Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Adrián Hernández-Díazcouder | -- | 3855 | 2024-01-30 15:12:14 | | | |
| 2 | Jessie Wu | Meta information modification | 3855 | 2024-01-31 02:02:28 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hernández-Díazcouder, A.; Romero-Nava, R.; Sánchez-Muñoz, F.; Guzmán-Martín, C.A.; Huang, F. The Roles of MicroRNAs in Asthma. Encyclopedia. Available online: https://encyclopedia.pub/entry/54535 (accessed on 27 June 2026).
Hernández-Díazcouder A, Romero-Nava R, Sánchez-Muñoz F, Guzmán-Martín CA, Huang F. The Roles of MicroRNAs in Asthma. Encyclopedia. Available at: https://encyclopedia.pub/entry/54535. Accessed June 27, 2026.
Hernández-Díazcouder, Adrián, Rodrigo Romero-Nava, Fausto Sánchez-Muñoz, Carlos A. Guzmán-Martín, Fengyang Huang. "The Roles of MicroRNAs in Asthma" Encyclopedia, https://encyclopedia.pub/entry/54535 (accessed June 27, 2026).
Hernández-Díazcouder, A., Romero-Nava, R., Sánchez-Muñoz, F., Guzmán-Martín, C.A., & Huang, F. (2024, January 30). The Roles of MicroRNAs in Asthma. In Encyclopedia. https://encyclopedia.pub/entry/54535
Hernández-Díazcouder, Adrián, et al. "The Roles of MicroRNAs in Asthma." Encyclopedia. Web. 30 January, 2024.
Copy Citation
Asthma is one of the most common chronic non-communicable diseases worldwide, characterized by variable airflow limitation secondary to airway narrowing, airway wall thickening, and increased mucus resulting from chronic inflammation and airway remodeling. Current epidemiological studies reported that hypovitaminosis D is frequent in patients with asthma and is associated with worsening the disease and that supplementation with vitamin D3 improves asthma symptoms. However, despite several advances in the field, the molecular mechanisms of asthma have yet to be comprehensively understood. MicroRNAs play an important role in controlling several biological processes and their deregulation is implicated in diverse diseases, including asthma.
asthma
inflammation
microRNAs
Vitamin D3
1. The Role of MicroRNA-21 in Asthma and Its Modulation by Vitamin D3
miR-21 is an important miRNA regulating the inflammatory response in various diseases, including asthma. In clinical studies, for instance, a recent study conducted on asthmatic patients reported increased levels of miR-21-5p in exosomes in patients with moderate asthma compared to those with healthy subjects, while patients with severe asthma presented reduced levels of miR-21-5p in exosomes [1]. Likewise, a recent study on patients with mild and moderate–severe asthma reported that moderate–severe asthma patients presented elevated levels of miR-21-5p in exosomes from the plasma compared with both mild asthma and healthy subjects [2]. Also, the same study reported that the levels of miR-21-5p in exosomes correlated positively with IgE levels and correlated negatively with plasma levels of TNF-α and IL-6 in patients with moderate–severe asthma [2]. Another recent study on asthmatic patients reported that asthmatic patients presented high levels of miR-21-5p and IL-4 in serum compared to healthy subjects [3]. The same study found a positive correlation between miR-21-5p levels and IL-4 levels; likewise, IL-4 predicted miR-21-5p expression levels [3]. A study with children reported that asthmatic children presented an increase in 42.6-fold in miR-21-5p plasma levels and an increase in IL-13 levels compared to healthy children [4]. Also, the same study reported that the plasma levels of miR-21-5p correlated positively with IL-13 levels and eosinophil percentage and correlated negatively with lung function (FEV1) [4]. In contrast, a study conducted on patients with a crossover phenotype of bronchial asthma (AB) and chronic obstructive pulmonary disease (COPD) reported that in those patients, there were decreased circulating levels of miR-21-5p compared with healthy subjects [5]. Furthermore, the same study found that low levels of miR-21-5p were associated with less reversibility of bronchial obstruction [5]. Therefore, evidence suggests that miR-21-5p participates in allergic lung inflammation and the pathogenesis of asthma.
In experimental studies within animal models, miR-21-5p was implicated in the development and exacerbation of asthma. For instance, in a recent study with a rat asthma model induced by ovalbumin (OVA), the expression of miR-21-5p was observed mainly in alveolar macrophages [6]. To validate the above result, rat alveolar macrophages were stimulated with LPS (to mimic an inflammatory state present in asthma), resulting in the increased expression of miR-21-5p in exosomes [6]. Besides, the transfer of the miR-21-5p-enriched exosomes from alveolar macrophages to rat airway epithelial cells induced an epithelial–mesenchymal transition through the regulation of Smad7 (Figure 1a) [6]. Moreover, it was reported that the expression of miR-21-5p in the human airway smooth muscle (ASM) cells induced an increase in the appearance and migration of these cells because of the regulation of homologous phosphatase and tensin (PTEN), resulting in remodeling of the airways (Figure 1a) [7]. The above evidence suggests that miR-21-5p is involved in the inflammatory state and the remodeling of airway epithelial cells through the induction of the epithelial–mesenchymal transition, which is an important factor for the development and exacerbation of asthma. In contrast, an OVA-induced asthma model in miR-21 knockout (KO) mice showed that the lack of miR-21 led to a decrease in airway hyperresponsiveness and eosinophilic inflammation and a reduction in Th2 cell cytokines (IL-4, IL-5, and IL-13) in the bronchoalveolar lavage fluid (BALF), along with an increase in the levels of IL-12 and IFN-γ due to the suppression of the polarization of M2 macrophages (Figure 1c) [8]. A recent study conducted in an asthmatic mouse model induced by OVA found the up-regulation of miR-21-5p in airway epithelial cells compared to control mice [9]. Likewise, the same study reported that the transfer of miR-21-5p-enriched exosomes of mast cells to asthmatic mice induced a state of oxidative stress and inflammation in epithelial cells of the airway as well as exacerbation of asthmatic symptoms by the negative regulation of dimethylarginine dimethylaminohydrolase 1 (DDAH1), resulting in the inhibition of Wnt/β-catenin signaling activity (Figure 1a) [9]. Therefore, evidence indicates that miR-21-5p is involved in remodeling and inflammation in asthma pathology.
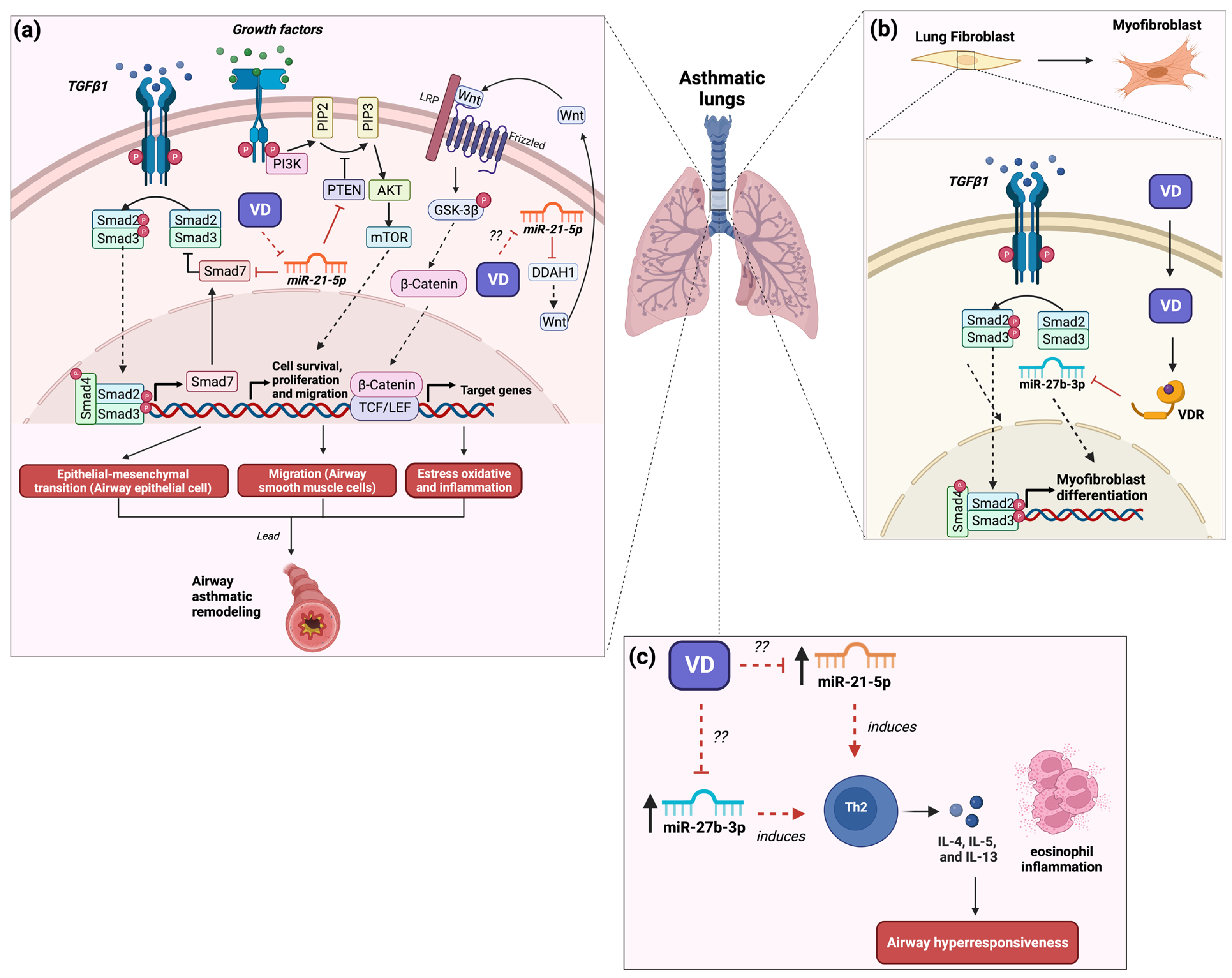
Figure 1. Role of miR-21 and miR-27b in the development of asthma and the possible implication of vitamin D3 supplementation. (a) The expression of miR-21-5p regulates signaling pathways such as TGFβ, mTOR, and Wnt/β-catenin involved in airway remodeling and inflammation. Vitamin D3 supplementation may decrease the expression of miR-21-5p, improving asthma symptoms by reducing TGFβ and mTOR activity and increasing β-catenin activity. The question marks indicate a possible effect of vitamin D3 on oxidative stress and inflammation through the regulation of miR-21. (b) The expression of miR-27b-3p in lung tissue promotes fibroblast differentiation to myofibroblast. The interaction between vitamin D3 and VDR reduces the expression of miR-27b-3p, inhibiting the differentiation of lung fibroblasts. (c) The expression of miR-21-5p and miR-27b-3p induce Th2 differentiation, IL-4, IL-5, and IL-13 release, and eosinophil inflammation, resulting in airway hyperresponsiveness. The question marks suggest a possible effect of vitamin D3 on airway hyperresponsiveness via the regulation of miR-21 and miR-27b, which could result in the reduction of Th2 cell levels. Studies showed that vitamin D3 supplementation improves airway hyperresponsiveness, which may be by regulating these miRNAs [10]. The red lines illustrate the regulatory effects of both vitamin D3 and miRNAs. Created with BioRender.com.
On the other hand, few clinical studies reported a relationship between miR-21-5p and vitamin D status. In aorta tissue from atherosclerotic patients with a vitamin D deficiency (<25 nm/L), the specimens presented a greater expression of miR-21-5p with a mean of 6.7-fold, while in patients with vitamin D sufficiency (>25 nm/L), the tissue presented a lower expression of miR-21-5p with a mean of 1.7-fold [10]. Another study in diabetic type 1 patients found that diabetic patients with vitamin D deficiency (>25 nm/L) presented an increase in circulating levels of miR-21-5p, while non-diabetic patients with both sufficiency and insufficiency of vitamin D presented a decrease in circulating levels of miR-21-5p [11]. The above evidence suggests that miR-21-5p could be regulated by vitamin D. A recent study found that in TGFβ1-stimulated human bronchial fibroblast cells (asthma cell model) treated with calcitriol, the downregulation of miR-21-5p expression was induced (Figure 1a) [12]. This miRNA is related to TGFβ/Smad signaling pathway activation and fibrosis involved in the pathogenesis of pulmonary fibrosis [13]. Hence, the treatment with vitamin D3 has beneficial effects on asthma by inhibiting the remodeling of epithelial and fibroblasts in the lung through the regulation of miR-21 expression. Thus, the beneficial effects of vitamin D3 on lung tissue remodeling may be through the regulation of the expression of miR-21-5p. Thus, the beneficial effects of vitamin D3 on lung tissue remodeling may be through the regulation of the expression of miR-21-5p. Nevertheless, to date, few studies exist that explore the role of the treatment of vitamin D3 on the regulation of miR-21-5p in asthma patients as a plausible therapeutic strategy.
2. The Role of MicroRNA-27b in Asthma and Its Modulation by Vitamin D3
miR-27b-3p is a miRNA that plays an important role in regulating various physical and pathological processes, including asthma, by regulating the inflammatory response and apoptosis [14][15]. In clinical studies, for example, a study in children with mild to moderate asthma identified a negative association between levels of miR-27b-3p in serum and lung function FEV1% [16]. Moreover, a study conducted on pediatric patients with dust mite-induced asthma and dust mite-induced asthma with food allergy presented a downregulation of miR-27b-3p expression compared with children with only food allergy and healthy children [17]. It also was found that Spleen tyrosine kinase (SYK) and epidermal growth factor receptor (EGFR) regulate the development of asthma by the induction of the PI3K-AKT pathway and influence cell differentiation of T or B cells [18]. These genes are targets of miR-27b-3p, which suggests that miR-27b-3p may regulate the inflammation present in asthma by regulating the PI3K-AKT pathway [17]. However, more studies on this interaction need to be validated. In contrast, a study with patients with severe asthma showed that the treatment with anti-IL-5 promoted the downregulation of miR-27b-3p, improved lung function, and decreased peripheral eosinophil counts [19]. Furthermore, in asthmatic patients an up-regulation of miR-27b-3p in blood samples compared to healthy controls was found and it was showed, through bioinformatic analysis, that serpin family E member 1 (SERPINE1) and RAR-related orphan receptor A (RORA) are genes target of miR-27b-3p, which are genes associated with allergic asthma [20]. Likewise, human BSMC from patients with asthma presented an up-regulation of miR-27b-3p expression compared with cells from healthy controls. Through bioinformatic analysis, the same study showed that miR-27b-3p participates in abnormal activation of the phosphatase and tensin homolog (PTEN) signaling, leading to the implementation of the growth program in cells from asthmatic patients [21].
On the other hand, in experimental studies, CD4+ Th2 cells from a mouse asthma model induced by OVA challenge showed an up-regulation in both miR-23b and miR-27b expression and a downregulation in both miR-106b and miR-203 in naïve CD4+ T cells [22]. The same study demonstrated that the inhibition (miR-23b and miR-27b) and overexpression (miR-106 and miR-203) simultaneously resulted in a progressive reduction in the Th2 phenotype by the reduction in IL-5, IL-9, and IL-13 production (Figure 1c) [22]. Therefore, the evidence suggests that miR-27b-3p could be important in asthma. However, to date, few studies have shown the role of miR-27b-3p in asthma. Hence, more experimental and clinical studies are needed to elucidate the function of miR-27b-3p in the asthma context.
Few studies reported that treating vitamin D3 reduces remodeling in asthma by regulating miR-27b. In experimental studies, for instance, in transforming growth factor (TGF) β1-stimulated human lung fibroblasts, the expression of miR-27b induces the differentiation to myofibroblast by negative regulation of VDR (Figure 1b) [23]. Likewise, treating 1,25(OH)2D3 in these cells inhibited its differentiation by the downregulation of miR-27b [23], suggesting that the effect of vitamin D3 on remodeling in asthma is mediated by the downregulation of miR-27b. In contrast, epithelial and splenocyte cells reported that the treatment with vitamin D3 increased the expression of miR-27b-3p [24][25]. The above evidence indicates that miR-27b plays an important role in the development of asthma. Likewise, the beneficial effects of the supplementation with vitamin D3 are mediated by the negative regulation of miR-27b. Hence, more studies need to elucidate the role of vitamin D3 on the regulation of miR-27b in improving asthma.
3. The Role of MicroRNA-145 in Asthma and Its Modulation by Vitamin D3
miR-145-5p is a member of the miR-143/145 cluster, which is involved in lipid metabolism and inflammatory pathways in a tissue-specific manner [26]. Moreover, miR-145-5p is a miRNA mainly considered a tumor suppressor in several cancers. However, it has also been shown to affect the pathogenesis of several diseases, including asthma [27]. Clinical studies have shown that asthmatic patients present high plasma levels of miR-145-5p compared with healthy controls [28][29] and that high levels of this miRNA correspond to a high count of eosinophils in blood [28]. A cross-sectional study conducted in children with several asthma phenotypes showed that the levels of miR-145-5p in exhaled breath condensate correlates positively with asthma [30]. Hence, these results could suggest that miR-145-5p may play a role in the development of asthma. In contrast, a study with asthmatic patients found low levels of miR-145-5p in plasma compared with healthy controls [28]. Moreover, a study conducted in children with asthma with reduced FEV1 values (both reduced growth and early decline) presented a high reduction in miR-145-5p levels in plasma compared with both children with only reduced growth and children with normal growth [31], while children with early decline showed an increase in miR-145-5p levels compared with children with reduced growth [31]. The same study showed that children who ended follow-up (when they were aged 23–30 years) developed COPD and those subjects presented a substantial reduction in levels of miR-145-5p in plasma compared with those whose non-developed COPD [31].
Experimental models of asthma have studied the role of miR-145-5p in asthma pathogenesis. Mouse models of asthma induced by challenge with house dust mite (HDM) found a strong up-regulation of miR-145-5p in the airway wall [32]. The same study also showed that the inhibition of miR-145-5p in asthmatic mice significantly reduced the development of airway hyperresponsiveness (reduced the number of both mucus-producing cells and eosinophils present in the airways) by the reduction in the production of IL-15 and IL-13 from antigen-specific Th2 cells; these effects were comparable with dexamethasone treatment [32]. Another study with a mouse model of asthma induced by HDM found an up-regulation of miR-145-5p expression and a down-regulation of kinesin Family Member 3A (KIF3A) in airway epithelial cells [33]. The same study found that the overexpression of miR-145-5p in bronchial epithelial cells promoted the secretion of chemokines and inflammatory factors and the dysfunction of the epithelial barrier by down-regulation of KIF3A [33]; its inhibition resulted in the improvement of symptoms and the inflammatory state due to an increase in the expression of KIF3A (Figure 2a) [33]. In contrast, an asthma model in mice induced by OVA showed that the expression of miR-145-5p led to less airway remodeling by regulation of EGFR compared with asthmatic mice with inhibited miR-145-5p [34]. Likewise, the negative regulation of EGFR by miR-145-5p promoted a reduction in both the proportion of Th2 and Th17 cells in blood and inflammatory factors in asthmatic mice compared with asthmatic mice with miR-145-5p inhibited [34]. Thus, in both human and murine models, evidence indicates that miR-145-5p participates in hallmark features of allergic airway disease.
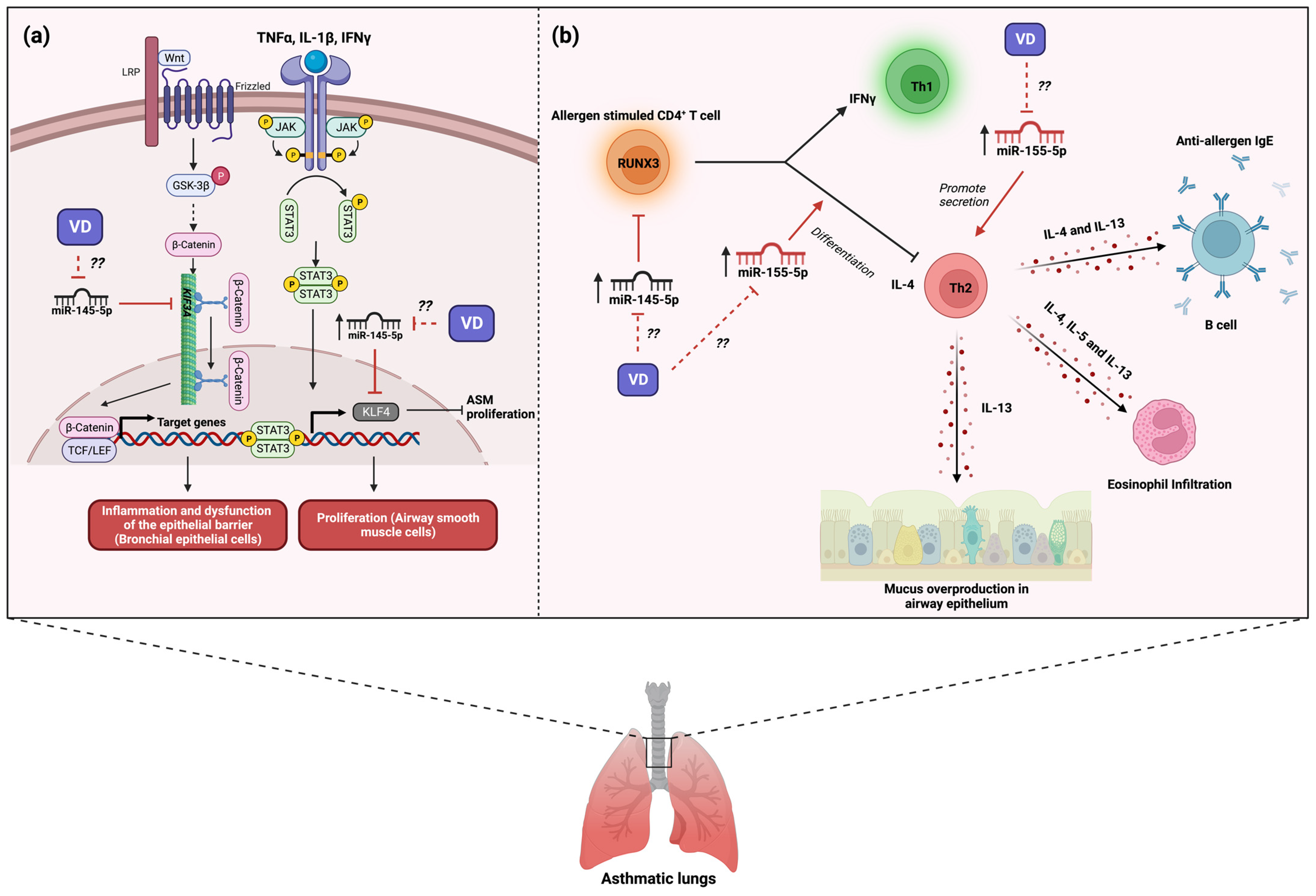
Figure 2. Role of miR-145-5p and miR-155-5p in the development of asthma and possible implication of vitamin D3 supplementation. (a) The expression of miR-145-5p in asthma is involved in the interference of the Wnt/β-catenin and cytokine signaling, leading to inflammation, dysfunction of bronchial epithelium, and the proliferation of airway smooth muscle. The question marks indicate that Vitamin D3 supplementation may alleviate the inflammation, avoiding airway epithelium dysfunction and smooth muscle proliferation by inhibiting miR-145-5p expression. (b) Likewise, both miR-145-5p and miR-155-5p induce Th2 differentiation, promoting the Th2 phenotype. The expression of miR-155-5p promotes the secretion of Th2-associated cytokines, which induce mucus overproduction, eosinophil infiltration, and anti-allergen IgE production. The question marks indicate that vitamin D3 supplementation may modulate the balance of Th1/Th2 cells through the regulation of miR-145-5p and miR-155-5p expression, potentially leading to the alleviation of asthma symptoms. The red lines illustrate the regulatory effects of both vitamin D3 and miRNAs. Created with BioRender.com.
At the cellular level, human ASM cells treated with TNF-α, IL-1β, and IFN-γ (imitate the inflammatory conditions in patients with asthma) induced an up-regulation of miR-145-5p and a down-regulation of Krüppel-like factor 4 (KLF4), an inhibitor of smooth muscle cell proliferation [35]. The same study showed that the negative regulation of KLF4 by overexpression of miR-145-5p enhanced the proliferation and migration of ASM cells in vitro [35], suggesting that this miRNA may participate in the smooth muscle remodeling present in asthma pathology (Figure 2a). Moreover, Qiu Yu-Ying and collaborators found that CD4+ T cells from patients with asthma presented both an up-regulation of miR-145-5p and a down-regulation of Runt-related transcription factor 3 (RUNX3) expression compared with healthy controls [36]. Given that RUNX3 plays a role in the maintenance of Th1/Th2 balance, they conducted an individual and simultaneous inhibition of miRNAs (miR-371, miR-138, miR-544, miR-145, and miR-214) and found that only simultaneous inhibition of miRNAs increased the mRNA and protein of RUNX3 [36]. Regarding the above study, Fan Linxia and collaborators showed that T CD4+ cells from patients with asthma presented an up-regulation in both miR-145-5p expression and IL-4 (Th2) expression and a down-regulation in both IFN-γ (Th1) and RUNX3 (Figure 2b) [37]. The inhibition of miR-145-5p in T CD4+ cells from patients with asthma promoted an enhanced percentage of IFN-γ+ CD4+ T cells by regulation of RUNX3 [37]; thus, miR-145-5p can modulate the Th1/th2 balance in asthma (Figure 2b). Therefore, all evidence suggests that miR-145-5p is an important miRNA in asthma development by regulating Th1/th2 balance and airway remodeling. However, to date, no clinical trials are driven to miR-145-5p regulation for asthma treatment.
Few studies reported a relationship between miR-145-5p and vitamin D status. In clinical studies, for instance, a recent study showed that obese and non-obese type 2 diabetes mellitus (T2DM) with deficiencies of vitamin D presented an up-regulation of miR-145-5p in plasma (5.91-fold) compared with those with insufficiency (4.61-fold) and sufficiency (4.29-fold) [38]. Moreover, recently, it was demonstrated that miR-145-5p is regulated by the vitamin D3 treatment in gastric cancer cells [39] and in aortic tissue [40]. Evidence suggests that the supplementation of vitamin D3 may induce the reduction in expression of miR-145-5p, improving asthma status. However, to date, there are no studies that indicate the above. Therefore, it is necessary to evaluate the effect of the supplementation of vitamin D3 on the expression regulation of miR-145-5p in asthma
4. The Role of MicroRNA-155 in Asthma and Its Modulation by Vitamin D3
MiR-155 is encoded by the miR host gene 155 (MIRHG155), identified initially as the B-cell integration cluster gene, which is a miRNA important for the immune response and is involved in several diseases, including asthma. A clinical study conducted on asthmatic children reported that those children presented high plasmatic levels of miR-155-5p compared to healthy children, which was associated with an increased risk of childhood asthma [41]. It is well known that indoor air pollution aggravates allergic asthma. The same study reported that the plasmatic levels of miR-155-5p positively correlated with particulate matter 2.5 and formaldehyde [41]. A recent study found that asthmatic patients presented high serum levels of miR-155-5p compared to healthy subjects [3]. Also, only asthma presence in these patients was a significant predictor of miR-155-5p expression [3]. A study conducted in asthmatic children reported that those children presented an up-regulation of miR-155-5p in plasma comparisons with healthy children, which increased with disease severity, along with increased IL-13 levels [42]. Furthermore, the same study reported that plasma levels of miR-155-5p positively correlated with plasma levels of IL-13 and negatively with lung functions (FEV1 and FVC) [42]. Therefore, evidence suggests that altered expression of miR-155-5p plays an important role in the development and exacerbation of asthma.
In experimental animals, miR-155-p has been found to participate in the inflammatory response of airway hyperresponsiveness and allergic airway inflammation. For instance, the lung tissue from a mouse asthma model found the up-regulation of miR-155-5p compared to control mice [43]. Likewise, the same study demonstrated that silencing of miR-155-5p reduced the levels of cytokines IL-4, IL-5, and IL-13 and the total eosinophil, macrophage, and lymphocyte count in BALF from asthmatic mice [43]. According to the above study, in a miR-155 KO mouse model of asthma, it was shown that a lack of miR-155 promotes a reduction in Th2 cells number and cytokines IL-4, IL-5, and IL-13, resulting in the decrease in eosinophil-mediated inflammation and mucus hypersecretion in the lung compared to wild-type asthmatic mice (Figure 2b) [44]. In another miR-155 KO mouse asthma model, it was observed that the lack of miR-155 promotes the reduction in reactive species oxygen (ROS) and cyclooxygenase 2 (COX-2) levels, Th2 and Th17-associated cytokines (IL-4, IL-13, and IL-17) levels in BALF, and in eosinophil infiltration [45]; the expression of miR-155 presented opposite results in the miR-155 KO mouse asthma model (Figure 2b) [45]. Moreover, a study with IL-4- and IL-13-treated macrophages stimulated with IL-33 (an asthma cellular model) induced an increase in the expression of miR-155-5p, along with an increase in inflammatory mediators (Ccl3, Ccl5, Ccl17, Ccl24, and Il1b) compared to macrophages without pretreatment [46]. Therefore, evidence suggests that the allergic inflammatory response is potentiated through the increase in the expression of miR-155-5p.
Experimental studies have reported that treating vitamin D3 improves the inflammatory response by regulating miR-155 expression. A previous study reported that LPS-pretreated macrophages stimulated with 1,25(OH)2D3 induced the downregulation of the expression of miR-155-5p via blocking NF-κB activation [47]. Moreover, it was also shown that 1,25(OH)2D3 treatment up-regulates SOCS1 by decreasing miR-155-5p expression [47]. Given that SOCS1 has an important role in the negative feedback in the inflammatory response, vitamin D3 limits the inflammatory response by promoting negative feedback action by downregulating miR-155-5p. DENV-infected monocyte-derived macrophages (MDM) treated with 1,25(OH)2D3 promoted the downregulation of TLR4 and reduced NF-kB activity, resulting in the decrease in miR-155-5p expression and IL-1β and TNFα production, along with the up-regulation of SOCS1 expression compared with DENV-infected cells not treated with MDM [48]. Another study in 1,25(OH)2D3-pretreated adipocytes stimulated with TNFα showed a decreased expression of miR-155-5p compared with non-pretreated adipocytes [49]. This downregulation of miR-155-5p was due to the pretreatment with 1,25(OH)2D3 reduced NF-κB activation by reducing phosphorylation levels of p65 and IκB [49]. Hence, the above evidence suggests that vitamin D3 supplementation could regulate the inflammatory response through the regulation of miR-155-5p. However, little is known about the beneficial effects of vitamin D3 on miR-155-5p expression in asthmatic patients. Therefore, studies to elucidate the anti-inflammatory effect of vitamin D3 through the regulation of miR-155 in asthma are needed.
5. Conclusions
In conclusion, miRNAs play an important role in the development and exacerbation of asthma. From emerging studies using the supplementation of vitamin D3, it has been shown that vitamin D3 can improve asthma symptoms. Evidence suggests that supplementation with vitamin D3 improves asthma by regulating miRNAs. Since miR-21, miR-27b, miR-145, and miR-155 are related to the disbalance of Th1/Th2 cells, inflammation, and airway remodeling, which worsens asthma symptoms, the effect of vitamin D3 on the regulation of these miRNAs in asthma must therefore be elucidated as a therapeutic strategy.
References
- Rostami Hir, S.; Alizadeh, Z.; Mazinani, M.; Mahlooji Rad, M.; Fazlollahi, M.R.; Kazemnejad, A.; Zavaran Hosseini, A.; Moin, M. Exosomal MicroRNAs as Biomarkers in Allergic Asthma. Iran. J. Allergy Asthma Immunol. 2021, 20, 160–168.
- Vázquez-Mera, S.; Martelo-Vidal, L.; Miguéns-Suárez, P.; Saavedra-Nieves, P.; Arias, P.; González-Fernández, C.; Mosteiro-Añón, M.; Corbacho-Abelaira, M.D.; Blanco-Aparicio, M.; Méndez-Brea, P.; et al. Serum Exosome Inflamma-MiRs Are Surrogate Biomarkers for Asthma Phenotype and Severity. Allergy 2023, 78, 141–155.
- ElKashef, S.M.M.A.E.; Ahmad, S.E.A.; Soliman, Y.M.A.; Mostafa, M.S. Role of MicroRNA-21 and MicroRNA-155 as Biomarkers for Bronchial Asthma. Innate Immun. 2021, 27, 61.
- Hammad Mahmoud Hammad, R.; Hamed, D.H.E.D.; Eldosoky, M.A.E.R.; Ahmad, A.A.E.S.; Osman, H.M.; Abd Elgalil, H.M.; Mahmoud Hassan, M.M. Plasma MicroRNA-21, MicroRNA-146a and IL-13 Expression in Asthmatic Children. Innate Immun. 2018, 24, 171–179.
- Joo, H.; Park, S.Y.; Park, S.Y.; Park, S.Y.; Kim, S.H.; Cho, Y.S.; Yoo, K.H.; Jung, K.S.; Rhee, C.K. Phenotype of Asthma-COPD Overlap in COPD and Severe Asthma Cohorts. J. Korean Med. Sci. 2022, 37, e236.
- Li, X.; Yang, N.; Cheng, Q.; Zhang, H.; Liu, F.; Shang, Y. Mir-21-5p in Macrophage-Derived Exosomes Targets Smad7 to Promote Epithelial Mesenchymal Transition of Airway Epithelial Cells. J. Asthma Allergy 2021, 14, 513–524.
- Liu, Y.; Yang, K.; Shi, H.; Xu, J.; Zhang, D.; Wu, Y.; Zhou, S.; Sun, X. MiR-21 Modulates Human Airway Smooth Muscle Cell Proliferation and Migration in Asthma through Regulation of PTEN Expression. Exp. Lung Res. 2015, 41, 535–545.
- Lee, H.Y.; Hur, J.; Kang, J.Y.; Rhee, C.K.; Lee, S.Y. MicroRNA-21 Inhibition Suppresses Alveolar M2 Macrophages in an Ovalbumin-Induced Allergic Asthma Mice Model. Allergy Asthma Immunol. Res. 2021, 13, 312.
- Zou, Y.; Zhou, Q.; Zhang, Y. MicroRNA-21 Released from Mast Cells-Derived Extracellular Vesicles Drives Asthma in Mice by Potentiating Airway Inflammation and Oxidative Stress. Am. J. Transl. Res. 2021, 13, 7475.
- Sheane, B.; Smyth, P.; Scott, K.; Aziz, R.; Buckley, M.; Lodge, E.; Kiely, N.; Kingston, M.; McGovern, E.; Healy, M.; et al. An Association between MicroRNA-21 Expression and Vitamin D Deficiency in Coronary Artery Disease. Microrna 2015, 4, 57–63.
- Grieco, G.E.; Cataldo, D.; Ceccarelli, E.; Nigi, L.; Catalano, G.; Brusco, N.; Mancarella, F.; Ventriglia, G.; Fondelli, C.; Guarino, E.; et al. Serum Levels of MiR-148a and MiR-21-5p Are Increased in Type 1 Diabetic Patients and Correlated with Markers of Bone Strength and Metabolism. Non-Coding RNA 2018, 4, 37.
- Xu, Y.; Qian, J.; Yu, Z. Budesonide Up-Regulates Vitamin D Receptor Expression in Human Bronchial Fibroblasts and Enhances the Inhibitory Effect of Calcitriol on Airway Remodeling. Allergol. Immunopathol. 2019, 47, 585–590.
- Liu, G.; Friggeri, A.; Yang, Y.; Milosevic, J.; Ding, Q.; Thannickal, V.J.; Kaminski, N.; Abraham, E. MiR-21 Mediates Fibrogenic Activation of Pulmonary Fibroblasts and Lung Fibrosis. J. Exp. Med. 2010, 207, 1589–1597.
- D’Onofrio, N.; Prattichizzo, F.; Martino, E.; Anastasio, C.; Mele, L.; La Grotta, R.; Sardu, C.; Ceriello, A.; Marfella, R.; Paolisso, G.; et al. MiR-27b Attenuates Mitochondrial Oxidative Stress and Inflammation in Endothelial Cells. Redox Biol. 2023, 62, 102681.
- Tang, Y.; Yang, L.J.; Liu, H.; Song, Y.J.; Yang, Q.Q.; Liu, Y.; Qian, S.W.; Tang, Q.Q. Exosomal MiR-27b-3p Secreted by Visceral Adipocytes Contributes to Endothelial Inflammation and Atherogenesis. Cell Rep. 2023, 42, 111948.
- Kho, A.T.; Sharma, S.; Davis, J.S.; Spina, J.; Howard, D.; McEnroy, K.; Moore, K.; Sylvia, J.; Qiu, W.; Weiss, S.T.; et al. Circulating MicroRNAs: Association with Lung Function in Asthma. PLoS ONE 2016, 11, e0157998.
- Dong, X.; Zhong, N.; Fang, Y.; Cai, Q.; Lu, M.; Lu, Q. MicroRNA 27b-3p Modulates SYK in Pediatric Asthma Induced by Dust Mites. Front. Pediatr. 2018, 6, 301.
- Macglashan, D.; Moore, G.; Muchhal, U. Regulation of IgE-Mediated Signalling in Human Basophils by CD32b and Its Role in Syk down-Regulation: Basic Mechanisms in Allergic Disease. Clin. Exp. Allergy 2014, 44, 713–723.
- Wang, M.; Liu, M.; Wang, C.; Xiao, Y.; An, T.; Zou, M.; Cheng, G. Association between Vitamin D Status and Asthma Control: A Meta-Analysis of Randomized Trials. Respir. Med. 2019, 150, 85–94.
- Coskunpinar, E.; Akcesme, B.; Tas, S.K.; Aynaci, A. Investigation of MiRNAs That Are Effective in the Pathogenesis of Asthma. J. Asthma 2023, 60, 2145–2152.
- Alexandrova, E.; Miglino, N.; Hashim, A.; Nassa, G.; Stellato, C.; Tamm, M.; Baty, F.; Brutsche, M.; Weisz, A.; Borger, P. Small RNA Profiling Reveals Deregulated Phosphatase and Tensin Homolog (PTEN)/Phosphoinositide 3-Kinase (PI3K)/Akt Pathway in Bronchial Smooth Muscle Cells from Asthmatic Patients. J. Allergy Clin. Immunol. 2016, 137, 58–67.
- Kılıç, A.; Santolini, M.; Nakano, T.; Schiller, M.; Teranishi, M.; Gellert, P.; Ponomareva, Y.; Braun, T.; Uchida, S.; Weiss, S.T.; et al. A Systems Immunology Approach Identifies the Collective Impact of 5 MiRs in Th2 Inflammation. JCI Insight 2018, 3, e97503.
- Li, F.; Zhang, A.; Shi, Y.; Ma, Y.; Du, Y. 1a,25-DihydroxyVitamin D3 Prevents the Differentiation of Human Lung Fibroblasts via MicroRNA-27b Targeting the Vitamin D Receptor. Int. J. Mol. Med. 2015, 36, 967–974.
- Kordkhayli, M.M.; Mansouri, F.; Talebi, F.; Noorbakhsh, F.; Saboor-Yaraghi, A.A. Influence of Vitamins A and D on the Expression of MicroRNA27-3p Isoforms and GATA3 in Experimental Autoimmune Encephalomyelitis. Iran. J. Allergy Asthma Immunol. 2022, 21, 429–440.
- Ge, X.; Yuan, L.; Wei, J.; Nguyen, T.; Tang, C.; Liao, W.; Li, R.; Yang, F.; Zhang, F.; Zhao, B.; et al. Vitamin D/VDR Signaling Induces MiR-27a/b Expression in Oral Lichen Planus. Sci. Rep. 2020, 10, 301.
- Quintanilha, B.J.; Reis, B.Z.; Corrêa, T.A.F.; Duarte, G.B.D.S.; Rogero, M.M. MicroRNAs and Inflammation Biomarkers in Obesity. In Precision Medicine for Investigators, Practitioners and Providers; Academic Press: Cambridge, MA, USA, 2020; pp. 179–185.
- Kadkhoda, S.; Ghafouri-Fard, S. Function of MiRNA-145–5p in the Pathogenesis of Human Disorders. Pathol. Res. Pract. 2022, 231, 153780.
- Panganiban, R.P.; Wang, Y.; Howrylak, J.; Chinchilli, V.M.; Craig, T.J.; August, A.; Ishmael, F.T. Circulating MicroRNAs as Biomarkers in Patients with Allergic Rhinitis and Asthma. J. Allergy Clin. Immunol. 2016, 137, 1423–1432.
- Lacedonia, D.; Palladino, G.P.; Foschino-Barbaro, M.P.; Scioscia, G.; Elisiana, G. Carpagnano Expression Profiling of MiRNA-145 and MiRNA-338 in Serum and Sputum of Patients with COPD, Asthma, and Asthma–COPD Overlap Syndrome Phenotype. Int. J. Chron. Obs. Pulmon. Dis. 2017, 12, 1811.
- Mendes, F.C.; Paciência, I.; Ferreira, A.C.; Martins, C.; Rufo, J.C.; Silva, D.; Cunha, P.; Farraia, M.; Moreira, P.; Delgado, L.; et al. Development and Validation of Exhaled Breath Condensate MicroRNAs to Identify and Endotype Asthma in Children. PLoS ONE 2019, 14, e0224983.
- Tiwari, A.; Li, J.; Kho, A.T.; Sun, M.; Lu, Q.; Weiss, S.T.; Tantisira, K.G.; McGeachie, M.J. COPD-Associated MiR-145-5p Is Downregulated in Early-Decline FEV1 Trajectories in Childhood Asthma. J. Allergy Clin. Immunol. 2021, 147, 2181–2190.
- Collison, A.; Mattes, J.; Plank, M.; Foster, P.S. Inhibition of House Dust Mite-Induced Allergic Airways Disease by Antagonism of MicroRNA-145 Is Comparable to Glucocorticoid Treatment. J. Allergy Clin. Immunol. 2011, 128, 160–167.e4.
- Xiong, T.; Du, Y.; Fu, Z.; Geng, G. MicroRNA-145-5p Promotes Asthma Pathogenesis by Inhibiting Kinesin Family Member 3A Expression in Mouse Airway Epithelial Cells. J. Int. Med. Res. 2019, 47, 3307.
- Cheng, Z.; Dai, L.L.; Wang, X.; Jia, L.Q.; Jing, X.G.; Li, P.F.; Liu, M.; Wang, H.; An, L. MicroRNA-145 down-Regulates Mucin 5AC to Alleviate Airway Remodeling and Targets EGFR to Inhibit Cytokine Expression. Oncotarget 2017, 8, 46312.
- Liu, Y.; Sun, X.; Wu, Y.; Fang, P.; Shi, H.; Xu, J.; Li, M. Effects of MiRNA-145 on Airway Smooth Muscle Cells Function. Mol. Cell Biochem. 2015, 409, 135–143.
- Qiu, Y.Y.; Zhang, Y.W.; Qian, X.F.; Bian, T. MiR-371, MiR-138, MiR-544, MiR-145, and MiR-214 Could Modulate Th1/Th2 Balance in Asthma through the Combinatorial Regulation of Runx3. Am. J. Transl. Res. 2017, 9, 3184.
- Fan, L.; Wang, X.; Fan, L.; Chen, Q.; Zhang, H.; Pan, H.; Xu, A.; Wang, H.; Yu, Y. MicroRNA-145 Influences the Balance of Th1/Th2 via Regulating RUNX3 in Asthma Patients. Exp. Lung Res. 2016, 42, 417–424.
- Aladel, A.; Khatoon, F.; Khan, M.I.; Alsheweir, A.; Almutairi, M.G.; Almutairi, S.O.; Almutairi, F.K.; Osmonaliev, K.; Beg, M.M.A. Evaluation of MiRNA-143 and MiRNA-145 Expression and Their Association with Vitamin-D Status Among Obese and Non-Obese Type-2 Diabetic Patients. J. Multidiscip. Healthc. 2022, 15, 2979.
- Chang, S.; Gao, L.; Yang, Y.; Tong, D.; Guo, B.; Liu, L.; Li, Z.; Song, T.; Huang, C. MiR-145 Mediates the Antiproliferative and Gene Regulatory Effects of Vitamin D3 by Directly Targeting E2F3 in Gastric Cancer Cells. Oncotarget 2015, 6, 7675–7685.
- Carrillo-lópez, N.; Panizo, S.; Arcidiacono, M.V.; de la Fuente, S.; Martínez-arias, L.; Ottaviano, E.; Ulloa, C.; Ruiz-torres, M.P.; Rodríguez, I.; Cannata-Andía, J.B.; et al. Vitamin D Treatment Prevents Uremia-Induced Reductions in Aortic MicroRNA-145 Attenuating Osteogenic Differentiation despite Hyperphosphatemia. Nutrients 2022, 14, 2589.
- Liu, Q.; Wang, W.; Jing, W. Indoor Air Pollution Aggravates Asthma in Chinese Children and Induces the Changes in Serum Level of MiR-155. Int. J. Environ. Health Res. 2018, 29, 22–30.
- Karam, R.A.; Abd Elrahman, D.M. Differential Expression of MiR-155 and Let-7a in the Plasma of Childhood Asthma: Potential Biomarkers for Diagnosis and Severity. Clin. Biochem. 2019, 68, 30–36.
- Chen, H.; Xu, X.; Cheng, S.; Xu, Y.; Xuefei, Q.; Cao, Y.; Xie, J.; Wang, C.Y.; Xu, Y.; Xiong, W. Small Interfering RNA Directed against MicroRNA-155 Delivered by a Lentiviral Vector Attenuates Asthmatic Features in a Mouse Model of Allergic Asthma. Exp. Ther. Med. 2017, 14, 4391.
- Malmhäll, C.; Alawieh, S.; Lu, Y.; Sjöstrand, M.; Bossios, A.; Eldh, M.; Rådinger, M. MicroRNA-155 Is Essential for TH2-Mediated Allergen-Induced Eosinophilic Inflammation in the Lung. J. Allergy Clin. Immunol. 2014, 133, 1429–1438.
- Qiu, L.; Zhang, Y.; Do, D.C.; Ke, X.; Zhang, S.; Lambert, K.; Kumar, S.; Hu, C.; Zhou, Y.; Ishmael, F.T.; et al. MiR-155 Modulates Cockroach Allergen and Oxidative Stress–Induced Cyclooxygenase-2 in Asthma. J. Immunol. 2018, 201, 916–929.
- Chia, N.; Kumar, R.K.; Foster, P.S.; Herbert, C. Enhanced Pro-Inflammatory Response of Macrophages to Interleukin-33 in an Allergic Environment. Int. Arch. Allergy Immunol. 2018, 176, 74–82.
- Chen, Y.; Liu, W.; Sun, T.; Huang, Y.; Wang, Y.; Deb, D.K.; Yoon, D.; Kong, J.; Thadhani, R.; Li, Y.C. 1,25-Dihydroxyvitamin D Promotes Negative Feedback Regulation of TLR Signaling via Targeting MicroRNA-155–SOCS1 in Macrophages. J. Immunol. 2013, 190, 3687–3695.
- Arboleda, J.F.; Fernandez, G.J.; Urcuqui-Inchima, S. Vitamin D-Mediated Attenuation of MiR-155 in Human Macrophages Infected with Dengue Virus: Implications for the Cytokine Response. Infect. Genet. Evol. 2019, 69, 12–21.
- Karkeni, E.; Bonnet, L.; Marcotorchino, J.; Tourniaire, F.; Astier, J.; Ye, J.; Landrier, J.F. Vitamin D Limits Inflammation-Linked MicroRNA Expression in Adipocytes in Vitro and in Vivo: A New Mechanism for the Regulation of Inflammation by Vitamin D. Epigenetics 2018, 13, 156–162.
More
Information
Subjects:
Immunology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
583
Revisions:
2 times
(View History)
Update Date:
31 Jan 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No