; Morehead, J.; Fowler, K.; Andrews, N.W. Synaptotagmin VII Regulates Ca2+-Dependent Exocytosis of Lysosomes in Fibroblasts. J. Cell Biol. 2000, 148, 1141–1150.
- Chakrabarti, S.; Kobayashi, K.S.; Flavell, R.A.; Marks, C.B.; Miyake, K.; Liston, D.R.; Fowler, K.T.; Gorelick, F.S.; Andrews, N.W. Impaired membrane resealing and autoimmune myositis in synaptotagmin VII-deficient mice. J. Cell Biol. 2003, 162, 543–549.
- Liu, J.; Aoki, M.; Illa, I.; Wu, C.; Fardeau, M.; Angelini, C.; Serrano, C.; Urtizberea, J.A.; Hentati, F.; Hamida, M.B.; et al. Dysferlin, a novel skeletal muscle gene, is mutated in Miyoshi myopathy and limb girdle muscular dystrophy. Nat. Genet. 1998, 20, 31–36.
- Peulen, O.; Rademaker, G.; Anania, S.; Turtoi, A.; Bellahcene, A.; Castronovo, V. Ferlin Overview: From Membrane to Cancer Biology. Cells 2019, 8, 954.
- Han, R.; Bansal, D.; Miyake, K.; Muniz, V.P.; Weiss, R.M.; McNeil, P.L.; Campbell, K.P. Dysferlin-mediated membrane repair protects the heart from stress-induced left ventricular injury. J. Clin. Investig. 2007, 117, 1805–1813.
- Huang, Y.; Laval, S.H.; van Remoortere, A.; Baudier, J.; Benaud, C.; Anderson, L.V.; Straub, V.; Deelder, A.; Frants, R.R.; den Dunnen, J.T.; et al. AHNAK, a novel component of the dysferlin protein complex, redistributes to the cytoplasm with dysferlin during skeletal muscle regeneration. FASEB J. 2007, 21, 732–742.
- Park, S.H.; Han, J.; Jeong, B.-C.; Song, J.H.; Jang, S.H.; Jeong, H.; Kim, B.H.; Ko, Y.-G.; Park, Z.-Y.; Lee, K.E.; et al. Structure and activation of the RING E3 ubiquitin ligase TRIM72 on the membrane. Nat. Struct. Mol. Biol. 2023, 30, 1695–1706.
More
Information
Subjects:
Cell Biology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
397
Revisions:
2 times
(View History)
Update Date:
22 Feb 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No