Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Manmohan Singhal | -- | 5668 | 2024-02-01 08:56:16 | | | |
| 2 | Catherine Yang | Meta information modification | 5668 | 2024-02-01 09:56:26 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Kumari, N.; Anand, S.; Shah, K.; Chauhan, N.S.; Sethiya, N.K.; Singhal, M. Plant-Based Bioactive Compounds as Therapeutics in Parkinson’s Disease. Encyclopedia. Available online: https://encyclopedia.pub/entry/54629 (accessed on 25 June 2026).
Kumari N, Anand S, Shah K, Chauhan NS, Sethiya NK, Singhal M. Plant-Based Bioactive Compounds as Therapeutics in Parkinson’s Disease. Encyclopedia. Available at: https://encyclopedia.pub/entry/54629. Accessed June 25, 2026.
Kumari, Nitu, Santosh Anand, Kamal Shah, Nagendra Singh Chauhan, Neeraj K. Sethiya, Manmohan Singhal. "Plant-Based Bioactive Compounds as Therapeutics in Parkinson’s Disease" Encyclopedia, https://encyclopedia.pub/entry/54629 (accessed June 25, 2026).
Kumari, N., Anand, S., Shah, K., Chauhan, N.S., Sethiya, N.K., & Singhal, M. (2024, February 01). Plant-Based Bioactive Compounds as Therapeutics in Parkinson’s Disease. In Encyclopedia. https://encyclopedia.pub/entry/54629
Kumari, Nitu, et al. "Plant-Based Bioactive Compounds as Therapeutics in Parkinson’s Disease." Encyclopedia. Web. 01 February, 2024.
Copy Citation
Neurological ailments, including stroke, Alzheimer’s disease (AD), epilepsy, Parkinson’s disease (PD), and other related diseases, have affected around 1 billion people globally to date. PD stands second among the common neurodegenerative diseases caused as a result of dopaminergic neuron loss in the midbrain’s substantia nigra regions. Medicinal plants, herbal formulations, and natural bioactive molecules have been gaining much more attention in recent years as synthetic molecules orchestrate a number of undesired effects. Several in vitro, in vivo, and in silico studies in the recent past have demonstrated the therapeutic potential of medicinal plants, herbal formulations, and plant-based bioactives.
phytocompounds
Parkinson’s disease
phenolic acids
antiparkinsonian
neuroprotection
1. Introduction
The brain is the most complicated organ of the body consisting of a complex network of neurons, and functions as a site of intelligence, memory, and cognition, the initiator of body movement, the interpreter of the senses, and the manager of behaviors. It mainly consists of billions of nerves, which are in regular communication through trillions of connections called synapses [1][2]. The brain is subjected to various forms of stresses, including oxidative stress (OS) resulting from the body’s oxygen requirements/utilization and high content of unsaturated fatty acids [3][4]. Nerve cells in the mid-brain degrade slowly, leading to movement- and coordination-related problems, ultimately resulting in neuropathophysiology. Neurodegeneration is associated with the progressive damage of neuronal tissue that causes the irrecoverable loss of neuronal function, subsequent decline in cognitive function, and motor activity [5][6][7]. Among the neurodegenerative diseases, Parkinson’s disease (PD), mild cognitive impairment (MCI), Alzheimer’s disease (AD), epilepsy, multiple sclerosis (MS), amyotrophic lateral sclerosis (ALS), and Huntington’s disease (HD) are of primary importance with distinguished pathological etiologies and clinical management [8][9].
PD is the second common chronic neurodegenerative disorder after AD seen mostly in the aging population [10][11]. It is characterized by synucleipathy, wherein neurons in specific part of the brain undergo damage, resulting in motor signs of muscle stiffness, tremor, and postural instability. Additionally, synucleipathy more specifically refers to a class of neurodegenerative disorders that are characterized by a prion-like spread through interconnected neuronal networks and the abnormal aggregation of α-syn inside glial or neuronal cells. However, the complete etiologies of PD still remain unclear [12][13][14]. The disease’s progression has been pointed out as being related to the depletion of dopamine in the nigrostriatal pathway. Further, intracytoplasmic inclusions of dopamine called Lewy bodies have been identified in patients with PD [12][13]. The deterioration of dopamine-producing neurons inside the substantia nigra (SN), with reduced dopamine release in the striatum, is a major cause of the disease. The SN is located in the midbrain, posterior to the crus cerebri fibers of the cerebral peduncle. It consists of two regions: the substantia nigra pars compacta (SNpc), which harbors dopaminergic neurons, and the substantia nigra pars reticulata (SNpr), comprising gamma-aminobutyric acid-responsive (or GABAergic) neuronal cells. The putamen, striatum, and caudate nuclei are well connected with dopaminergic projections from the SNpc. Each side of an adult SNpc has 400–500 thousand dopaminergic cells, constituting a negligible fraction of total brain mass. As evident in PD, these tiny clusters of cells have a disproportionate impact on motor output and behavior. When compared to other neurons, dopaminergic neurons in the SNpc are more vulnerable to oxidative stress. In the ventral midbrain, these neurons extend to the striatum and play a crucial role in regulating motor behavior in mammals. The SNpc comprises a cluster of cells that releases the neurotransmitter dopamine in the striatum connected to the basal ganglia. The basal ganglion in turn is linked to the thalamus and motor cortex, which are associated with controlling motor output. PD is predominantly attributed to the loss of the majority of cells in the SNpc region [15][16][17]. The factors that are involved in the progression of PD includes ROS, neuroinflammation, mitochondrial dysfunction, the misfolding of proteins, and protein agglomeration, along with several other environmental factors and biological processes [11].
2. Medicinal Plants, Herbal Formulations and Plant-Based Bioactives (Polyphenols, Terpenes, and Alkaloids) as a Potent Therapeutics for PD Management
Medicinal plants possessing high concentrations of additional nutritional elements provide both health advantages and increased nutritional value due to their potency to influence metabolic processes. In the recent decade, medicinal plants and bioactive constituents with diverse structures have shown to be promising resources for PD drug research.
2.1. Polyphenolic Compounds
Polyphenols are a class of significant natural bioactive molecule has been found to be broadly distributed in dietary vegetation and display potential neuroprotective properties against neuroinflammation and neuronal death as evidenced from both in vitro and in vivo experimentation. They have been further classified as flavonoids and non-flavonoids consisting of bioactive compounds including stilbenes, lignans, phenolic acids, curcuminoids, and coumarins.
Polyphenols are secondary metabolites synthesized in plants by the polyketide or shikimate pathway and commonly found in vegetables, fruits, nuts, and seeds. They possess multiple phenol units (C6H5OH) with hydroxyl groups (OH) linked to the aromatic benzene ring. Polyphenols have been found to exhibit potential therapeutic properties. A polyphenols-rich diet has demonstrated significant modulatory effects on the pathophysiological mechanisms of many underlying chronic diseases, especially in diabetes and cardiovascular and neurodegenerative diseases as evidenced from several experimental and clinical studies, indicating their significant prophylactic and therapeutic potential [18]. A growing body of evidence has demonstrated that the supplementation of polyphenolic compounds limits the risk for neurodegenerative disorders. Several of these compounds have been found to have cell-protective abilities against oxysterols (e.g., 7-ketocholesterol),mitigate mitochondrial dysfunction and cell injury. Polyphenols, namely resveratrol, quercetin, and apigenin, have shown potential scavenging activity against reactive oxygen species (ROS) induced by oxysterols and thereby counteracting ROS [19]. They possess influential antioxidant properties owing to their free radical scavenging potential and iron chelating action. Additionally, these have been further documented to display antiviral, antibacterial, anti-inflammatory, anticarcinogenic, and neuroprotective properties [20]. Recently, it has been established that plant polyphenols orchestrate neuroprotective abilities such as the capacity to combat misfolded protein gathering, the probability to endorse cognition, memory, ROS, neuroinflammation, neurotrophin secretion, and the capability to shield nerve cells after neurotoxins exposure [21]. These compounds possess at least one OH group existing over the aromatic side chain with its backbone having a simple moiety to a multifaceted polymer [22].
-
Structure activity relationship
Polyphenolic compounds contain an OH group at an ortho or para position acting as a hydrogen donor and reductant during redox reactions [23]. Antioxidant activity increases with the higher number of total OH groups present. Therefore, polyphenols counteract the oxidation of biomolecules by donating protons rapidly to radicals or by reacting with them to form products that block them from reacting with other biological molecules. Moreover, polyphenols also have the ability to interact with enzymes or receptors in signal transduction, thus modulating cellular oxidation and enhancing the antioxidant status [24]. Preclinical and clinical studies strongly support the protective action of polyphenols in neurodegenerative diseases owing to their high antioxidant activity orchestrated by the presence of OH groups in their structure [25].
-
In vitro and in vivo studies
Recent studies have demonstrated that dietary polyphenols reduce the breakdown of monoamine oxidase A (MAO-A)- and monoamine oxidase B (MAO-B)-dependent monoaminergic neurotransmitters, maintaining the levels of dopamine and serotonin in animal brain tissue [26]. More specifically, MAO-A is responsible for the metabolism of tyramine, norepinephrine (NE), serotonin (5-HT), and dopamine (DA). However, MAO-B mainly metabolizes DA and some less clinically relevant chemicals [27]. In this context, the supplementation of polyphenols to rats after post-fluid percussion injury demonstrated a promising neuroprotective action [28]. Further, research findings have also revealed that a polyphenol-rich diet normalizes brain-derived neurotrophic factor (BDNF) levels and synapsin 1-dependent synaptic plasticity. These studies support the role of polyphenols in the enhancement of memory, learning abilities, and hippocampal neurogenesis. Thus, polyphenols maintains normal brain health by directly influencing the central nervous system and the underlying machinery [29][30].
Polyphenols have been chiefly divided into two categories, namely flavonoids and non-flavonoids in accordance with current recognized classifications [31]. On the basis of the oxidation state and hydroxylation mode, the flavonoids have been further subdivided into flavanones, anthocyanins, flavanols, isoflavones, and flavones, whereas the non-flavonoids are further classified as stilbenes, phenolic acids, phenolic alcohols, lignans, coumarins, and curcuminoids [32][33][34].
2.1.1. Flavonoids
Flavonoids have a common 1,2-diphenylpropane or 1,3-diphenylpropane (C6-C3-C6) basic structure [35]. Various biological properties of flavonoids include antithrombotic, anticancer, anti-inflammatory, antimicrobial, antiviral, and immunomodulation. Flavonoids have been reported in a wide variety of vegetables (tomatoes, onion, cabbage, cauliflower) and fruits (apple, grapes, berries, banana). Li et al. have revealed that these compounds are beneficial to skeletal muscles, liver, pancreas, adipocytes, and neuronal cells [36]. The results of the randomized clinical trials have demonstrated the enhancement of cognitive abilities resulting from the dietary intake of flavonoids-rich foods, irrespective of age and health status [37]. According to structure activity relationship studies, it has been documented that the double bond between the second and third carbon atoms, the 3′,4′-catechol, the ketone group, and the hydroxyl group at the third position present in the flavonoid backbone are responsible for the free radical scavenging and antioxidant properties of flavonoids. Because the double bond between C2–C3 is conjugated to the carbonyl group present in the C ring, unsaturated flavonoids have a higher capacity to scavenge free radicals in relation to saturated compounds like flavanones [38][39]. Mittal et al. mentioned that the extract of Ginkgo biloba rich in flavonoids orchestrates a protective impact on dopaminergic neurons in animal models of PD [37]. In vitro and in vivo findings suggest that flavonoids intake (with supplements or with normal diet) could be a promising intervention for the attenuation and/or prevention of the deterioration effects of cognitive decline associated with various neuronal disorders [40]. Flavonoids consists of potent bioactive compounds including genistein, baicalein, epigallocatechin-3-gallate (EGCG), and hesperidin (Figure 1).

Figure 1. Different types of flavonoids.
Acacetin is a flavone found naturally in plants including Linaria spp., Chrysanthemum morifolium, Calaminth spp., Carthamus tinctorius, Turnera diffusa (known as damiana), and Robiniapseudo acacia (also called black locust) [41][42]. Findings of various studies have reported that neuroinflammation is not only involved in inflammatory diseases but also in neurodegenerative diseases, including PD. Neuroinflammation in PD is associated with microglial and T-lymphocyte activation with an upregulation of pro-inflammatory cytokines like prostaglandin E2 (PGE2), tumor necrosis factor-α (TNF-α), and nitric oxide (NO). Experiments conducted in rodent PD models have suggested that neuroinflammation is prominently implicated in neuronal cell death. Acacetin displays antiparkinsonian activities by diminishing the inflammatory factors associated with the inflammation. It also helps in tumbling dopamine-producing nerve cells, cyclooxygenase-2 (COX-2) glial stimulation, intensifying DA levels and inducible NO synthase (iNOS) [43][44].
Baicalein is one of the main flavonoids and has been reported to be found in roots of the Chinese medicinal herb Scutellaria baicalensis [45]. It has a wide array of biological functions, namely anti-inflammatory, antioxidant, antiviral, anticancer, and cardioprotection [46]. It also inhibits acetylcholinesterase and has neuroprotective ability. Baicalein exhibits antiparkinsonian activity by defending PD through caspase-mediated cell death inhibition and by increasing the feasibility of the SHSY5Y cell line. Specific proteins are also suppressed by this flavonoid, via controlling the neuronal cell damage and the ratio of Bcl-2-associated X protein linked with X protein/B-cell lymphoma (Bax/Bcl-2). It was further reported that baicalein potentially reduces the ROS generation, ATP deficit, apoptosis, and mitochondrial transmembrane breach in PC12 cells, when subjected to rotenone-induced neurotoxicity. Treatment with baicalein increases and maintains basal ganglia dopamine and 5-hydroxytryptamine levels. Additionally, it also decreases the oligomerization and aggregation of α-syn in SH-SY5Y and Hela cells [47][48][49].
Epigallocatechin-3-gallate (EGCG), an essential polyphenolic compound from green tea, has been found to function as an important therapeutic for the treatment of PD [50]. The antiparkinsonian outcome from EGCG is controlled through a rise in reactivators including coactivator-(PGC-1α), peroxisome proliferator-activated receptor, and SIRT1 protein expression. Hence, these are amongst the important metabolic supervisory transcriptase agents which are destined to have an input displaying the inflection in the cellular performance of cells in the anxiety state of PD [51][52].
Theaflavin (TF), is a major constituent of black tea, consisting of three vital compounds, namely TF-3,3′-digallate, TF-3′-gallate, and TF-3-gallate, responsible for lessening the adverse effect in SN TH-positive neurons and employing anti-apoptotic action via suppressing the activities of caspase-9, -8, and -3 in SN [53]. Various scientific studies have been carried out which demonstrated TF to possess excellent neuroprotective role against PD. TF has also been reported to eradicate toxic amyloid deposits. When compared to EGCG, TF3 was less susceptible to air oxidation and had an increased efficacy under oxidizing conditions. Interestingly, TF has been found equal in efficiency to EGCG at inhibiting β-amyloid and α-synuclein-induced neurotoxicity due to its potential antioxidant properties [54][55].
Fustin is a flavanonol extracted from Rhus verniciflua (heartwood). The flavanonol fustin orchestrates its neuroprotective ability by suppressing cell apoptosis, which is further facilitated by the drop in p38 phosphorylation, ROS generation, caspase-3 activation, and the Bax/Bcl-2 ratio [56].
Hesperidin, a flavanone glycoside, has also been found to be associated with multiple neuroprotective activities including the suppression of neuroinflammation, inhibition of oxidative damage, and anti-apoptosis [57]. It diminishes iron-induced death, mitochondrial dysfunction, OS, and reinstates levels of dopamine in the Drosophila melanogaster model of PD. Furthermore, hesperidin also prevents neuroinflammation which was attributed by the augmented synthesis of transforming growth factor-(TGF-) β and IL-10 in an MS mouse model. It possesses the ability to cross the blood–brain barrier (BBB) which can offer its application as a capable therapeutic towards the treatment and management of neurodegenerative diseases [58][59][60].
Anthocyanins are water-soluble flavonoids extensively found in numerous vegetables, as well as fruits such as purple grapes, blackcurrants, blueberries, cherries, and raspberries, respectively. Several studies conducted on cell lines, animal models, and humans have indicated that anthocyanins orchestrate anti-carcinogenic, anti-diabetic, cardiovascular disease prevention, and brain homeostasis [61]. Research findings have documented that anthocyanins exhibits neuroprotective potential through Aβ-inhibition, suppression of inflammatory responses, and reduction in oxidative damage [62][63].
Genistein, an isoflavone derived from Glycine max, has been explored for its implications in various diseases. This compound has attracted attention owing to its pharmacological roles, such as neuroprotection, cardioprotection, anti-cancer, antioxidant activity, anti-inflammatory effects, and obesity prevention. Recently, the synergistic effect of galantamine and genistein was evaluated to explore its neuroprotective ability against Aβ1–42-triggered toxicity in AD. The results of the study demonstrated decreased genotoxicity and cell death by influencing the RAGE/LRP-1 pathway in Wistar rats. Genistein has been extensively studied for its neuroprotective potential in numerous neurodegenerative disorders including PD [64][65][66].
2.1.2. Non-Flavonoids
Non-flavonoids are also polyphenolic compounds exhibiting various types of neuroprotective effects in PD by different mechanisms of action. This group of compounds comprises phenolic acids, phenolic alcohols, stilbenes, and curcumin.
Phenolic Acids
Phenolic acids are secondary metabolites produced by plants. They exist as acidic complexes in the form of hydroxycinnamic and hydroxybenzoic acids (Figure 2A,B) [67]. They possess important biological and pharmacological properties including anti-inflammatory, anticarcinogenic, anticancer, antioxidant, and antimutagenic. Due to the presence of the phenol group and resonance-stabilized conformation, phenolic acids stand out among other chemicals of natural origin for their potent antioxidant and anti-inflammatory activity, which is achieved by a radical scavenging mechanism. Studies carried out using PC12 cells have demonstrated that phenolic acids mitigate 1-methyl-4-phenylpyridinium (MPP+)-induced cell death by boosting the neurite network and triggering the production of proteins essential for synaptogenesis (synaptophysin and synapsin I) and axonal growth (GAP-43). Findings of studies have also demonstrated that phenolic acid reduces the sickness behavior and neuroinflammation induced by lipopolysaccharide (LPS) in mice. TNF-α, a measure of inflammation, has been found to be decreased in the serum in a dose-dependent way, when phenolic acids were administered orally (30 mg/kg) one hour before LPS exposure (1.5 mg/kg) [68]. Phenolic acids are composed of a wide array of bioactive compounds including gallic acid, coumaric acid, ellagic acid (EA), salvianic acid, and rosmarinic acid.
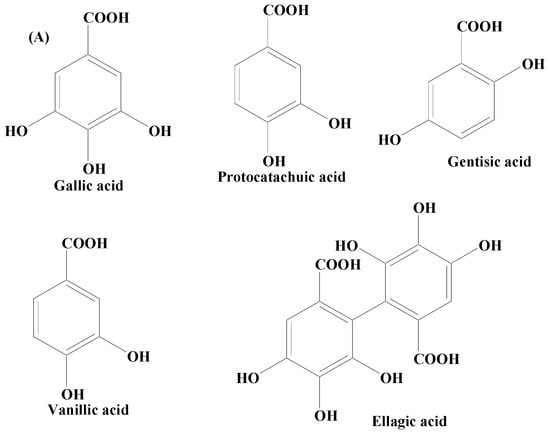
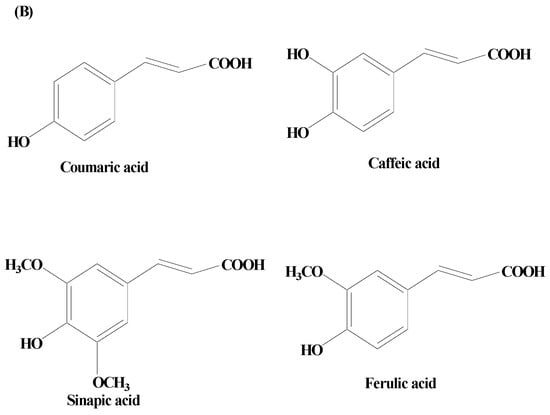
Figure 2. (A) Phenolic acids—hydroxycinnamic acid. (B) Phenolic acids—hydroxybenzoic acid.
Gallic acid (GA) is a phenolic acid widely distributed in grapes, berries, nuts, honey, tea, and vegetables, either in the bound or free form as a derivative. It has been utilized in a variety of healthcare conditions and has been shown to be effective in preventing stroke, cardiovascular problems, Parkinson’s disease, and Alzheimer’s disease. GA’s capacity to cross the BBB, scavenge abnormal levels of ROS and RNS, and bind transition metal ions are the underlying machinery for orchestrating its neuroprotective actions. GA has been demonstrated to end the vicious cycle of OS and tissue injury as a result of its scavenging capacity and the activation of important antioxidant enzymes in the brain. It was further shown that GA reduces neurobehavioral activities by lowering interleukin-1, nitric oxide, myeloperoxidase activity, and TNF-α, and increasing glutathione levels, antioxidant activities, lowering OS, and caspase-3 levels. It also checks apoptosis by bringing down the levels of caspase 3. GA supplementation further enhances the neuromotor abilities that deteriorate during psychosis. It has been found to be promising in reducing lipid peroxidation, controlling dopamine levels, and inflammatory signals [69][70][71].
A polyphenolic compound, para-coumaric acid (p-CA), is a plant-derived secondary metabolite. p-CA is a dietary polyphenol distributed in several natural food sources, such as tomato, carrot, green pepper, and strawberry, functioning majorly as an antioxidant [72][73]. However, a growing body of evidence has reported its beneficial effects including anti-inflammation, antihyperlipidemia, antihyperglycemia, antineurodegeneration, anticancer, anticardiac infarction, and antimicrobial. Research findings have demonstrated the potent neuroprotective potential of p-CA. In vivo findings have shown that p-CA supplementation diminishes the degeneration of axon in sciatic nerves of rat and OS, followed by ischemia or reperfusion [74]. p-CA has also exhibited neuroprotective potential in models of global and local cerebral ischemia by preventing apoptosis and ROS generation [75].
Ellagic acid (EA) is a phenolic compound extensively distributed in dicotyledonous plants possessing robust antioxidant and anti-inflammatory properties. Research also revealed that it improves neural feasibility, lessens neuronic faults, and decreases injury associated with neurodegenerative diseases [76]. EA has also been found to mitigate oxidative damage by controlling the pathways like Nrf2 and NF-κB and also by refining the antioxidants, as well as antioxidant enzymes’ action. Further, findings have revealed that EA could promisingly reduce malondialdehyde heights and amplify proceedings of total GSH, catalase, and superoxide dismutase (SOD) in a PD animal model [77][78].
Salvianolic acid B is obtained from Salvia miltiorrhiza plant. Research findings have suggested that this compound is potentially capable in amending the rate of cell death. Further, it orchestrates its bioactive role through various ways including maintenance of ROS levels, releasing alteration of cells nuclear morphology protecting matrix metalloproteases, tempering cell’s apoptotic and antiapoptotic mediators, and dropping Bax/Bcl-2 ratio, thereby lowering caspase-3 enzyme activity [79][80][81].
Syringic acid is a naturally occurring derivative of benzoic acid distributed widely in edible plants and fruits. It exhibits antiparkinsonian potential by reducing lipid peroxidation, refining the GSH level, and conquering the pro-inflammatory expression of cytokines including COX-2 enzyme, interleukin (IL)-β1, and TNF-α. Motor dysfunction is prevented by means of striatal DA damage and also their metabolites in MPTP-induced experimental model and consequently, the expression level of VMAT-2 and TH inside the SN is amended [82][83][84].
Rosmarinic acid, an important phenolic compound derived from cinnamate is reported in several naturally occurring plants, namely Melissa officinalis, Ocimum basilicum, Salvia officinalis, Origanum majorana, and Rosmarinus officinalis. It has displayed an outstanding antiparkinsonian action by improving the viability of cells, shielding matrix metalloproteases through intracellular ROS production hindrance, elevating DA levels, and controlling the ratio of Bcl-2/Bax. Rosmarinic acid has also been demonstrated to positively influence cell nuclear condensation, the mitochondrial respiratory chain, and several cell morphological fluctuations. In addition, it also helps in the deactivation of caspase-3 which in turn helps in re-establishing the activity of complex I in the mitochondrial electron transport chain [85][86][87].
Phenolic Alcohols
Phenolic alcohols reported in various plants are also polyphenolic compounds containing an OH group linked to an aromatic hydrocarbon. Plants synthesize this class of compounds in order to combat environmental stresses including pathogens or insects that attack plants [88][89]. Phenolic alcohols orchestrate antioxidant and anti-inflammatory activities by rescuing nerve terminals in the striatum and dopaminergic neurons in the SNpc area and restoring SOD, CAT, and glutamate levels, preventing lipid oxidation, reducing the level of ionized calcium binding adaptor molecule (Iba-1), GFAP hyperactivity, pro-inflammatory cytokines, iNOS, and COX-2 activities, respectively. They contain an OH moiety linked to an aromatic hydrocarbon which helps to scavenge free radicals and protect the neuronal damage. In vitro and in vivo studies have shown that phenolic alcohols have significant levels of anti-inflammatory and antioxidant properties due to which they play a very important role in managing PD symptoms [90]. The major phenolic alcohols found predominantly in plants are 6-Shogaol and sesamol.
6-Shogaol (6S) is a pungent ingredient extracted from ginger. It exhibits a wide range of pharmacological properties including neuroprotective potential by overpowering neuroinflammatory factors including TNF-α, NO, COX-2, and iNOS. In addition to these, it also acts through the activation of microglia in the SNpc. Several investigations have documented its neuropharmacological effects for neurodegenerative disorders. In an AD transgenic mice model, 6S prevented aberrant buildup of the Aβ-peptide in the hippocampus and cortical areas and improved memory impairment. Additionally, it has been documented to reduce memory loss, neuronal damage, and neuroinflammatory effects in mice. In studies on 1-methyl-4-phenyl 1,2,3,6-tetrahydropyridine (MPTP)-induced PD models, the anti-PD effects of 6S have been investigated. These studies revealed that 6S remarkably prevents dopaminergic neuronal damage, MPTP-induced motor impairment, and striatal dopamine depletion. In MPTP-induced PD mice, 6S prevents gliosis, dopaminergic neuronal degeneration, and motor impairment. Further, it prevents nuclear factor B’s increased nuclear translocation, as well as apoptotic cell death [91][92][93][94].
Sesamol, a lignin obtained from the shrub Sesamum indicum, holds a wide range of defined neuroprotective potential, and remains utilized as therapeutics against PD. Owing to high metabolic activity, brain requires a lot of energy inputs for physiological functions. The brain also features a high membrane surface to cytoplasm ratio, a low repair capability, a non-replicating nature of neurons, and a comparatively low antioxidant machinery. Due to an imbalance between pro-oxidant and antioxidant agents in the brain, increased free radicals, which are mostly created through oxidative phosphorylation, play a significant role in neurological illnesses. This supports the need to target antioxidant systems in order to combat OS and the resulting brain disorders. By encouraging antioxidative defense systems for neutralizing free radicals and by limiting transcription, the antioxidant system is crucial for saving neuronal cells from OS and maintaining the proper redox balance in the brain tissue. Sesamol has been found to boost the action of various antioxidant enzymes, namely glutathione reductase, CAT, SOD, GPx, and also non-enzymatic antioxidants (vitamin E, GSH, and vitamin C), thereby reducing the levels of the lipid peroxidation and nitrites [95][96][97][98].
Stilbenes
Stilbenes are polyphenolic compounds found in various plant species. They have been shown to have anti-inflammatory properties, estrogen receptor agonist qualities, and effects on cell proliferation, cell signaling pathways, and apoptosis. Stilbenes also exhibit antifungal, antiviral, and antibacterial properties. They possess numerous beneficial attributes for the inhibition of abundant pathophysiological issues, namely age-linked ailments (example: type 2 diabetes mellitus, and obesity), OS, and neurodegenerative disorders including PD [99][100]. In addition, an oligostilbene compound, Amurensin G, found in Vitis amurensis (a type of wild grape) root was revealed to maintain the survivability of SH-SY5Y cells by downregulating α-syn and ubiquitinated proteins [101][102].
A stilbene compound, resveratrol, occurring naturally in several plants including Polygonum cuspidatum has demonstrated protective potential in the animal and cellular models of PD (Figure 3). Heme oxygenase-1 (HOX-1) is a 32 kDa stress response protein implicated in the prevention of PD. HOX-1 functions in response to stress and eventually becomes downregulated. Resveratrol exhibits neuroprotection against paraquat-induced PC12 cells via HOX upregulation. Supplementation of resveratrol has been found to upregulate the SOD enzyme activity [103]. The in vitro findings on antiparkinsonian activity of this compound have revealed the capability of resveratrol to downregulate the level of caspase-3 enzyme activity and the lactate dehydrogenase (LDH) leakage. On a similar line, its hydroxylated derivative oxy-resveratrol also has reported neuroprotective potential through the decrease in ROS generation inside the cell, reduction in phospho-JNK-1 and 2, and cytosolic SIRT1. Due to its hydroxyl group and competing with coenzyme Q, resveratrol decreases the activity of complex III, hence lowering the formation of ROS. Resveratrol has also been demonstrated to protect oxygen glucose deprivation and reperfusion (OGD/R), at least in part, according to a study on the PC12 cell line. It exerts neuroprotective effects by reducing OGD/R-induced OS and maintaining mitochondrial function through PINK1/Parkin-mediated mitophagy [41]. Studies carried out using two-month-old male rats with middle cerebral artery occlusion (MCAO) treated with rehabilitation training and resveratrol indicated that resveratrol enhanced neurological and motor function in MCAO rats via activating the brain-derived neurotrophic factor/tyrosine kinase receptor B (BDNF/TrkB) signaling pathways and SIRT1 signaling network. In both in vivo and in vitro experimental models of neurodegeneration, studies have shown that resveratrol moderates mitochondrial activity, maintains redox homeostasis, and cellular dynamics [104].

Figure 3. Natural stilbene—resveratrol.
Curcumin
Curcumin, also known as diferuloylmethane is a polyphenolic compound obtained from the plant rhizome Curcuma longa. Curcumin exhibits, a broad array of pharmacological properties owing to its hydroxy (antioxidant activity) and methoxy (antitumor and anti-inflammatory activities) groups (Figure 4). It has been established to possess various health benefits. Curcumin’s therapeutic and prophylactic efficacy has been demonstrated in many neurodegenerative, oncological, inflammatory, and autoimmune diseases. Studies carried out on an antiparkinsonian rat model revealed that curcumin treatment potentially controls the PD complications via the dopaminergic neuronal damage and depletion of DA. Moreover, iron chelating activity in addition to condensed iron-positive cells in SN has also been mitigated by curcumin. Research findings have shown that curcumin has the ability to downregulate the level of the caspase-3 enzyme, besides amplifying the LRRK2 mRNA and protein expression in vitro. Accumulated evidence showed that curcumin exhibits various neuroprotective properties, including chelating metal ions’ antioxidation, inhibition of the aggregation of misfolded proteins, and attenuating neuroinflammation [105][106]. According to Jin et al., curcumin was found to alleviate PD by energizing the BDNF/PI3k/Akt transduction pathway [107]. Additionally, a nanoformulation of this medication plus levodopa was recently suggested for the treatment of PD [108]. In a PD model, curcumin also offers protective effects on the cerebellum [109]. Earlier biochemical findings established that curcumin proficiently blocked aggregation of α-syn in vitro. Various improved equivalents of curcumin having enhanced constancy have also been confirmed to be effective in preventing depolymerizing α-syn fibrils and α-syn amyloid aggregation. In vivo studies have demonstrated that curcumin has no effect on how α-syn condensates develop or their initial shape. It does however effectively prevent α-syn from amyloid genesis by reducing its fluidity within the condensates. Additionally, it prevents α-syn E46K and H50Q mutants that are linked to PD illness from aggregating amyloid under phase separation. Curcumin can also weaken α-syn amyloid aggregates that have already developed in the condensates [105][106].

Figure 4. Curcumin.
2.2. Terpenes
Terpenes are one among the most widely distributed compounds in the plant kingdom. They possess the utmost molecular dissimilarity amongst the secondary metabolites. Terpenes are mainly obtained from coniferous plants, namely juniperus, abies, pinus, and picea. They are mainly hydrocarbons constituting the main bioactive components of natural products including essential oil, wax, rubber, and resin. Terpenoids have biological and pharmacological potential including anticancer, antiviral, anti-inflammatory, antifungal, antihyperglycemic, antiparasitic, and antimicrobial [110][111][112][113]. Some of the important terpenes includes carnosic acid, ginkgolide B, and celastrol (Figure 5).
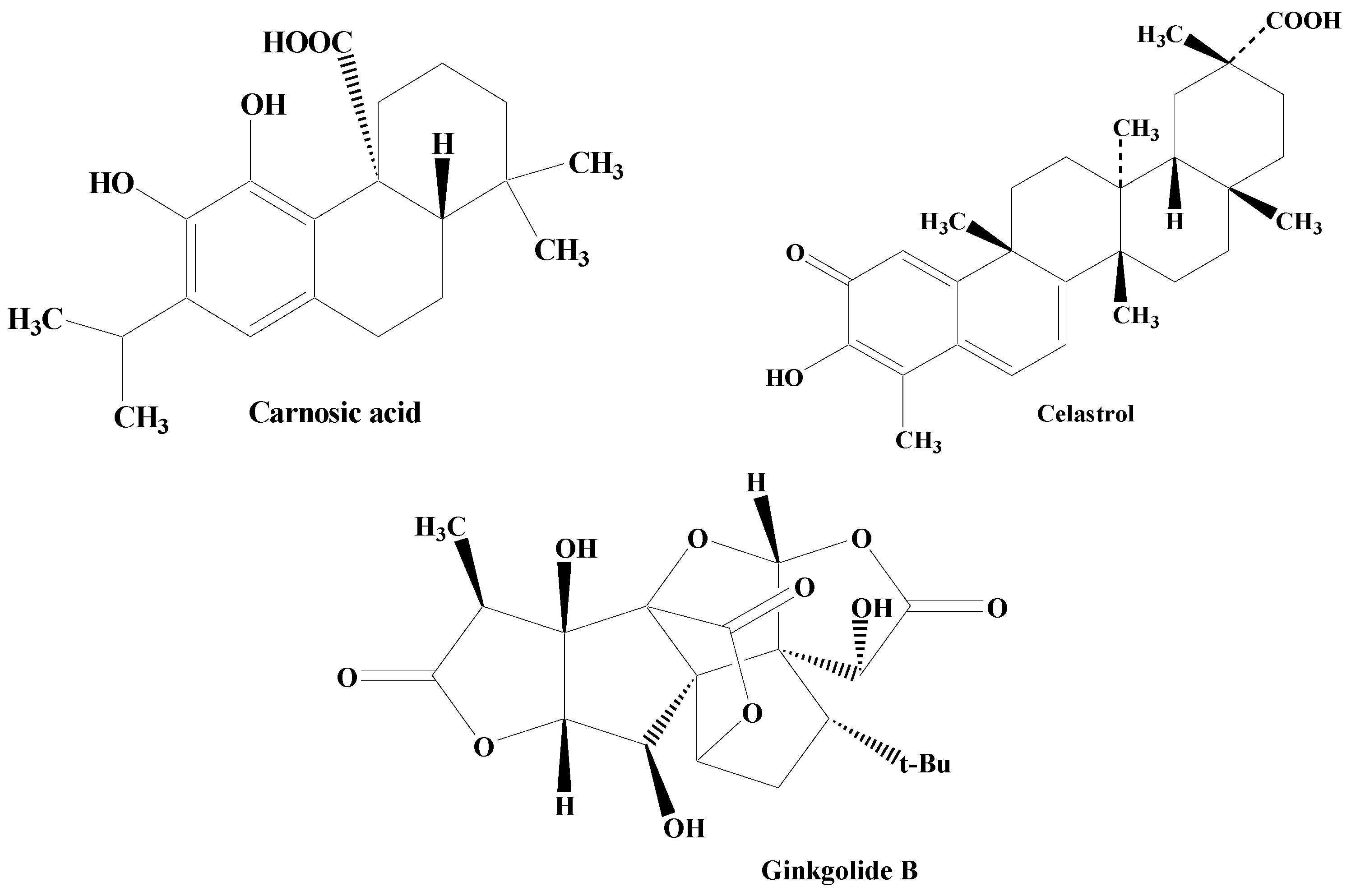
Figure 5. Structures of various terpenes.
Carnosic acid, reported in the herb rosemary, is a phenolic diterpene. It amplifies neural cell capability by enhancing the antioxidant presentation in cellular models of cell apoptosis, and also through interacting with the γ-glutamyl cysteine ligase catalytic subunit, SOD, and GSR stimulation of nuclear factor-E2-linked factor 2 (Nrf2) pathways, brain-derived neurotrophic factor (BDNF) release, and γ-glutamyl cysteine ligase modifier subunit [114][115][116].
One of the important diterpenes, Ginkgolide B, extracted from Ginkgo biloba shields against neuronal damage by lowering the calcium concentration within the cell, declining the action of caspase-3 enzyme and cell death. The calcium-binding protein calbindin D28K encourages neuronal process extension in dopaminergic neurons, thereby possessing the potential to defend dopaminergic neurons against uncontrolled PD. The terpene ginkgolide B holds an outstanding capability of restoring the protein calbindin D28K mRNA, as evidenced by in vitro studies [117][118][119][120][121][122].
A triterpene celastrol found in Tripterygium wilfordii lessens the loss of dopaminergic neurons, thereby minimizing the DA and DOPAC level exhaustion indicating its antiparkinsonian potential. It is associated with the synthesis of various noteworthy intermediates in the inflammation like NF-κβ and TNF-α. Celastrol also orchestrates neuroprotective activity through the attenuation of the loss in the SNpc and dropping reduction in its levels in the striatum. In addition to this, it has also been found to augment the expression of HSP70 in the SNpc. Celastrol is reported to be involved in the nuclear translocation of cytoplasmic HSP70 facilitating HSP70 expression. After inducing expression of HSP70, inflammation is documented to be reduced through the prevention of TNF-α and NF-κB stimulation [123][124][125][126].
-
Structure Activity Relationship
Terpenes are a subclass of hydrocarbons that make up the majority of natural products like rubber, resin, wax, and essential oils. Structurally, terpenes are made up of isoprene units. The fundamental structure of terpenes, isoprene (2-methyl-1,3 butadiene), is made up of short carbon units with two double bonds and five carbon atoms. Terpenes have been classified based on the number of isoprene units. These units arrange themselves in the form of head–tail to form compounds with straight chains or rings. Due to the presence of isoprene in their structure, terpenes have been demonstrated to have antiparkinsonian, antimutagenic, antioxidant, and anticarcinogenic potential. As a result, they are frequently utilized in aromatherapy and phytotherapy. Terpenes have been employed in a variety of industries including food, medicine, cosmetics, pharmacy, and cleaning because of their multifaceted properties. Additionally, in recent years, it has been advised to use several terpenes derived from plants as supplements to enhance overall health [110][127][128].
-
In vitro and in vivo studies
In vitro investigation has showed that the terpene ginkgolide B has a remarkable capacity for repairing the protein calbindin D28K mRNA [108]. Investigations carried out in PC-12 and SH-SY-5Y cell lines demonstrated that ginkgolide B can promote antioxidant mechanisms via the Akt/Nrf2/ARE pathways. In vitro and in vivo studies have demonstrated that terpenes prevent demyelination and work with astrocytes to orchestrate a neuroprotective effect [129][130][131][132].
2.3. Alkaloids
Alkaloids are secondary metabolites, consisting of nitrogen, which at the beginning have been considered as the principal group of bioactive natural compounds isolated from plants (Figure 6). Additionally, alkaloids exhibit much structural diversity including in the chemical skeleton such as tetra-hydro-isoquinoline, indole, pyrrolizidine, tropane, piperidine, quinolizidine, indolizidine, pyridine, pyridinone, quinoline, quinazoline, xanthine, steroid, terpenoid, chromone, and flavoalkaloids, respectively [133]. A wide variety of biological roles such as antidepressant, emetic, diuretic, antimicrobial, antiviral, antihypertensive, anti-inflammatory, antitumor, anticholinergic, myorelaxant, hypoalgesia, and sympathomimetic have been displayed by alkaloids [134]. Various studies have shown that numerous alkaloid components possess a promising relaxing ability for a diverse neuron-related disorders including PD [135]. Thus, natural product-based alkaloids having polypharmacology variation characteristics are very beneficial in the progress of drug development in managing PD [136][137][138]. Zingerone and acetylcorynoline are among the important alkaloids possessing diverse pharmacological functions.
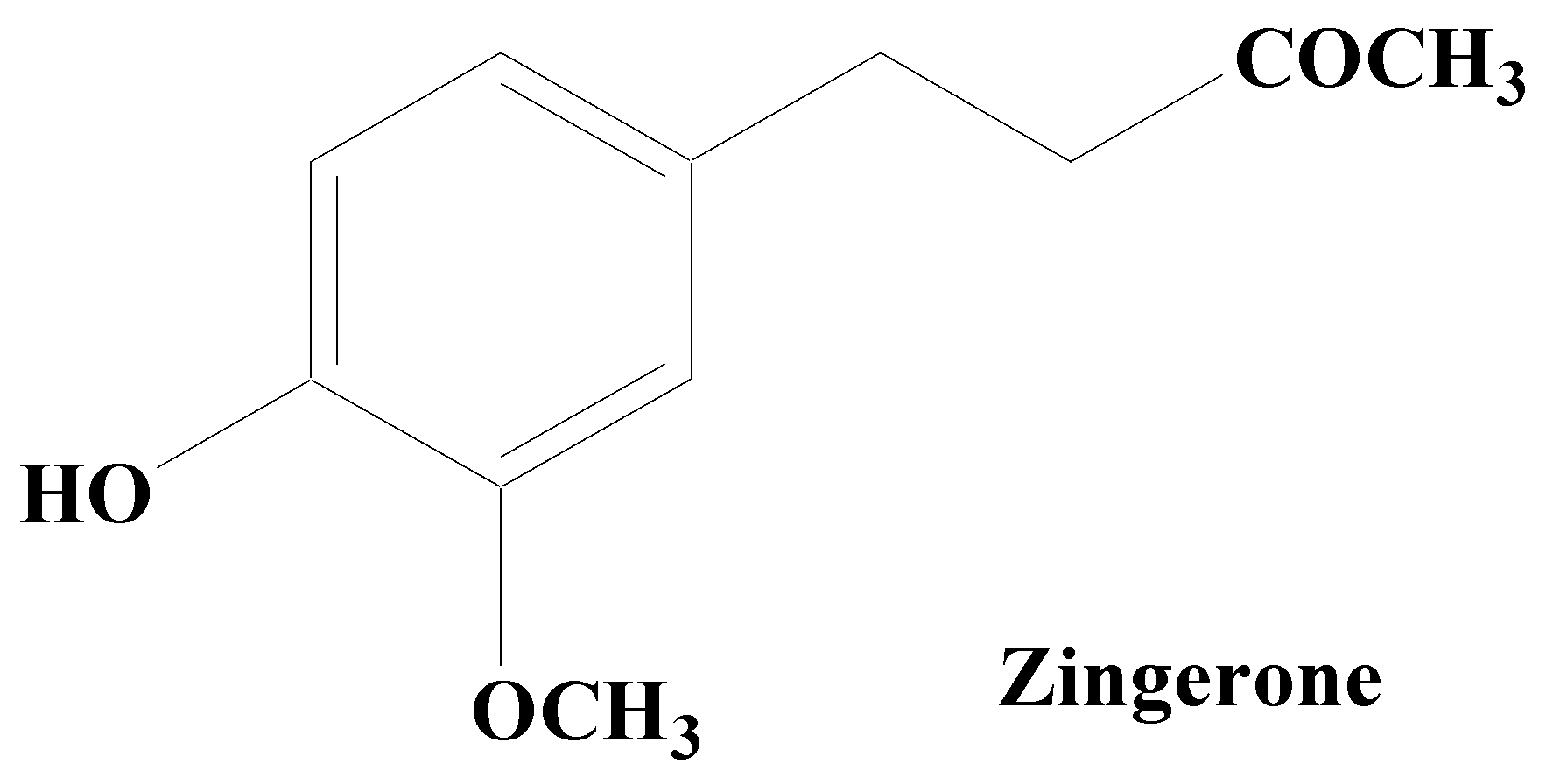
Figure 6. Zingerone.
Zingerone, an alkaloid component found in the rhizome of ginger has been established to possess an outstanding antiparkinsonian potential. By lowering the expression of glial fibrillary acidic protein and IL-1ß in the hippocampus, it inhibits the overactivation of astrocytes and attenuates LPS-induced neuronal cell death. Low antioxidant levels lead to the creation of free radicals (ROS/RNS) and the consequent inflammation is considered to be the major cause for neurodegeneration in PD. Zingerone potentially inhibits the inflammatory cascade components including TNF-α, NO, COX-2, and iNOS that may contribute to lowering memory impairment in animal models of dementia by limiting glial cell activation. It modulates the depletion of dopamine and their metabolic products, namely homovanillic acid and DOPAC. In addition, it also boosts the antioxidative defense like superoxide scavenging and hydroxyl action, with the suppression of OS. Zingerone pretreatment has been found to upregulate the dopamine levels in the nigrostriatal region, which clearly suggests its protective role in the management of PD and its associated symptoms [139][140][141][142][143].
Acetylcorynoline, an alkaloid obtained from Corydalis bungeana, has also been revealed several neuroprotective abilities including inhibition of dopaminergic neuron loss, augmented levels of α-syn, and exhaustion of DA level. Programmed cell death has been documented to be suppressed by dropping the abnormal-1 (egl-1) expression levels, an apoptosis regulator, exhibiting its antiparkinsonian potential. It is well reported that acetylcorynoline is able to prevent pathogenesis in PD via increasing protein breakdown by proteasomes. Acetylcorynoline has been found to facilitate the increased expression of rpn-5, a proteasome-governing subunit, suggesting its antiparkinsonian activity [144].
-
Structure Activity relationship
The alkaloids’ nucleus contains a benzene ring linked to several (three to four) ether bonds having the capacity to generate hydroxyl groups. However, as there are not many hydroxyl groups present on these compounds, they have a low degree of polarity, which makes it easier for them to cross the BBB. Multiple ether bonds are transformed into hydroxyl groups attached to the benzene ring, which makes these aromatic hydroxyl groups powerful antioxidant structures for crossing the BBB. Based on the structural perspectives, alkaloids may act primarily as antioxidants, with anti-inflammatory, autophagy modulation, and suppression of calcium overload effects [145][146][147][148].
-
In vitro and in vivo studies
Several studies have demonstrated a neuroprotective effect of alkaloids against H2O2-induced oxidative damage in SH-SY5Y cell lines and AChE inhibitory activity [149][150]. Studies conducted in vivo subjected to aluminum-induced neuroinflammation and excitotoxicity have revealed that alkaloid therapy was successful in controlling glutamate and acetylcholinesterase levels, which were otherwise raised by aluminum. The findings of this study also been demonstrated to reduce the excitotoxic harm induced by aluminum, as well as the level of expression of the inflammatory markers IL-6 and TNF-α. The increased expression of inflammatory markers in the groups that received alkaloid treatment is suggestive of the neuroprotective ability of alkaloids in the recovery of neuroplasticity. The neuroprotective ability of alkaloid is further supported by histopathology, wherein the therapy greatly reduced neuronal loss and degeneration while restoring healthy and viable neurons. The study’s findings support the hypothesis that alkaloid has neuroprotective properties against neuroinflammation and excitotoxicity brought on by aluminum [151][152][153].
References
- Bassett, D.S.; Gazzaniga, M.S. Understanding complexity in the human brain. Trends Cogn. Sci. 2011, 15, 200–209.
- Colom, R.; Karama, S.; Jung, R.E.; Haier, R.J. Human intelligence and brain networks. Dialogues Clin. Neurosci. 2010, 12, 489–501.
- Salim, S. Oxidative Stress and the Central Nervous System. J. Pharmacol. Exp. Ther. 2017, 360, 201–205.
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583.
- Vasic, V.; Barth, K.; Schmidt, M.H.H. Neurodegeneration and Neuro-Regeneration-Alzheimer’s Disease and Stem Cell Therapy. Int. J. Mol. Sci. 2019, 20, 4272.
- Gao, H.; Hong, J. Why neurodegenerative diseases are progressive: Uncontrolled inflammation drives disease progression. Trends Immunol. 2008, 29, 357–365.
- Zia, A.; Pourbagher-Shahri, A.M.; Farkhondeh, T. Saeed Samarghandian Molecular and cellular pathways contributing to brain aging. Behav. Brain Funct. 2021, 17, 6.
- Rauf, A.; Badoni, H.; Abu-Izneid, T.; Olatunde, A.; Rahman, M.M.; Painuli, S.; Semwal, P.; Wilairatana, P.; Mubarak, M.S. Neuroinflammatory Markers: Key Indicators in the Pathology of Neurodegenerative Diseases. Molecules 2022, 27, 3194.
- Choonara, Y.E.; Pillay, V.; Toit, L.C.; Modi, G.; Naidoo, D.; Ndesendo, V.M.K.; Sibambo1, S.R. Trends in the Molecular Pathogenesis and Clinical Therapeutics of Common Neurodegenerative Disorders. Int. J. Mol. Sci. 2009, 10, 2510–2557.
- Reeve, A.; Simcox, E.; Turnbulla, D. Ageing and Parkinson’s disease: Why is advancing age the biggest risk factor? Ageing Res. Rev. 2014, 14, 19–30.
- Pang, S.Y.Y.; Ho, P.W.L.; Liu, H.F.; Leung, C.; Li, L.; Chang, E.E.S.; Ramsden, D.B.; Ho, S. The interplay of aging, genetics and environmental factors in the pathogenesis of Parkinson’s disease. Transl. Neurodegener. 2019, 8, 23.
- Kouli, A.; Torsney, K.M.; Kuan, W.L. Parkinson’s Disease: Etiology, Neuropathology, and Pathogenesis. In Parkinson’s Disease: Pathogenesis and Clinical Aspects ; Stoker, T.B., Greenland, J.C., Eds.; Codon Publications: Brisbane, Australia, 21 December 2018; Chapter 1. Available online: https://www.ncbi.nlm.nih.gov/books/NBK536722/ (accessed on 13 November 2023).
- Ma, J.; Gao, J.; Wang, J.; Xie, A. Prion-Like Mechanisms in Parkinson’s Disease. Front. Neurosci. 2019, 13, 552.
- Calabresi, P.; Mechelli, A.; Natale, G.; Volpicelli-Daley, L.; Lazzaro, G.D.; Ghiglieri, V. Alpha-synuclein in Parkinson’s disease and other synucleinopathies: From overt neurodegeneration back to early synaptic dysfunction. Cell Death Dis. 2023, 14, 176.
- Ni, A.; Ernst, C. Evidence That Substantia Nigra Pars Compacta Dopaminergic Neurons Are Selectively Vulnerable to Oxidative Stress Because They Are Highly Metabolically Active. Front. Cell. Neurosci. 2022, 16, 826193.
- González-Hernández, T.; Rodríguez, M. Compartmental organization and chemical profile of dopaminergic and GABAergic neurons in the substantia nigra of the rat. J. Comp. Neurol. 2000, 421, 107–135.
- Gröger, A.; Kolb, R.; Schäfer, R.; Klose, U. Dopamine Reduction in the Substantia Nigra of Parkinson’s Disease Patients Confirmed by In Vivo Magnetic Resonance Spectroscopic Imaging. PLoS ONE 2014, 9, e84081.
- Singh, A.K.; Singla, R.K.; Pandey, A.K. Chlorogenic Acid: A Dietary Phenolic Acid with Promising Pharmacotherapeutic Potential. Curr. Med. Chem. 2023, 30, 3905–3926.
- Uddin, M.S.; Al Mamun, A.; Kabir, M.T.; Ahmad, J.; Jeandet, P.; Sarwar, M.S.; Ashraf, G.M.; Aleya, L. Neuroprotective role of polyphenols against oxidative stress-mediated neurodegeneration. Eur. J. Pharmacol. 2020, 886, 173412.
- Arias-Sánchez, R.A.; Torner, L.; Fenton Navarro, B. Polyphenols and Neurodegenerative Diseases: Potential Effects and Mechanisms of Neuroprotection. Molecules 2023, 28, 5415.
- Efimova, S.S.; Ostroumova, O.S. Modulation of the Dipole Potential of Model Lipid Membranes with Phytochemicals: Molecular Mechanisms, Structure-Activity Relationships, and Implications in Reconstituted Ion Channels. Membranes 2023, 13, 453.
- Anand, S.; Sowbhagya, R.; Ansari, M.A.; Alzohairy, M.A.; Alomary, M.N.; Almalik, A.I.; Ahmad, W.; Tripathi, T.; Elderdery, A.Y. Polyphenols and Their Nanoformulations: Protective Effects against Human Diseases. Life 2022, 12, 1639.
- Liu, S.; Cheng, L.; Liu, Y.; Zhan, S.; Wu, Z.; Zhang, X. Relationship between Dietary Polyphenols and Gut Microbiota: New Clues to Improve Cognitive Disorders, Mood Disorders and Circadian Rhythms. Foods 2023, 12, 1309.
- Platzer, M.; Kiese, S.; Tybussek, T.; Herfellner, T.; Schneider, F.; Schweiggert-Weisz, U.; Eisner, P. Radical Scavenging Mechanisms of Phenolic Compounds: A Quantitative Structure-Property Relationship (QSPR) Study. Front. Nutr. 2022, 9, 882458.
- Lopez-Corona, A.V.; Valencia-Espinosa, I.; González-Sánchez, F.A.; Sánchez-López, A.L.; Garcia-Amezquita, L.E.; Garcia-Varela, R. Antioxidant, Anti-Inflammatory and Cytotoxic Activity of Phenolic Compound Family Extracted from Raspberries (Rubus idaeus): A General Review. Antioxidants 2022, 11, 1192.
- Zhang, Z.; Hamada, H.; Gerk, P.M. Selectivity of Dietary Phenolics for Inhibition of Human Monoamine Oxidases A and B. BioMed Res. Int. 2019, 2019, 8361858.
- Shih, J.C.; Chen, K.; Ridd, M.J. MONOAMINE OXIDASE: From Genes to Behavior. Annu. Rev. Neurosci. 1999, 22, 197–217.
- Carecho, R.; Carregosa, D.; Ratilal, B.O.; Figueira, I.; Ávila-Gálvez, M.A.; Santos, C.N.; Loncarevic-Vasiljkovic, N. Dietary (Poly)phenols in Traumatic Brain Injury. Int. J. Mol. Sci. 2023, 24, 8908.
- Grabska-Kobyłecka, I.; Szpakowski, P.; Król, A.; Książek-Winiarek, D.; Kobyłecki, A.; Głąbiński, A.; Nowak, D. Polyphenols and Their Impact on the Prevention of Neurodegenerative Diseases and Development. Nutrients 2023, 15, 3454.
- Vauzour, D. Dietary Polyphenols as Modulators of Brain Functions: Biological Actions and Molecular Mechanisms Underpinning Their Beneficial Effects. Oxid. Med. Cell. Longev. 2012, 2012, 914273.
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects—A review. J. Funct. Foods 2015, 18, 820–897.
- Li, M.; Qian, M.; Jiang, Q.; Tan, B.; Yin, Y.; Han, X. Evidence of Flavonoids on Disease Prevention. Antioxidants 2023, 12, 527.
- Sun, W.; Shahrajabian, M.H. Therapeutic Potential of Phenolic Compounds in Medicinal Plants—Natural Health Products for Human Health. Molecules 2023, 28, 1845.
- Yan, L.; Guo, M.; Zhang, Y.; Yu, L.; Wu, J.; Tang, Y.; Ai, W.; Zhu, F.; Law, B.Y.; Chen, Q.; et al. Dietary Plant Polyphenols as the Potential Drugs in Neurodegenerative Diseases: Current Evidence, Advances, and Opportunities. Oxid. Med. Cell. Longev. 2022, 2022, 5288698.
- Chen, S.; Wang, X.; Cheng, Y.; Gao, H.; Chen, X. A Review of Classification, Biosynthesis, Biological Activities and Potential Applications of Flavonoids. Molecules 2023, 28, 4982.
- Bellavite, P. Neuroprotective Potentials of Flavonoids: Experimental Studies and Mechanisms of Action. Antioxidants 2023, 12, 280.
- Mittal, P.; Dhankhar, S.; Chauhan, S.; Garg, N.; Bhattacharya, T.; Ali, M.; Chaudhary, A.A.; Rudayni, H.A.; Al-Zharani, M.; Ahmad, W. A Review on Natural Antioxidants for Their Role in the Treatment of Parkinson’s Disease. Pharmaceuticals 2023, 16, 908.
- Spiegel, M.; Andruniów, T.; Sroka, Z. Flavones’ and Flavonols’ Antiradical Structure–Activity Relationship—A Quantum Chemical Study. Antioxidants 2020, 9, 461.
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750.
- Shahpiri, Z.; Bahramsoltani, R.; Farzaei, M.H.; Farzaei, F.; Rahimi, R. Phytochemicals as future drugs for Parkinson’s disease: A comprehensive review. Rev. Neurosci. 2016, 27, 651–668.
- Devi, S.; Kumar, V.; Singh, S.K.; Dubey, A.K.; Kim, J.J. Flavonoids: Potential Candidates for the Treatment of Neurodegenerative Disorders. Biomedicines 2021, 9, 99.
- Singh, S.; Gupta, P.; Meena, A.; Luqman, S. Acacetin, a flavone with diverse therapeutic potential in cancer, inflammation, infections and other metabolic disorders. Food Chem. Toxicol. 2020, 145, 111708.
- Kim, H.G.; Ju, M.S.; Ha, S.K.; Lee, H.; Lee, H.; Kim, S.Y.; Oh, M.S. Acacetin protects dopaminergic cells against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neuroinflammation in vitro and in vivo. Biol. Pharm. Bull. 2012, 35, 1287–1294.
- Kim, S.M.; Park, Y.J.; Shin, M.; Kim, H.; Kim, M.J.; Lee, S.H.; Yun, S.P.; Kwon, S. Acacetin inhibits neuronal cell death induced by 6-hydroxydopamine in cellular Parkinson’s disease model. Bioorg. Med. Chem. Lett. 2017, 27, 5207–5212.
- Zhao, Q.; Chen, X.; Martin, C. Scutellaria baicalensis, the golden herb from the garden of Chinese medicinal plants. Sci. Bull. 2016, 61, 1391–1398.
- Jadhav, R.; Kulkarni, Y.A. Effects of baicalein with memantine on aluminium chloride-induced neurotoxicity in Wistar rats. Front. Pharmacol. 2023, 14, 1034620.
- Li, Y.; Zhao, J.; Hölscher, C. Therapeutic Potential of Baicalein in Alzheimer’s Disease and Parkinson’s Disease. CNS Drugs 2017, 31, 639–652.
- Wang, Y.; Wei, N.; Li, X. Preclinical Evidence and Possible Mechanisms of Baicalein for Rats and Mice with Parkinson’s Disease: A Systematic Review and Meta-Analysis. Front. Aging Neurosci. 2020, 12, 277.
- Chen, M.; Peng, L.; Gong, P.; Zheng, X.; Sun, T.; Zhang, X.; Huo, J. Baicalein Induces Mitochondrial Autophagy to Prevent Parkinson’s Disease in Rats via miR-30b and the SIRT1/AMPK/mTOR Pathway. Front. Neurol. 2021, 12, 646817.
- Wang, Y.; Wu, S.; Li, Q.; Lang, W.; Li, W.; Jiang, X.; Wan, Z.; Chen, J.; Wang, H. Epigallocatechin-3-gallate: A phytochemical as a promising drug candidate for the treatment of Parkinson’s disease. Front. Pharmacol. 2022, 13, 977521.
- Sergi, C.M. Epigallocatechin gallate for Parkinson’s disease. Clin. Exp. Pharmacol. Physiol. 2022, 49, 1029–1041.
- Xu, Y.; Xie, M.; Xue, J.; Xiang, L.; Li, Y.; Xiao, J.; Xiao, G.; Wang, H. EGCG ameliorates neuronal and behavioral defects by remodeling gut microbiota and TotM expression in Drosophila models of Parkinson’s disease. FASEB J. 2020, 34, 5931–5950.
- Caruana, M.; Vassallo, N. Tea Polyphenols in Parkinson’s Disease. Adv. Exp. Med. Biol. 2015, 863, 117–137.
- Anandhan, A.; Tamilselvam, K.; Radhiga, T.; Rao, S.; Essa, M.M.E.; Manivasagam, T. Theaflavin, a black tea polyphenol, protects nigral dopaminergic neurons against chronic MPTP/probenecid induced Parkinson’s disease. Brain Res. 2012, 1433, 104–113.
- Grelle, G.; Otto, A.; Lorenz, M.; Frank, R.F.; Wanker, E.E.; Bieschke, J. Black tea theaflavins inhibit formation of toxic amyloid-β and α-synuclein fibrils. Biochemistry 2011, 50, 10624–10636.
- Park, B.C.; Lee, Y.S.; Park, H.; Kwak, M.; Yoo, B.K.; Kim, J.Y.; Kim, J. Protective effects of fustin, a flavonoid from Rhus verniciflua Stokes, on 6-hydroxydopamine-induced neuronal cell death. Exp. Mol. Med. 2007, 39, 316–326.
- Hajialyani, M.; Farzaei, M.H.; Echeverría, J.; Nabavi, S.M.; Uriarte, E.; Sobarzo-Sánchez, E. Hesperidin as a Neuroprotective Agent: A Review of Animal and Clinical Evidence. Molecules 2019, 24, 648.
- Atoki, A.V.; Aja, P.M.; Shinkafi, T.S.; Ondari, E.N.; Awuchi, C.G. Hesperidin plays beneficial roles in disorders associated with the central nervous system: A review. Int. J. Food Prop. 2023, 26, 1867–1884.
- Tamilselvam, K.; Braidy, N.; Manivasagam, T.; Essa, M.M.; Prasad, N.R.; Karthikeyan, S.; Thenmozhi, A.J.; Selvaraju, S.; Guillemin, G.J. Neuroprotective Effects of Hesperidin, a Plant Flavanone, on Rotenone-Induced Oxidative Stress and Apoptosis in a Cellular Model for Parkinson’s Disease. Oxid. Med. Cell. Longev. 2013, 2013, 102741.
- Antunes, M.S.; Goes, A.T.R.; Boeira, S.P.; Prigol, M.; Jesse, C.R. Protective effect of hesperidin in a model of Parkinson’s disease induced by 6-hydroxydopamine in aged mice. Nutrition 2014, 30, 1415–1422.
- Li, P.; Feng, D.; Yang, D.; Li, X.; Sun, J.; Wang, G.; Tian, L.; Jiang, X.; Bai, W. Protective effects of anthocyanins on neurodegenerative diseases. Trends Food Sci. Technol. 2021, 117, 205–217.
- Winter, A.N.; Bickford, P.C. Anthocyanins and Their Metabolites as Therapeutic Agents for Neurodegenerative Disease. Antioxidants 2019, 8, 333.
- Strathearn, K.E.; Yousef, G.G.; Grace, M.H.; Roy, S.L.; Tambe, M.A.; Ferruzzi, M.G.; Wu, Q.; Simon, J.E.; Lila, M.A.; Rochet, J. Neuroprotective effects of anthocyanin- and proanthocyanidin-rich extracts in cellular models of Parkinsons disease. Brain Res. 2014, 1555, 60–77.
- Li, R.; Robinson, M.; Ding, X.; Geetha, T.; Al-Nakkash, L.; Broderick, T.L.; Babu, J.R. Genistein: A focus on several neurodegenerative diseases. J. Food Biochem. 2022, 46, e14155.
- Siddique, Y.H.; Naz, F.; Jyoti, S.; Ali, F.; Rahul. Effect of Genistein on the Transgenic Drosophila Model of Parkinson’s Disease. J. Diet. Suppl. 2019, 16, 550–563.
- Liu, L.; Chen, W.; Xie, J.; Wong, M. Neuroprotective effects of genistein on dopaminergic neurons in the mice model of Parkinson’s disease. Neurosci. Res. 2008, 60, 156–161.
- Khoddami, A.; Wilkes, M.A.; Roberts, T.H. Techniques for Analysis of Plant Phenolic Compounds. Molecules 2013, 18, 2328–2375.
- Caruso, G.; Godos, J.; Privitera, A.; Lanza, G.; Castellano, S.; Chillemi, A.; Bruni, O.; Ferri, R.; Caraci, F.; Grosso, G. Phenolic Acids and Prevention of Cognitive Decline: Polyphenols with a Neuroprotective Role in Cognitive Disorders and Alzheimer’s Disease. Nutrients 2022, 14, 819.
- Shabani, S.; Rabiei, Z.; Amini-Khoei, H. Exploring the multifaceted neuroprotective actions of gallic acid: A review. Int. J. Food Prop. 2020, 23, 736–752.
- Chandrasekhar, Y.; Kumar, G.P.; Ramya, E.M.; Anilakumar, K.R. Gallic Acid Protects 6-OHDA Induced Neurotoxicity by Attenuating Oxidative Stress in Human Dopaminergic Cell Line. Neurochem. Res. 2018, 43, 1150–1160.
- Abdelsalam, S.A.; Renu, K.; Zahra, H.A.; Abdallah, B.M.; Ali, E.M.; Veeraraghavan, V.P.; Sivalingam, K.; Ronsard, L.; Ammar, R.B.; Vidya, D.S.; et al. Polyphenols Mediate Neuroprotection in Cerebral Ischemic Stroke-An Update. Nutrients 2023, 15, 1107.
- Pathan, A.S.; Jain, P.G.; Kumawat, V.S.; Katolkar, U.N.; Surana, S.J. Neuroprotective Effects of P-Coumaric Acid on Haloperidol-Induced Catalepsy Through Ameliorating Oxidative Stress and Brain Dopamine Level. J. Pharmacol. Pharmacother. 2022, 13, 364–374.
- Dolrahman, N.; Mukkhaphrom, W.; Sutirek, J.; Thong-Asa, W. Benefits of p-coumaric acid in mice with rotenone-induced neurodegeneration. Metab. Brain Dis. 2023, 38, 373–382.
- Guven, M.; Yuksel, Y.; Sehitoglu, M.H.; Tokmak, M.; Aras, A.B.; Akman, T.; Golge, U.H.; Goksel, F.; Karavelioglu, E.; Cosar, M. The Effect of Coumaric Acid on Ischemia-Reperfusion Injury of Sciatic Nerve in Rats. Inflammation 2015, 38, 2124–2132.
- Pluta, R.; Miziak, B.; Czuczwar, S.J. Apitherapy in Post-Ischemic Brain Neurodegeneration of Alzheimer’s Disease Proteinopathy: Focus on Honey and Its Flavonoids and Phenolic Acids. Molecules 2023, 28, 5624.
- Sharifi-Rad, J.; Quispe, C.; Castillo, C.M.S.; Caroca, R.; Lazo-Vélez, M.A.; Antonyak, H.; Polishchuk, A.; Lysiuk, R.; Oliinyk, P.; Masi, L.D.; et al. Ellagic Acid: A Review on Its Natural Sources, Chemical Stability, and Therapeutic Potential. Oxid. Med. Cell. Longev. 2022, 2022, 3848084.
- Ardah, M.T.; Bharathan, G.; Kitada, T.; Haque, M.E. Ellagic Acid Prevents Dopamine Neuron Degeneration from Oxidative Stress and Neuroinflammation in MPTP Model of Parkinson’s Disease. Biomolecules 2020, 10, 1519.
- Sarkaki, A.; Farbood, Y.; Dolatshahi, M.; Mansouri, S.M.T.; Khodadadi, A. Neuroprotective Effects of Ellagic Acid in a Rat Model of Parkinson’s Disease. Acta Med. Iran. 2016, 54, 494–502.
- Tian, L.; Wang, X.; Sun, Y.; Li, C.; Xing, Y.; Zhao, H.; Duan, M.; Zhou, Z.; Wang, S. Salvianolic acid B, an antioxidant from Salvia miltiorrhiza, prevents 6-hydroxydopamine induced apoptosis in SH-SY5Y cells. Int. J. Biochem. Cell Biol. 2008, 40, 409–422.
- Zhou, J.; Qu, X.; Li, Z.; Ji, W.; Liu, Q.; Ma, Y.; He, J. Salvianolic Acid B Attenuates Toxin-Induced Neuronal Damage via Nrf2-Dependent Glial Cells-Mediated Protective Activity in Parkinson’s Disease Models. PLoS ONE 2014, 9, e101668.
- Zhao, R.; Liu, X.; Zhang, L.; Yang, H.; Zhang, Q. Current Progress of Research on Neurodegenerative Diseases of Salvianolic Acid B. Oxid. Med. Cell. Longev. 2019, 2019, 3281260.
- Güzelad, O.; Özkan, A.; Parlak, H.; Sinen, O.; Afşar, E.; Öğüt, E.; Yıldırım, F.B.; Bülbül, M.; Ağar, A.; Aslan, M. Protective mechanism of Syringic acid in an experimental model of Parkinson’s disease. Metab. Brain Dis. 2021, 36, 1003–1014.
- Ogut, E.; Armagan, K.; Gül, Z. The role of syringic acid as a neuroprotective agent for neurodegenerative disorders and future expectations. Metab. Brain Dis. 2022, 37, 859–880.
- Cheemanapalli, S.; Mopuri, R.; Golla, R.; Anuradha, C.M.; Chitta, S.K. Syringic acid (SA)—A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557.
- Presti-Silva, S.M.; Herlinger, A.L.; Martins-Silva, C.; Pires, R.G.W. Biochemical and behavioral effects of rosmarinic acid treatment in an animal model of Parkinson’s disease induced by MPTP. Behav. Brain Res. 2023, 440, 114257.
- Lv, R.; Du, L.; Liu, X.; Zhou, F.; Zhang, Z.; Zhang, L. Rosmarinic acid attenuates inflammatory responses through inhibiting HMGB1/TLR4/NF-κB signaling pathway in a mouse model of Parkinson’s disease. Life Sci. 2019, 223, 158–165.
- Cai, G.; Lin, F.; Wu, D.; Lin, C.; Chen, H.; Wei, Y.; Weng, H.; Chen, Z.; Wu, M.; Huang, E.; et al. Rosmarinic Acid Inhibits Mitochondrial Damage by Alleviating Unfolded Protein Response. Front. Pharmacol. 2022, 13, 859978.
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320.
- Dehghanian, Z.; Habibi, K.; Dehghanian, M.; Aliyar, S.; Asgari Lajayer, B.; Astatkie, T.; Minkina, T.; Keswani, C. Reinforcing the bulwark: Unravelling the efficient applications of plant phenolics and tannins against environmental stresses. Heliyon 2022, 8, e09094.
- Giuliano, C.; Cerri, S.; Blandini, F. Potential therapeutic effects of polyphenols in Parkinson’s disease: In vivo and in vitro pre-clinical studies. Neural Regen. Res. 2021, 16, 234–241.
- Huh, E.; Choi, J.G.; Choi, Y.; Ju, I.G.; Noh, D.; Shin, D.; Kim, D.H.; Park, H.; Oh, M.S. 6-Shogaol, an Active Ingredient of Ginger, Improves Intestinal and Brain Abnormalities in Proteus Mirabilis-Induced Parkinson’s Disease Mouse Model. Biomol. Ther. 2023, 31, 417–424.
- Ha, S.K.; Moon, E.; Ju, M.S.; Kim, D.H.; Ryu, J.H.; Oh, M.S.; Kim, S.Y. 6-Shogaol, a ginger product, modulates neuroinflammation: A new approach to neuroprotection. Neuropharmacology 2012, 63, 211–223.
- Gwon, M.G.; Gu, H.; Leem, J.; Park, K.K. Protective Effects of 6-Shogaol, an Active Compound of Ginger, in a Murine Model of Cisplatin-Induced Acute Kidney Injury. Molecules 2021, 26, 5931.
- Sandoval-Avila, S.; Diaz, N.F.; Gómez-Pinedo, U.; Canales-Aguirre, A.A.; Gutiérrez-Mercado, Y.K.; Padilla-Camberos, E.; Marquez-Aguirre, A.L.; Díaz-Martínez, N.E. Neuroprotective effects of phytochemicals on dopaminergic neuron cultures. Neurologia 2019, 34, 114–124.
- Angeline, M.S.; Sarkar, A.; Anand, K.; Ambasta, R.K.; Kumar, P. Sesamol and naringenin reverse the effect of rotenone-induced PD rat model. Neuroscience 2013, 254, 379–394.
- Singh, N.; Vishwas, S.; Kaur, A.; Kaur, H.; Kakoty, V.; Khursheed, R.; Chaitanya, M.V.N.L.; Babu, M.R.; Awasthi, A.; Corrie, L.; et al. Harnessing role of sesamol and its nanoformulations against neurodegenerative diseases. Biomed. Pharmacother. 2023, 167, 115512.
- Bosebabu, B.; Cheruku, S.P.; Chamallamudi, M.R.; Nampoothiri, M.; Shenoy, R.R.; Nandakumar, K.; Parihar, V.K.; Kumar, N. An Appraisal of Current Pharmacological Perspectives of Sesamol: A Review. Mini Rev. Med. Chem. 2020, 20, 988–1000.
- Shah, A.; Lobo, R.; Krishnadas, N.; Surubhotla, R. Sesamol and Health—A Comprehensive Review. Indian J. Pharm. Educ. 2019, 53, S28–S42.
- Teka, T.; Zhang, L.; Ge, X.; Li, Y.; Han, L.; Yan, X. Stilbenes: Source plants, chemistry, biosynthesis, pharmacology, application and problems related to their clinical Application—A comprehensive review. Phytochemistry 2022, 197, 113128.
- El Khawand, T.; Courtois, A.; Valls, J.; Richard, T.; Krisa, S. A review of dietary stilbenes: Sources and bioavailability. Phytochem. Rev. 2018, 17, 1007–1029.
- Huang, K.S.; Lin, M. Oligostilbenes from the roots of Vitis amurensis. J. Asian Nat. Prod. Res. 1999, 2, 21–28.
- Ryu, H.; Oh, W.K.; Jang, I.; Park, J. Amurensin G induces autophagy and attenuates cellular toxicities in a rotenone model of Parkinson’s disease. Biochem. Biophys. Res. Commun. 2013, 433, 121–126.
- Zhang, L.; Dong, M.; Deng, J.; Zhang, C.; Liu, M. Resveratrol exhibits neuroprotection against paraquat-induced PC12 cells via heme oxygenase 1 upregulation by decreasing MiR-136-5p expression. Bioengineered 2022, 13, 7065–7081.
- Santos, M.G.D.; Schimith, L.E.; André-Miral, C.; Muccillo-Baisch, A.L.; Arbo, B.D.; Hort, M.A. Neuroprotective Effects of Resveratrol in In vivo and In vitro Experimental Models of Parkinson’s Disease: A Systematic Review. Neurotox. Res. 2022, 40, 319–345.
- Nebrisi, E.E. Neuroprotective Activities of Curcumin in Parkinson’s Disease: A Review of the Literature. Int. J. Mol. Sci. 2021, 22, 11248.
- Patel, A.; Olang, C.A.; Lewis, G.; Mandalaneni, K.; Anand, N.; Gorantla, V.R. An Overview of Parkinson’s Disease: Curcumin as a Possible Alternative Treatment. Cureus 2022, 14, e25032.
- Jin, T.; Zhang, Y.; Botchway, B.O.A.; Zhang, J.; Fan, R.; Zhang, Y.; Liu, X. Curcumin can improve Parkinson’s disease via activating BDNF/PI3k/Akt signaling pathways. Food Chem. Toxicol. 2022, 164, 113091.
- Mogharbel, B.F.; Cardoso, M.A.; Irioda, A.C.; Stricker, P.E.F.; Slompo, R.C.; Appel, J.M.; Oliveira, N.B.; Perussolo, M.C.; Saçaki, C.S.; Rosa, N.N.; et al. Biodegradable Nanoparticles Loaded with Levodopa and Curcumin for Treatment of Parkinson’s Disease. Molecules 2022, 27, 2811.
- Fikry, H.; Saleh, L.A.; Gawad, S.A. Neuroprotective effects of curcumin on the cerebellum in a rotenone- induced Parkinson’s Disease Model. CNS Neurosci. Ther. 2022, 28, 732–748.
- Xu, Y.; Wei, H.; Gao, J. Natural Terpenoids as Neuroinflammatory Inhibitors in LPS-stimulated BV-2 Microglia. Mini Rev. Med. Chem. 2021, 21, 520–534.
- Del Prado-Audelo, M.L.; Cortés, H.; Caballero-Florán, I.H.; González-Torres, M.; Escutia-Guadarrama, L.; Bernal-Chávez, S.A.; Giraldo-Gomez, D.M.; Magaña, J.J.; Leyva-Gómez, G. Therapeutic Applications of Terpenes on Inflammatory Diseases. Front. Pharmacol. 2021, 12, 704197.
- González-Burgos, E.; Gómez-Serranillos, M.P. Terpene compounds in nature: A review of their potential antioxidant activity. Curr. Med. Chem. 2012, 19, 5319–5341.
- Masyita, A.; Sari, R.M.; Astuti, A.D.; Yasir, B.; Rumata, N.R.; Emran, T.B.; Nainu, F.; Simal-Gandaraf, J. Terpenes and terpenoids as main bioactive compounds of essential oils, their roles in human health and potential application as natural food preservatives. Food Chem. X 2022, 13, 100217.
- Wu, C.; Tsai, C.; Chang, S.; Lin, C.; Huang, L.; Tsai, C. Carnosic acid protects against 6-hydroxydopamine-induced neurotoxicity in in vivo and in vitro model of Parkinson’s disease: Involvement of antioxidative enzymes induction. Chem. Biol. Interact. 2015, 225, 40–46.
- Lai, C.Y.; Lin, C.Y.; Wu, C.R.; Tsai, C.H.; Tsai, C.W. Carnosic Acid Alleviates Levodopa-Induced Dyskinesia and Cell Death in 6-Hydroxydopamine-lesioned Rats and in SH-SY5Y Cells. Front. Pharmacol. 2021, 12, 703894.
- Mirza, F.J.; Zahid, S.; Holsinger, R.M.D. Neuroprotective Effects of Carnosic Acid: Insight into Its Mechanisms of Action. Molecules 2023, 28, 2306.
- Jiang, Y.; Liu, Y.; Ding, C.; Feng, X. Protective Effect of Ginkgolide B on Dopaminergic Neurons and Expression of Neuroinflammatory Factors in 6-OHDA-Induced Parkinson’s Disease Mice. J. Biomater. Tissue Eng. 2020, 10, 1312–1317.
- Yin, J.; Miao, Q.; Wang, Q.; Huang, J.; Xiao, B.; Ma, C. The protective effect of Ginkgolide B on MPTP-induced Parkinson mice. J. Neurol. Sci. 2021, 429, 119481.
- Zhao, Y.; Xiong, S.; Liu, P.; Liu, W.; Wang, Q.; Liu, Y.; Tan, H.; Chen, X.; Shi, X.; Wang, Q.; et al. Polymeric Nanoparticles-Based Brain Delivery with Improved Therapeutic Efficacy of Ginkgolide B in Parkinson’s Disease. Int. J. Nanomed. 2020, 15, 10453–10467.
- Nabavi, S.M.; Habtemariam, S.; Daglia, M.; Braidy, N.; Loizzo, M.R.; Tundis, R.; Nabavi, S.F. Neuroprotective Effects of Ginkgolide B Against Ischemic Stroke: A Review of Current Literature. Curr. Top. Med. Chem. 2015, 15, 2222–2232.
- Gachowska, M.; Szlasa, W.; Saczko, J.; Kulbacka, J. Neuroregulatory role of ginkgolides. Mol. Biol. Rep. 2021, 48, 5689–5697.
- Xia, S.; Fang, D. Pharmacological action and mechanisms of ginkgolide B. Chin. Med. J. 2007, 120, 922–928.
- Shi, J.; Li, J.; Xu, Z.; Chen, L.; Luo, R.; Zhang, C.; Gao, F.; Zhang, J.; Fu, C. Celastrol: A Review of Useful Strategies Overcoming its Limitation in Anticancer Application. Front. Pharmacol. 2020, 11, 558741.
- Lin, M.; Lin, C.C.; Chen, Y.; Yang, H.; Hung, S. Celastrol Inhibits Dopaminergic Neuronal Death of Parkinson’s Disease through Activating Mitophagy. Antioxidants 2019, 9, 37.
- Cui, Y.; Jiang, X.; Feng, J. The therapeutic potential of triptolide and celastrol in neurological diseases. Front. Pharmacol. 2022, 13, 1024955.
- Bai, X.; Fu, R.; Zhang, S.; Yue, S.; Chen, Y.; Xu, D.; Tang, Y. Potential medicinal value of celastrol and its synthesized analogues for central nervous system diseases. Biomed. Pharmacother. 2021, 139, 111551.
- Mosquera, M.E.G.; Jiménez, G.; Tabernero, V.; Vinueza-Vaca, J.; García-Estrada, C.; Kosalková, K.; Sola-Landa, A.; Monje, B.; Acosta, C.; Alonso, R.; et al. Terpenes and Terpenoids: Building Blocks to Produce Biopolymers. Sustain. Chem. 2021, 2, 467–492.
- Souza, M.T.S.; Almeida, J.R.G.S.; Araujo, A.A.S.; Duarte, M.C.; Gelain, D.P.; Moreira, J.C.F.; Santos, M.R.V.; Quintans-Júnior, L.J. Structure–activity relationship of terpenes with anti-inflammatory profile- a systematic review. Basic Clin. Pharmacol. Toxicol. 2014, 115, 244–256.
- Zhang, W.; Song, J.; Yan, R.; Li, L.; Xiao, Z.; Zhou, W.; Wang, Z.; Xiao, W.; Du, G. Diterpene ginkgolides protect against cerebral ischemia/reperfusion damage in rats by activating Nrf2 and CREB through PI3K/Akt signaling. Acta Pharmacol. Sin. 2018, 39, 1259–1272.
- Moratilla-Rivera, I.; Sánchez, M.; Valdés-González, J.A.; Gómez-Serranillos, M.P. Natural Products as Modulators of Nrf2 Signaling Pathway in Neuroprotection. Int. J. Mol. Sci. 2023, 24, 3748.
- Wang, L.; Lei, Q.; Zhao, S.; Xu, W.J.; Dong, W.; Ran, J.H.; Shi, Q.H.; Fu, J.F. Ginkgolide B Maintains Calcium Homeostasis in Hypoxic Hippocampal Neurons by Inhibiting Calcium Influx and Intracellular Calcium Release. Front. Cell. Neurosci. 2021, 14, 627846.
- Akkol, E.K.; Çankaya, I.T.; Karatoprak, G.S.; Carpar, E.; Sobarzo-Sánchez, E.; Capasso, R. Natural Compounds as Medical Strategies in the Prevention and Treatment of Psychiatric Disorders Seen in Neurological Diseases. Front. Pharmacol. 2021, 12, 669638.
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in Chemical Structures and Biological Properties of Plant Alkaloids. Molecules 2021, 26, 3374.
- Omar, F.; Tareq, A.M.; Alqahtani, A.M.; Dhama, K.; Sayeed, M.A.; Emran, T.B.; Simal-Gandara, J. Plant-Based Indole Alkaloids: A Comprehensive Overview from a Pharmacological Perspective. Molecules 2021, 26, 2297.
- Hussain, G.; Rasul, A.; Anwar, H.; Aziz, N.; Razzaq, A.; Wei, W.; Ali, M.; Li, J.; Li, X. Role of Plant Derived Alkaloids and Their Mechanism in Neurodegenerative Disorders. Int. J. Biol. Sci. 2018, 14, 341–357.
- Kempste, P.; Ma, A. Parkinson’s disease, dopaminergic drugs and the plant world. Front. Pharmacol. 2022, 13, 970714.
- Aryal, B.; Raut, B.K.; Bhattarai, S.; Bhandari, S.; Tandan, P.; Gyawali, K.; Sharma, K.; Ranabhat, D.; Thapa, R.; Aryal, D.; et al. Potential Therapeutic Applications of Plant-Derived Alkaloids against Inflammatory and Neurodegenerative Diseases. Evid.-Based Complement. Altern. Med. 2022, 2022, 7299778.
- Kong, Y.R.; Tay, K.C.; Su, Y.X.; Wong, C.K.; Tan, W.N.; Khaw, K.Y. Potential of Naturally Derived Alkaloids as Multi-Targeted Therapeutic Agents for Neurodegenerative Diseases. Molecules 2021, 26, 728.
- Kabuto, H.; Yamanushi, T.T. Effects of zingerone and eugenol on the pathological progress in the 6-hydroxydopamine-induced Parkinson’s disease mouse model. Neurochem. Res. 2011, 36, 2244–2249.
- Kabuto, H.; Nishizawa, M.; Tada, M.; Higashio, C.; Shishibori, T.; Kohno, M. Zingerone prevents 6-hydroxydopamine-induced dopamine depression in mouse striatum and increases superoxide scavenging activity in serum. Neurochem. Res. 2005, 30, 325–332.
- Saraiva, A.A.; Silva, J.P.O.D.; Sousa, J.V.M.; Brandim, A.D. Molecular Docking of Zingerone and Gamma-Mangostin to Inhibit MAO-B and Catechol-O-Methyltransferase (COMT) in the Treatment of Parkinson’s Disease. Res. Soc. Dev. 2022, 11, e189111637853.
- Rashid, S.; Wali, A.F.; Rashid, S.M.; Alsaffar, R.M.; Ahmad, A.; Jan, B.L.; Paray, B.A.; Alqahtani, S.M.A.; Arafah, A.; Rehman, M.U. Zingerone Targets Status Epilepticus by Blocking Hippocampal Neurodegeneration via Regulation of Redox Imbalance, Inflammation and Apoptosis. Pharmaceuticals 2021, 14, 146.
- Ahmad, B.; Rehman, M.U.; Amin, I.; Arif, A.; Rasool, S.; Bhat, S.A.; Afzal, I.; Hussain, I.; Bilal, S.; Mir, M.R. A Review on Pharmacological Properties of Zingerone (4-(4-Hydroxy-3-methoxyphenyl)-2-butanone). Sci. World J. 2015, 2015, 816364.
- Fu, R.; Wang, Y.; Chen, C.; Tsai, R.; Liu, S.; Chang, W.; Lin, H.; Lu, C.; Wei, J.; Wang, Z.; et al. Acetylcorynoline attenuates dopaminergic neuron degeneration and α-synuclein aggregation in animal models of Parkinson’s disease. Neuropharmacology 2014, 82, 108–120.
- Han, N.; Yang, Z.; Liu, Z.; Liu, H.; Yin, J. Research Progress on Natural Benzophenanthridine Alkaloids and their Pharmacological Functions: A Review. Nat. Prod. Commun. 2016, 11, 1181–1188.
- Charlton, N.C.; Mastyugin, M.; Török, B.; Török, M. Structural Features of Small Molecule Antioxidants and Strategic Modifications to Improve Potential Bioactivity. Molecules 2023, 28, 1057.
- Zhang, Y.; Yang, Y.; Yang, X. Blood-brain barrier permeability and neuroprotective effects of three main alkaloids from the fruits of Euodia rutaecarpa with MDCK-pHaMDR cell monolayer and PC12 cell line. Biomed. Pharmacother. 2018, 98, 82–87.
- Li, J.; Wu, Y.; Dong, S.; Yu, Y.; Wu, Y.; Xiang, B.; Li, Q. Research Progress on Neuroprotective Effects of Isoquinoline Alkaloids. Molecules 2023, 28, 4797.
- Xiao, X.; Tong, Z.; Zhang, Y.; Zhou, H.; Luo, M.; Hu, T.; Hu, P.; Kong, L.; Liu, Z.; Yu, C.; et al. Novel Prenylated Indole Alkaloids with Neuroprotection on SH-SY5Y Cells against Oxidative Stress Targeting Keap1–Nrf2. Mar. Drugs 2022, 20, 191.
- Kongkiatpaiboon, S.; Duangdee, N.; Prateeptongkum, S.; Chaijaroenkul, W. Acetylcholinesterase Inhibitory Activity of Alkaloids Isolated from Stephania venosa. Nat. Prod. Commun. 2016, 11, 1805–1806.
- Nishal, S.; Phaugat, P.; Bazaad, J.; Dhaka, R.; Khatkar, S.; Khatkar, A.; Khayatkashani, M.; Alizadeh, P.; Haghighi, S.M.; Mehri, M.; et al. A Concise Review of Common Plant-derived Compounds as a Potential Therapy for Alzheimer’s Disease and Parkinson’s Disease: Insight into Structure-Activity-Relationship. CNS Neurol. Disord. Drug Targets 2023, 22, 1057–1069.
- Bhusal, C.K.; Uti, D.E.; Mukherjee, D.; Alqahtani, T.; Alqahtani, S.; Bhattacharya, A.; Akash, S. Unveiling Nature’s potential: Promising natural compounds in Parkinson’s disease management. Park. Relat. Disord. 2023, 10, 105799.
- Baburaj, R.; Kuntal, D.A. Neuroprotective role of a protoberberine alkaloid against aluminium-induced neuroinflammation and excitotoxicity. Not. Sci. Biol. 2023, 15, 11488.
More
Information
Subjects:
Medicine, Research & Experimental
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
711
Revisions:
2 times
(View History)
Update Date:
01 Feb 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No