Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Masatoshi Hayashi | -- | 5105 | 2024-01-24 03:03:30 | | | |
| 2 | Camila Xu | Meta information modification | 5105 | 2024-01-24 03:15:10 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hayashi, M. Pathophysiology of Childhood-Onset Myasthenia. Encyclopedia. Available online: https://encyclopedia.pub/entry/54265 (accessed on 25 July 2026).
Hayashi M. Pathophysiology of Childhood-Onset Myasthenia. Encyclopedia. Available at: https://encyclopedia.pub/entry/54265. Accessed July 25, 2026.
Hayashi, Masatoshi. "Pathophysiology of Childhood-Onset Myasthenia" Encyclopedia, https://encyclopedia.pub/entry/54265 (accessed July 25, 2026).
Hayashi, M. (2024, January 24). Pathophysiology of Childhood-Onset Myasthenia. In Encyclopedia. https://encyclopedia.pub/entry/54265
Hayashi, Masatoshi. "Pathophysiology of Childhood-Onset Myasthenia." Encyclopedia. Web. 24 January, 2024.
Copy Citation
Myasthenia gravis (MG) is understood to be a neuromuscular disorder caused by an immune disturbance at the neuromuscular junction, which results in symptoms such as muscle weakness and fatigue.
autoimmunity
childhood-onset
genetic background
myasthenia gravis
neuromuscular junction
1. Introduction
Myasthenia gravis (MG) is understood to be a neuromuscular disorder caused by an immune disturbance at the neuromuscular junction, which results in symptoms such as muscle weakness and fatigue. In 1964, Elmqvist et al. reported abnormalities in miniature endplate potential (MEPP), which they identified as a pathophysiology of the neuromuscular junction [1]. In 1973, Jim Patrick et al. demonstrated that the immunization of rabbits with the acetylcholine receptor (AChR) can induce a myasthenic state similar to that in humans [2]; as expected from clinical findings by Simpson in 1960 [3], it became clear that MG is an autoimmune disease of the neuromuscular junction.
2. Pathophysiology of Myasthenic State
The pathophysiology of childhood MG, especially MG with post-pubertal onset, is basically similar to that of adults. In some cases, such as neonatal transient MG and congenital joint contractures, there is a pathophysiology specific to children, and there are problems with steroid administration and the timing of thymectomy during maturation. An understanding of the pathophysiology of MG in the growing pediatric population is desirable for selecting treatment. Myasthenia caused by impaired signaling at the neuromuscular junction can be broadly divided into acquired MG and congenital myasthenic syndrome (CMS). The pathophysiology, generally known as MG, refers to acquired MG, in which the AChR at the neuromuscular junction is reduced by immunological mechanisms [4]. Most often, this is caused by anti-AChR antibodies, which will be discussed next. It is said that complement is also involved in this process, which is accompanied by the morphological destruction of the neuromuscular junction. On the other hand, CMS is a genetic pathophysiology that is caused by a defect in the production of a protein involved in signal transduction at the neuromuscular junction.
2.1. Formation of Neuromuscular Junction
The neuromuscular junction must be well-formed for this neuromuscular signaling to be rapid and reliable. In denervated rat muscle, which was once used as an antigen in the measurement of AChR antibodies, denervation significantly alters the distribution of AChR in the postsynaptic membranes. There are two types of AChRs: the fetal type and the adult type. The adult type AChR consists of α, β, ε, and δ-subunit, while the fetal type consists of α, β, γ, and δ-subunit. Both are transmembrane proteins with an ion channel [5]. Surgical denervation and/or presynaptic blockade of neuromuscular transmission amplifies the subunit mRNA of junctional and extra-junctional AChR and increases both types of AChR in muscle cells [5]. Extra-junctional AChRs produced and distributed at the denervated post-synapses are reported to have γ-subunits instead of ε-subunits, as described above [6].
AChR proteins synthesized in muscle cells and extensively deployed and seeded in the postsynaptic membrane must be assembled at specific locations to form neuromuscular junctions. This requires stimulation from nerves, and a number of proteins are involved. (Figure 1) Nerve terminals secrete agrin, which binds to LRP4 and activates the MuSK molecule. Furthermore, the complex forms a dimer and activates Dok-7 in muscle cells. When the AChE/ColQ protein binds to the MuSK protein molecule, the complex is further activated and stabilized, and AChRs that were widely and thinly distributed in the surrounding area are gathered in the vicinity of MuSK to form neuromuscular junctions [7][8][9].
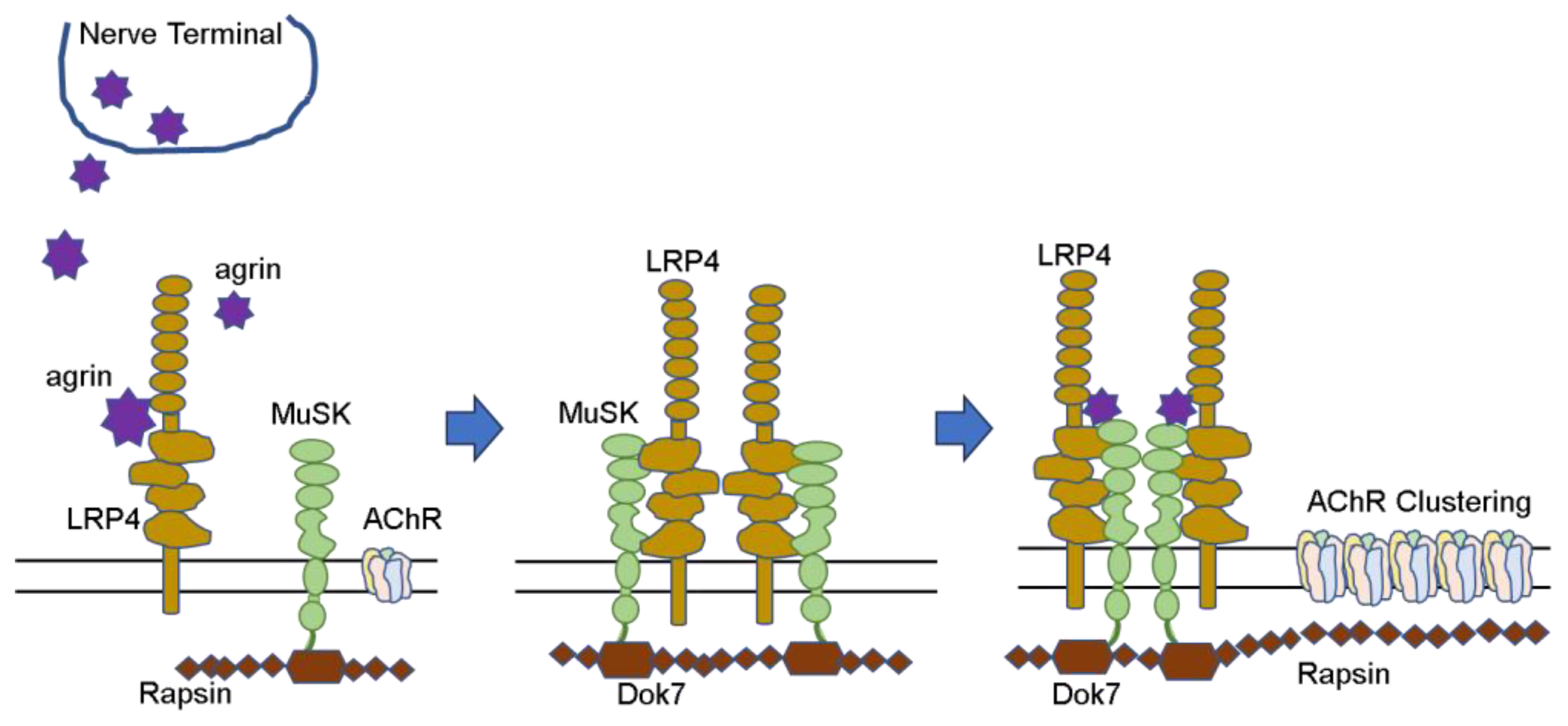
Figure 1. AChR clustering (Hayashi. No To Hattatsu 2022. [10]).
In addition to these, several other proteins have been reported to be involved in the formation of this AChR group. When these proteins malfunction, neuromuscular communication is impaired. Mutations that prevent the formation of normal protein molecules and, thus, impair neuromuscular communication are called CMS. The so-called acquired MG is mainly caused by autoantibodies that disrupt signal transduction at the neuromuscular junction, resulting in MG symptoms. The AChR and MuSK antibodies can be clearly identified as the causes of MG symptoms because the method of measurement is well established, the antibodies have been proven in many patient sera, and the same condition can be reproduced in animals by passive transfer. Recently, Lrp4 and agrin antibodies have been added to this group. Autoantibodies against various other proteins involved in neuromuscular junction formation are still under investigation. Sooner or later, it may become possible to identify genetic abnormalities associated with such proteins and to identify autoantibodies against them in patient sera.
2.2. Changes at the Neuromuscular Junction
2.2.1. Decreased AChR at the Neuromuscular Junction
Synapses at the neuromuscular junctions are inherently narrow. Degranulated acetylcholine molecules diffuse to the postsynaptic membrane on the opposite muscle side and then bind to AChRs present at the postsynaptic membrane. In 1973, Fambrough et al. took muscle biopsies of eight MG patients and measured the AChR density around the neuromuscular junction [4]. They reported that the AChR density was reduced by 11–30% in MG compared to normal subjects. In the same year, Patrick et al. demonstrated that this disease was an autoimmune disease by immunizing rabbits several times with AChR extracted from the electric organ of Torpedo and reproducing pathophysiology with symptoms of muscle weakness similar to those in humans [2]. In 1974, Almon et al. reported that the binding activity of α-bungarotoxin to AChR was decreased in at least 5 of the 15 MG patients when patient serum was added to AChR extracted from rat leg muscles [11]. In 1975, Bender et al. morphologically demonstrated that α-bungarotoxin bound to the muscle tissue of MG is eliminated by reacting to patient serum [12]. This indicates that the AChRs present in the postsynaptic membranes react with the patient’s serum, causing their density to decrease at the MG patient’s own neuromuscular junction. Thus, a series of studies on the pathogenesis of MG was published in the mid-1970s, establishing that MG is an autoimmune disease caused by antibodies in the patient’s blood acting to disrupt signaling at the neuromuscular junction.
2.2.2. Structural Destruction of the Neuromuscular Junction
Engel et al. reported that normal neuromuscular junctions have narrow synaptic clefts and well-constructed synaptic folds. In contrast, in MG, the synaptic cleft is enlarged, the synaptic folds disappear, and the debris floats in the enlarged synaptic cleft [13]. This is caused by complement reactions to the autoantibodies described below.
2.3. Autoantibodies against the Neuromuscular Junction
In acquired MG, autoantibodies against the neuromuscular junction are formed, which attack the neuromuscular junction, resulting in impaired neuromuscular communication and muscle weakness. AChR is a membrane protein with a molecular weight of approximately 290,000 that is present on the surface of muscle cells, spans the membrane, and is composed of five subunits of four types: two α, β, γ (or ε), and δ subunits. More than half of the autoantibodies present in patient sera are antibodies that target what is called the main immunogenic region, the α subunit of AChR [14].
There are three possible mechanisms by which AChR antibodies cause the reductions in AChR on postsynaptic membranes at the neuromuscular junction in MG, involving binding antibodies, blocking antibodies, and complements [15].
2.3.1. Involvement of AChR Antibodies
First, binding antibodies often recognize the α-subunit of AChR, and the Fab portion of the antibody binds to the AChR and bridges the two AChRs, thereby accelerating their uptake into muscle cells and increasing their decay rates [15][16][17]. The ratio of decayed and lost AChRs to newly produced AChRs determines how much density of AChRs is represented on the muscle cell surface. Generally, AChR antibodies are binding antibodies.
Second, the blocking antibodies recognize and bind to the ACh binding site or its vicinity in the α-subunit of AChR. This prevents ACh from binding to AChR [15][18][19]. The α-subunit is a linear protein consisting of 437 amino acids, each of which has various electric charges and, thus, assumes a three-dimensional structure that penetrates the muscle cell membrane four times. The N-terminal 210 amino acids are completely outside the membrane, and Ach, with a molecular weight of 146, and antibodies, with a molecular weight of approximately 150,000, react with that part of the subunit. AChR is composed of two α-subunits and one each of β, γ (or ε), and δ. The ACh binding sites are located at the contact sites of the α/γ (or α/ε) and α/δ subunits [20].
Third, there is a mechanism by which the complement acts to disrupt the morphology of the neuromuscular junction [13][21]. The morphological changes at the neuromuscular junction caused by the complement are characterized by a wide synaptic gap and a coarse distribution of AChRs, which allows for little information exchange [22]. When the complement repeats the reaction up to C9, it forms a membrane attack complex (MAC) and destroys the membrane [21]. When the presence of the complement is confirmed at the neuromuscular junction [23], and the complement component C3 is removed using snake venom, this synaptic destruction is no longer seen and symptoms improve [24].
2.3.2. Neonatal Transient Myasthenia Gravis and Fetal Myasthenia
Some infants born to MG mothers develop neonatal transient MG [25][26][27][28]. IgG antibodies are also transferred to the fetus via the placenta except for subclass IgG2, whereas most AChR antibodies are IgG1 and IgG3. Therefore, if the mother’s antibody titer is high, the antibodies are naturally transferred to the fetus, and symptoms should appear. However, only 10–12% of newborns develop MG, and most are asymptomatic despite having AChR antibodies in their blood [29]. Idiopathic thrombocytopenic purpura, in which maternal autoantibodies are similarly transferred to the fetus and cause postnatal symptoms, also occurs at a frequency of 20% [30] to 17.8% [31]. However, it is not well understood why neonates do not develop the disease.
This neonatal transient MG declines over time because it does not occur due to antibodies produced by the child on its own but to the transplacental transfer of maternal antibodies. Even if symptoms appear in the first few days of life, they gradually subside with continued treatment and management during the weeks of symptoms.
In rare cases, if the maternal antibodies are specific, transfer to the fetus in high concentrations, and are strong enough to inhibit movement in utero, the fetus may develop a pathophysiology called congenital joint contracture (arthrogryposis congenita) [32][33]. There exist antibodies against the γ-subunit of fetal AChR, which inhibit the response of ACh to its receptor, slow down the ion channel response, and impair signal transduction at the neuromuscular junction. Moreover, a monoclonal antibody (mAb 131) against fetal AChR with such a function has been reported [34].
2.3.3. Antibody against Muscle-Specific Tyrosine Kinase (MuSK)
When AChR antibodies are negative, the disease is called seronegative MG. In 2001, Hoch et al. reported that MuSK antibodies are present in 70% of seronegative MG, which accounts for 20% of generalized MG [35]. McConville et al., in the same group, reported that in 66 patients with seronegative MG, 27 (41%) were positive for MuSK antibodies, 11 of whom had prominent bulbar symptoms [36]. Several subsequent reports have shown that about 20% of Caucasian MG patients have seronegative MG, of which 30–40% of the generalized type are positive for MuSK antibodies [37][38]; 38% of adult seronegative MG cases were reported to be positive for MuSK antibodies by the Mayo Clinic [39]. On the other hand, a survey of adult MG in Asia showed that 21% of generalized type seronegative MG in South Asia [40], 26.4% in China [41], and 26.7% in Korea [42] were positive for the MuSK antibody.
In Japan, Ohta et al. reported that 27% of 85 patients with generalized type seronegative MG had MuSK-MG [43], but the epidemiology of pediatric MuSK-MG is poorly investigated, as only case reports of pediatric cases can be found [44][45]. Thus, subtle differences in the frequency of the disease exist according to region and race.
However, in a report looking at the relationship between MuSK-MG and HLA, Niks et al. reported that MuSK was correlated with HLA-DR14-DQ5 in Dutch people [46], and Kanai et al. reported that HLA-DRB1*14 and DQB1*05 were correlated in Japanese people [47]. Subsequently, Hong et al. reported that HLA-DQB1*05, DRB1*14, and DRB1*16 were correlated in a meta-analysis including the reports of Niks and Kanai, indicating that common HLA types are involved beyond racial differences [48].
The subclass of MuSK antibodies consists mainly of IgG4, which, like AChR antibodies, crosses the placenta to cause symptoms. Neonatal transient MG has been reported from mothers with MuSK-MG [49]. It is rare in MuSK-MG in the ocular muscle type [50][51].
MuSK is responsible for the assembly of AChRs on the postsynaptic membrane in collaboration with several protein molecules to facilitate efficient signal transduction at the neuromuscular junction. (Figure 1) MuSK antibodies have an adverse effect on AChR assembly and reduce functional AChRs, but histopathology does not confirm the loss of AChRs [52]. While the main subclasses of AChR antibodies are IgG1 and IgG3, MuSK antibodies are mainly the IgG4 subclass and IgG monovalent, which crosses the placenta but has no complement binding properties. Konectzny et al. examined the sera of 14 MuSK-MG patients and found that the MuSK antibodies were predominantly IgG4 monovalent with some IgG1-3. MuSK antibodies do not cause the intracellular uptake of MuSKs but instead impair agrin-induced AChR assembly, resulting in MG symptoms [53][54].
The most common sites of symptoms of MuSK-MG are the face, neck, articulatory swallowing, and respiratory muscles, where muscle weakness and atrophy occur. However, it has been suggested that the neuromuscular junction structure of these muscles may be different from that of others or that the low expression of MuSK in the scapulohyoid muscle may result in different responses [55]. MuSK-MG presents with bulbar symptoms, which may become more severe with anticholinesterase agents, whereas the thymus gland is normal, and therefore, thymectomy is not generally performed. Thus, the pathophysiologies of MuSK-MG and AChR antibody-positive MG are different.
MuSK, a protein present at the neuromuscular junction, is essential in order for AChRs to assemble at the neuromuscular junction; when MuSK is deficient or when antibodies block MuSK’s natural function, AChRs fail to form clusters, resulting in the inefficient transmission of information from the nerve and the development of MG pathophysiology. Subsequently, several antibodies other than AChR and MuSK antibodies have been shown to cause MG.
2.3.4. Double or Triple Seronegative MG
When the AChR antibody is negative, it is called seronegative MG, but when both the AChR and MuSK antibodies are negative, the name “double seronegative MG” is used. When the LRP4 antibody is also negative, “triple seronegative MG” is used. Rodriguez Cruz et al. reported that of 42 MG patients considered to be double seronegative by the immunoprecipitation method, 16 (38.1%) were positive for AChR antibodies according to cell-based assay [56]. In order to test negative for AChR antibodies, it may be necessary to confirm the result not only by immunoprecipitation but also by cell-based assay. Recently, a study has been reported to measure MuSK antibodies using a cell-based assay [57], but this is still in the research stage and needs to be accumulated in the future. In the same paper reported by Rodriguez Cruz, 26 patients were also considered negative by the cell-based assay method, and LRP4 antibodies were negative in all 21 patients who could be tested [56]. However, Pevzner et al. reported that serum from 12 of 13 double-negative MG patients showed protein deposition at the neuromuscular junction in mice, and in 4 patients, AChR assembly on cultured muscle cells was suppressed by more than 50% [58]. Higuchi et al. reported that 9 of 300 AChR antibody-negative patients had Lrp4 antibodies, and 3 of these 9 patients were also positive for MuSK antibodies [59]. LRP4 antibody-positive MGs are present among double seronegative MGs, but they vary from 2–45% depending on the region [60][61].
Recently, antibodies against agrin have also been investigated, and agrin antibodies have been detected in double and triple seronegative MGs by measuring AChR, MuSK, and LRP4 antibodies [62][63][64][65]. LRP4 and agrin, together with MuSK, play a major role in the formation of AChR clusters. The presence of these antibodies prevents the complex formation of MuSK and LRP4, which in turn inhibits AChR assembly and disrupts signaling at the neuromuscular junction.
In general, four conditions are needed to determine whether autoantibodies are responsible for the disease [66][67]. Regarding AChR antibodies, all of these conditions are satisfied: (1) antibodies can be identified in patient sera, (2) the passive immunization of patient sera causes characteristic pathophysiology, (3) the active immunization with antigens causes disease, and (4) the removal of antibodies improves symptoms.
Looking at antibodies against MuSK, LRP4, and agrin for conditions (1) through (3), Shigemoto et al. induced MG symptoms in rabbits by immunizing them with MuSK protein. Pathophysiology showed a reduction in AChR clusters at the neuromuscular junction [68], while Viegas et al. created a MuSK-MG pathophysiology in mice by active immunization with the MuSK protein as well as passive immunization with the MuSK antibody [69].
Similarly, Shen et al. actively immunized mice with the extracellular domain of the LRP protein to induce MG symptoms and passively immunized mice with serum from rabbits immunized with the LRP4 protein to induce the same MG symptoms [61]. Ulsoy et al. [70] and Mori et al. [71] also observed MG symptoms in mice immunized with LRP4 and created autoimmune animals. LRP4 antibodies belong mainly to IgG1 and have complement activity [59].
Yu et al. suppressed MuSK phosphorylation and AChR assembly by passively immunizing mice with immunoglobulin generated from MG patient sera with LRP4/agrin antibodies [72], and Yan et al. immunized mice with agrin to induce the development of MG symptoms [64].
The pathophysiology of acquired MG is thought to be caused by the formation of autoantibodies against protein substances at the neuromuscular junction. The autoantibodies that are currently recognized are against AChR, MuSK, Lrp4, and agrin. All four auto-antibodies appear to meet all of the strict criteria.
There are many other proteins involved in AChR assembly, and several antibodies against these proteins have been identified. It remains to be verified whether these antibodies are really involved in the pathogenesis of the disease.
2.4. Congenital Myasthenic Syndrome (CMS)
CMS is characterized by pathological muscle weakness and fatigability caused by an inborn defect of a protein molecule at the neuromuscular junction and is usually diagnosed at the age of 2 years or younger. In addition, CMS is often associated with muscle atrophy and small deformities, so it is important to distinguish CMS from MG as well as from muscular dystrophy and congenital myopathy. Repeated nerve stimulation is essential for definitive diagnosis, and careful and repeated nerve stimulation is desirable in cases of muscle weakness and atrophy without elevated CK [73].
A report from the U.K. showed a high prevalence of 1.5 per million for childhood-onset autoimmune MG in those aged 18 years or younger, compared with 9.2 per million for CMS [74]. The Mayo Clinic reported an incidence rate of 1.2 per million for autoimmune MG and 2.3 per million for CMS in childhood-onset MG under 19 years of age [75]. No epidemiologic reports of CMS have been seen from Asia, only sporadic reports. From Japan, Azuma et al. reported 4 patients with ColQ abnormalities and 5 of their mutations and 5 patients with AChR abnormalities and 6 of their mutations, for a total of 9 patients and 11 of their mutations; however, underdiagnosis is considered highly likely [76]. Thus, to compare the frequency of childhood MG between Europe and East Asia, autoimmune MG is more common in East Asia and less common in Europe and the United States. Conversely, CMS shows a contrasting pattern of onset, being more common in the West and less common in East Asia, with racial differences also present.
CMS is a phenomenon caused by genetic abnormalities in various proteins at the neuromuscular junction that are necessary for neuromuscular signaling to occur, resulting in impaired protein synthesis. Ohno and others have recently reported a review of 35 different genetic abnormalities [77]. The final stage of AChR assembly may involve other unknown proteins, and further studies are needed.
2.5. Disease Classification: Ocular and Generalized MG
MG can be broadly classified into ocular MG and generalized MG. In children, ocular MG is common, accounting for 78.2% of cases, and is also known to have low AChR antibody titers and a high percentage of negative titers [78]. In a 2006 epidemiological survey from Japan, 80.6% of cases with onset at less than 5 years of age and 61.5% of cases with onset at 5 to 10 years of age were ocular MG [79]. Sero-negative MG is more common in pediatric patients with ocular MG [78]. Whether or not antibodies are truly absent is a major issue, and Tsujihata et al. confirmed the deposition of anti-AChR antibodies at the neuromuscular junction of the limb muscles by performing limb muscle biopsies of patients with ocular MG [23]. Patients who were thought to have seronegative MG were revealed to have seropositive MG, as antibody titers were detectable against cell-bound AChR due to differences in assay methods [56][80]. In Japan, Oda reported in 1993 that the cell-bound AChR assay using human ocular muscle as an antigen could identify antibodies in ocular MG serum that were negative by means of the radioimmunoprecipitation method [81].
Since the pathophysiology of MG is thought to be caused by autoantibodies, it is thought that even when symptoms are present only in the ocular muscles, antibodies are deposited in other muscles throughout the body, although they are not yet developed due to a threshold for onset. The AChR antibody deposition at the limb muscle’s neuromuscular junction, shown above (by Tsujihata), illustrates this idea [23].
In East Asia, ocular MG is more common in children, and most adults also present with ocular muscle symptoms and later develop the generalized type. Why is the ocular MG more common, and why is the ocular muscle more likely to be affected [54][82]? Comparing the neuromuscular junction of the limb muscles with that of the ocular muscles, there are several electrophysiological differences. The ocular muscles require rapid and complex movements of the eyeballs, as well as fixation. Furthermore, the AChR subunits are different, and it is known that ε is replaced by γ in the AChR subunit of the ocular muscle, whereas the limb muscle AChRs originally consisted of α, β, ε, and δ [83]. The γ-subunit forms the fetal AChR, and in rodents, many muscles are replaced by the epsilon form within the first week or so after birth [84]. However, this replacement does not occur in the ocular muscles. It has been reported that the neuromuscular junction of ocular muscles has a more complex morphology than that of limb muscles [85], with ε-subunits in simple innervated neuromuscular junctions such as the soleus muscle and γ-subunits in multiple innervated neuromuscular junctions of the external ocular muscle [84][85][86]. These differences in the neuromuscular junction accommodate the fine and complex movements of the eye, and the absolute differences in muscle size and thickness create the so-called “safety factor”, or margin of safety, which explains why the ocular muscles with less margin are more prone to symptoms. It has been explained that symptoms are more likely to occur in the ocular muscles with less room to spare [87][88].
2.6. Childhood Thymus and Thymic Selection
MG is often associated with thymic abnormalities such as thymoma and thymic hyperplasia. Without distinguishing between thymoma and thymic hyperplasia, Yoshikawa et al. reported that 22.1% of cases were thymoma in Japanese patients [89], and the same was true for 20.1% of Korean patients [90]. Murai et al. reported 32.0% as thymoma and 38.4% as thymic hyperplasia. In MG in children under 9 years of age, thymoma comprised 4.9%, and thymic hyperplasia was 16.8% less common than in adults [79]. Similarly, in China, thymoma and thymic hyperplasia comprised 14.8% and 66.4%, respectively, while thymoma made up 2.9% and thymic hyperplasia 86.5% in pediatric MG cases aged 14 years or younger [91]. Popperud et al. reported that 50 of 63 pediatric MG patients under the age of 18 had undergone thymectomy. Of the 21 prepubertal patients, 13 had thymectomy and 7 (54%) had thymic hyperplasia, and of the 42 postpubertal patients, 37 had thymectomy and 23 (62%) had thymic hyperplasia. No thymoma was reported either before or after puberty [92], and Heckman et al. also reported that thymoma is rare in children [93]. Thus, the critical difference is whether the thymus is a tumor or hyperplasia in both children and adults. If it is a thymoma, removal is the treatment of choice.
The thymus gland is an organ that all humans are born with, and it plays a major role in the development of immunity that is able to distinguish between self and non-self [94][95].
The thymus gland usually atrophies after completing its work of establishing immunity at a young age, and most disappear by about age 40 [94][96][97]. Therefore, if thymectomy is indicated in children, it should be performed after puberty. Thymoma, which occurs more frequently in adults, has a different implication. Since the thymus gland, which should be atrophied, is instead enlarged and harms the person as a tumor, its removal is considered to be the appropriate treatment.
2.7. Involvement of Cellular Immunity
The involvement of HLA was mentioned above as an influence of one’s immunogenetic background. In particular, HLA-DR is MHC-class II, which expresses antigen peptides bound to its pocket on the surfaces of antigen-presenting cells and reacts with the TCR of T cells. As an experimental model, Berman and Patrick examined the murine system using various types of mice [98]. The results showed that C57BL/6(B6), which is Th1-dominant, is used as an experimental model for MG instead of Balb/c, which is Th2-dominant. The mouse MHC, H-2 complex, is haplotype-b in B6 and haplotype-d in Balb/c. Christadoss et al. have shown that the differences in MG-susceptibility between species of mice are due to differences in T-cell activity in response to the same antigenic stimuli and that the MG-susceptibility of the H-2 gene is linked and that it is controlled by the I-A subregion of the MHC linked to the H-2 gene [99].
McIntyre and Seidman have created a B6.C-H-2bm12 (bm12) mouse with three mutations in the I-Aβ of the B6 mouse [100]. This bm12 mouse, with only three mutations in I-A, had a greatly reduced incidence of MG development [101]. This is reported to be due to a significant effect on the epitope repertoire of murine CD4+ T-cells sensitized to AChR [102][103]. Wu et al. created systemic MG by immunizing HLA-DR3 transgenic mice with human AChR epsilon-subunit [104].
Chronic myeloid leukemia (CML) treatment with intense chemotherapy and radiotherapy may result in thymic damage, which may lead to the release of abnormal lymphocytes into the peripheral circulation and the development of autoimmunity. Graft-versus-host disease (GVHD) is said to be caused by an immune imbalance due to decreased Tregs [105].
MG is also more likely to occur when autoimmune diseases are present in the family. The frequency of MG increases markedly in twins [106]. These events may indicate that MHC antigens expressed on antigen-presenting cells are transmitted within the family, making the family susceptible not only to MG but also to other autoimmune diseases.
MG is a T-cell-dependent, antibody-producing autoimmune disease. As such, treatment has included steroids, immunosuppressive agents, and, more recently, various biological agents, which are more easily implemented in treatment and more effective. However, the use of such drugs in children is limited by the difficulty of conducting clinical trials to confirm their safety [107][108]. Immunosuppressive agents such as azathioprine and cyclophosphamide have been used for severe cases in Japan, but they are not actively used because they are not covered by insurance. Currently, tacrolimus and ciclosporin A are the two calcineurin inhibitors that are covered by insurance, and eculizumab, a biologic, has recently become available for use in pediatric patients. Eculizumab is a C5 inhibitor of complement that prevents MAC (membrane attack complex) formation and the destruction of neuromuscular junctions. The calcineurin inhibitor, on the other hand, is quite effective, although it does not act directly on antibody production but rather suppresses T-cell activity upstream of it.
2.8. Lymphorrhage
MG is a T-cell-dependent disease in which antibody production occurs and neuromuscular signaling is disturbed. Autoantibodies against various proteins involved in AChR assembly, mainly AChR antibodies, are thought to be involved in this pathophysiology, including anti-MuSK, anti-LRP4, and anti-agrin antibodies. However, seronegative MG actually exists and accounts for 10% of generalized MG [109]. Oda fixed ocular muscle AChR to wells and measured AChR antibody titers in the serum of patients with ocular MG but reported that some cases were still negative [81]. A comparison of soluble IL2 receptor (sIL2R), a marker of T-cell activation, with AChR antibody titers showed a significant negative correlation, with lower antibody titers resulting in higher sIL2R [110]. This result suggests that T-cells may be activated in MG cases with low AChR antibody titers. In ocular MG, antibody titers are low, and T-cells may be activated. In cases where a patient is diagnosed with seronegative MG and no antibody is found, no matter the activity of the known antibodies, are there still antibodies that have not been found? Or are they just antibodies that are below the sensitivity of measurement? Lymphorrhage used to be a major issue in the days when many MG patients died and were autopsied [111], but since then, there has been controversy regarding the existence and pathological significance of lymphorrhage. In 1963, Fenichel et al. performed muscle biopsies on 37 MG patients and divided the tissues into three groups: 15 cases in the normal group, 11 cases in the small muscle group showing denervated muscle, and 12 cases in the lymphocytic infiltration group. Most of the biopsied muscles were quadriceps muscles, and the time from onset to biopsy was 1 to 8 years in most cases. Lymphocytic infiltration was seen at a high rate in muscle biopsies as well as autopsies and was more common in cases with a short period of time after onset and thymic abnormalities [112][113]. Pascuzzi and Campa biopsied the tricep muscles and found lymphorrhage in the muscle endplate, pointing to the possibility that cellular immunity is involved in the pathogenesis of MG [114]. Furthermore, Maselli et al. biopsied the anconeus muscles of eight MG patients and found cellular infiltration of the neuromuscular junction in seven patients [115]. On the other hand, Nakano et al. found inflammatory cell infiltration around the endplate in 12 of 30 patients, but the degree was mild, less than 10% of the endplate. They claimed that lymphorrhage was a nonspecific phenomenon [116]. Most of the muscle biopsy sites were external intercostal muscles.
In studies producing MG pathology in experimental animals, infiltration of lymphocytes around the neuromuscular junction has been seen in the acute phase, which occurs after antigen injection in rats. In the chronic phase, when AChR antibody titers rise, this lymphocytic infiltration disappears and morphological destruction of the neuromuscular junction is observed [117]. The same phenomenon is seen in the immunization of extracted and purified AChR [117], as well as in the passive transfer of AChR antibodies in serum [118][119][120].
In the mid-20th century, steroids began to be used, and a dramatic change in treatment occurred. Until then, there was no treatment available to deal with the rapidly progressing pathology of MG, so muscle tissue was viewed during autopsy at the end of the natural course of the disease. The literature of the time recorded the pathological findings in detail. The advent of anti-cholinesterase agents, ACTH, and thymectomy extended the clinical course of the disease. The use of steroids has saved many lives, deaths from MG have almost disappeared, and muscle tissue specimens are often obtained from localized muscle tissue such as the pectoralis major and intercostal muscles at the time of thymectomy. As studies using experimental models have shown, the pathophysiology is variable. The scene of the pathophysiology presented by the patient may differ depending on the time since the onset of the disease, and the judgment of the pathophysiology may differ depending on the object being viewed. The classic literature has repeatedly shown the importance of lymphorrhage.
References
- Elmqvist, D.; Hofmann, W.W.; Kugelberg, J.; Quastel, D.M.L. An electrophysiological investigation of neuromuscular transmission in myasthenia gravis. J. Physiol. 1964, 174, 417–434.
- Patrick, J.; Lindstrom, J.M. Autoimmune response to acetylcholine receptor. Science 1973, 180, 871–872.
- Simpson, J.A. Myasthenia gravis: A new hypothesis. Scott. Med. J. 1960, 5, 419.
- Fambrough, D.M.; Drachman, D.B.; Satyamurti, S. Neuromuscular junction in myasthenia gravis: Decreased acetylcholine receptors. Science 1973, 182, 293–296.
- Koneczny, I.; Herbst, R. Myasthenia gravis: Pathogenic effects of autoantibodies on neuromuscular architecture. Cells 2019, 8, 671.
- Witzemann, V.; Brenner, H.-R.; Sakmann, B. Neural factors regulate AChR submit mRNAs at rat neuromuscular synapses. J. Cell Biol. 1991, 114, 125–141.
- Takamori, M. Myasthenia gravis: From the viewpoint of pathogenicity focusing on acetylcholine receptor clustering, trans-synaptic homeostasis and synaptic stability. Front. Mol. Neurosci. 2020, 13, 86.
- Rodriguez Cruz, P.M.; Cossins, J.; Beeson, D.; Vincent, A. The neuromuscular junction in health and disease: Molecular mechanisms governing synaptic formation and homeostasis. Front. Mol. Neurosci. 2020, 13, 610964.
- Ohkawara, B.; Ito, M.; Ohno, K. Secreted signaling molecules at the neuromuscular junction in physiology and pathology. Int. J. Mol. Sci. 2021, 22, 2455.
- Hayashi, M. Childhood-onset myasthenia gravis. No Hattatsu 2022, 54, 235–242. (In Japanese)
- Almon, R.R.; Andrew, C.G.; Appel, S.H. Serum globulin in myasthenia gravis: Inhibition of a-bungarotoxin binding to acetylcholine receptors. Science 1974, 186, 55–57.
- Bender, A.N.; Ringel, S.P.; Engel, W.K.; Daniels, M.P.; Vogel, Z. Myasthenia gravis: A serum factor blocking acetylcholine receptors of the human neuromuscular junction. Lancet 1975, 1, 607–608.
- Engel, A.G.; Lindstrom, J.M.; Lambert, E.H.; Lennon, V.A. Ultrastructural localization of the acetylcholine receptor in myasthenia gravis and in its experimental autoimmune model. Neurology 1977, 27, 307–315.
- Tzartos, S.J.; Seybold, M.E.; Lindstrom, M. Specificities of antibodies to acetylcholine receptors in sera from myasthenia gravis patients measured by monoclonal antibodies. Proc. Natl. Acad. Sci. USA 1982, 79, 188–192.
- Conti-Fine, B.M.; Milani, M.; Kaminski, H.J. Myasthenia gravis: Past, present, and future. J. Clin. Investig. 2006, 116, 2843–2854.
- Drachman, D.B.; Adams, R.N.; Josifek, L.F.; Pestronk, A.; Stanley, E.F. Antibody-mediated mechanisms of ACh receptor loss in myasthenia gravis: Clinical relevance. Ann. N. Y. Acad. Sci. 1981, 377, 175–188.
- Kao, I.; Drachman, D.B. Myasthenic immunoglobulin accelerates acetylcholine receptor degradation. Science 1977, 196, 527–529.
- Gomez, C.M.; Drachman, D.B. Anti-acetylcholine receptor antibodies directed against the α-bunbgarotoxin binding site induce a unique form of experimental myasthenia. Proc. Natl. Acad. Sci. USA 1983, 80, 4089–4093.
- Whiting, P.J.; Vincent, A.; Newsom-Davis, J. Acetylcholine receptor antibody characteristics in myasthenia gravis. Fractionation of alpha-bungarotoxin binding site antibodies and their relationship to IgG subclass. J. Neuroimmunol. 1983, 5, 1–9.
- Unwin, N. Refined structure of the nicotinic acetylcholine receptor at 4A resolution. J. Mol. Biol. 2005, 346, 967–989.
- Tuzun, E.; Christadoss, P. Complement associated pathogenic mechanisms in myasthenia gravis. Autoimmun. Rev. 2013, 12, 904–911.
- Sahashi, K.; Engel, A.G.; Lambert, E.H.; Howard, F.M., Jr. Ultrastructural localization of the terminal and lytic ninth complement component (C9) at the motor end-plate in myasthenia gravis. J. Neuropathol. Exp. Neurol. 1980, 39, 160–172.
- Tsujihata, M.; Yoshimura, T.; Satoh, A.; Kinoshita, I.; Matsuo, H.; Mori, M.; Nagataki, S. Diagnostic significance of IgG, C3, and C9 at the limb muscle motor end-plate in minimal myasthenia gravis. Neurology 1989, 39, 1359–1363.
- Lennon, V.A.; Seybold, M.E.; Lindstrom, J.M.; Cochrane, C.; Ulevitch, R. Role of complement in the pathogenesis of experimental autoimmune myasthenia gravis. J. Exp. Med. 1978, 147, 973–983.
- Donaldson, J.O.; Penn, A.S.; Lisak, R.P.; Abramski, O.; Brenner, T.; Schotland, D.L. Antiacetylcholine receptor antibody in neonatal myasthenia gravis. Am. J. Dis. Child. 1981, 135, 222–226.
- Namba, T.; Brown, S.B.; Grob, D. Neonatal myasthenia gravis: Report of two cases and review of the literature. Pediatrics 1970, 45, 488–504.
- Lefvert, A.K.; Osterman, P.O. Newborn infants to myasthenic mothers: A clinical study and an investigation of acetylcholine receptor antibodies in 17 children. Neurology 1983, 33, 133–138.
- Tzartos, S.J.; Efthimiadis, A.; Morel, E.; Eymard, B.; Bach, J.F. Neonatal myasthenia gravis: Antigenic specificities of antibodies in sera from mothers and their infants. Clin. Exp. Immunol. 1990, 80, 376–380.
- Gilhus, N.E. Myasthenia gravis can have consequences for pregnancy and the developing child. Front. Neurol. 2020, 11, 554.
- Samuels, P.; Bussel, J.B.; Braitman, L.E.; Tomaski, A.; Druzin, M.L.; Mennuti, M.T.; Cines, D.B. Estimation of the risk of Thrombocytonia in the offspring of pregnant women with presumed immune thrombocytopenic purpura. N. Engl. J. Med. 1990, 323, 229–235.
- Fujimura, K.; Harada, Y.; Fujimoto, T.; Kuramotop, A.; Ikeda, Y.; Akatsuka, J.; Dan, K.; Omine, M.; Mizoguchi, H. Nationwide study of idiopathic thrombocytopenic purpura in pregnant women and the clinical influence on neonates. Int. J. Hematol. 2002, 75, 426–433.
- Vincent, A.; Newland, C.; Brueton, L.; Beeson, D.; Riemersma, S.; Huson, S.M.; Newsom-Davis, J. Arthrogryposis multiplex congenita with maternal autoantibodies specific for a fetal antigen. Lancet 1995, 346, 24–25.
- Riemersma, S.; Vincent, A.; Beeson, D.; Newland, C.; Hawke, S.; Garabedian, B.V.-D.; Eymard, B.; Newsom-Davis, J. Association of arthrogryposis multiplex congenita with maternal antibodies inhibiting fetal acetylcholine receptor function. J. Clin. Investig. 1996, 98, 2358–2363.
- Saxena, A.; Stevens, J.; Cetin, H.; Koneczny, I.; Webster, R.; Lazaridis, K.; Tzartos, S.; Vrolix, K.; Nogales-Gadea, G.; Machiels, B.; et al. Characterization of an anti-fetal AChR monoclonal antibopdy isolated from a myasthenia gravis patient. Sci. Rep. 2017, 7, 14426.
- Hoch, W.; McConville, J.; Helms, S.; Newsom-Davis, J.; Melms, A.; Vincent, A. Auto-antibodies to the receptor tyrosine kinase MuSK in patients with myasthenia gravis without acetylcholine receptor antibodies. Nature Med. 2001, 7, 365–368.
- McConville, J.; Farrugia, M.E.; Beeson, D.; Kishore, U.; Mercalfe, R.; Newsom-Davis, J.; Vincent, A. Detection and characterization of MuSK antibodies in seronegative myasthenia gravis. Ann. Neurol. 2014, 55, 580–584.
- Sanders, D.B.; Ei-Salem, K.; Massey, J.M.; McConville, J.; Vincent, A. Clinical aspects of MuSK antibody positive seronegative MG. Neurology 2003, 60, 1978–1980.
- Zagar, M.; Vranjes, D.; Sostarko, M.; Vorgrinc, Z.; Bilic, E.; Cepe, M.T. Myasthenia gravis patients with anti-MuSK antibodies. Coll. Antropol. 2009, 33, 1151–1154.
- Chan, K.H.; Lachance, D.H.; Harper, C.M.; Lennon, V.A. Frequency of seronegativity in adult-acquired generalized myasthenia gravis. Muscle Nerve 2007, 36, 651–658.
- Chang, T.; Gunaratne, P.; Gamage, R.; Riffsy, M.T.M.; Vincent, A. MuSK-antibody-positive myasthenia gravis in a south Asian population. J. Neurol. Sci. 2009, 284, 33–35.
- Zhang, Z.; Guan, Y.; Han, J.; Li, M.; Shi, M.; Deng, H. Regional features of MuSK antibody-positive myasthenia gravis in North China. Front. Neurol. 2020, 11, 516211.
- Lee, J.-Y.; Sung, J.J.; Cho, J.-Y.; Oh, D.-H.; Kim, H.-J.; Park, J.-H.; Lee, K.W.; Choi, Y.-C.; Vincent, A. MuSK antibody-positive, seronegative myasthenia gravis in Korea. J. Clin. Neurosci. 2006, 13, 353–355.
- Ohta, K.; Shigemoto, K.; Fujinami, A.; Maruyama, N.; Konishi, T.; Ohta, M. Clinical and experimental features of MuSK antibody positive MG in Japan. Eur. J. Neurol. 2007, 14, 1029–1034.
- Murai, H.; Noda, T.; Himeno, E.; Kawano, Y.; Ohyagi, Y.; Shiraishi, H.; Motomura, M.; Kira, J. Infantile onset myasthenia gravis with MuSDK antibodies. Neurology 2006, 67, 174.
- Takahashi, Y.; Sugiyama, M.; Ueda, Y.; Itoh, T.; Yagyu, K.; Shiraishi, H.; Ukeba-Terashita, Y.; Nakanishi, M.; Nagashima, T.; Imai, T.; et al. Childhood-onset anti-MuSK antibody positive myasthenia gravis demonstrates a distinct clinical course. Brain Dev. 2012, 34, 784–786.
- Niks, E.H.; Kuks, J.B.M.; Roep, B.O.; Haasnoot, G.W.; Verduijn, W.; Ballieux, B.E.P.B.; De Baets, M.H.; Vincent, A.; Verschuuren, J.J.G.M. Strong association of MuSK antibody-positive myasthenia gravis and HLA-DR14-DQ5. Neurology 2006, 66, 1772–1774.
- Kanai, T.; Uzawa, A.; Kawaguchi, N.; Sakamaki, T.; Yoshiyama, Y.; Himuro, K.; Oda, F.; Kuwabara, S. HLA-DRB1*14 and DQB1*05 are associated with Japanese anti-MuSK antibody-positive myasthenia gravis patients. J. Neurol. Sci. 2016, 363, 116–118.
- Hong, Y.; Li, H.-F.; Romi, F.; Skeie, G.O.; Gilhus, N.E. HLA and MuSK-positive myasthenia gravis: A systemtic review and meta-analysis. Acta Neurol. Scand. 2018, 138, 219–226.
- Inoue, K.; Tsugawa, J.; Fukae, J.; Fukuhara, K.; Kawano, H.; Fujioka, S.; Tsuboi, Y. Myasthenia gravis with anti-muscle-specific tyrosine kinase antibody during pregnancy and risk of neonatal myasthenia gravis: A case report and review of the literature. Case Rep. Neurol. 2020, 12, 114–120.
- Bennett, D.L.H.; Mills, K.R.; Riordan-Eva, P.; Barnes, P.R.J.; Rose, M.R. Anti-MuSK antibodies in a case of ocular myasthenia gravis. J. Neurol. Neurosurg. Psychiatry 2005, 564, 075812.
- Hosaka, A.; Takuma, H.; Ohta, K.; Tamaoka, A. An ocular form of myasthenia gravis with a high titer of anti-MuSK antibodies during a long-term follow-up. Int. Med. 2012, 51, 3077–3079.
- Shiraishi, H.; Motomura, M.; Yoshimura, T.; Fukudome, T.; Fukuda, T.; Nakao, Y.; Tsujihata, M.; Vincent, A.; Eguchi, K. Acetylcholine receptors loss and postsynaptic damage in MuSK antibody positive myasthenia gravis. Ann. Neurol. 2005, 57, 289–293.
- Koneczny, I.; Cossins, J.; Waters, P.; Beeson, D.; Vincent, A. MuSK myasthenia gravis IgG4 disrupts the interaction of LRP4 with MuSK but both IgG4 and IgG1-3 can disperse preformed agrin-independent AChR clusters. PLoS ONE 2013, 8, e80695.
- Meriggioli, M.N.; Sanders, D.B. Autoimmune myasthenia gravis: Emerging clinical and biological heterogeneity. Lancet Neurol. 2009, 8, 475–490.
- Otsuka, K.; Ito, M.; Ohkawara, B.; Masuda, A.; Kawakami, Y.; Sahashi, K.; Nishida, H.; Mabuchi, N.; Takano, A.; Engel, A.G.; et al. Collagen Q and anti-MuSK autoantibody competitively suppress agrin/LRP4/MuSK signaling. Sci. Rep. 2015, 5, 13928.
- Rodriguez Cruz, P.M.; Al-Hjjar, M.; Huda, S.; Jacobson, L.; Woodhall, M.; Jayawant, S.; Buckley, C.; Hilton-Jones, D.; Beeson, D.; Vincent, A.; et al. Clinical features and diagnostic usefulness of antibodies to cultured acetylcholine receptors in the diagnosis of seronegative myasthenia gravis. JAMA Neurol. 2015, 72, 642–649.
- Budhram, A. Fixed cell-based assays for autoantibody detection in myasthenia gravis: A diagnostic breakthrough. Lancet Reg. Health West. Pac. 2023, 38, 100876.
- Pevzner, A.; Schoser, B.; Peters, K.; Cosma, N.-C.; Karakatsani, A.; Schalke, B.; Melms, A.; Kröger, S. Anti-LRP4 autoantibodies in AChR- and MuSK-antibody-negative myasthenia gravis. J. Neurol. 2012, 259, 427–435.
- Higuchi, O.; Hamuro, J.; Motomura, M.; Yamanashi, Y. Autoantibodies to low-density lipoprotein receptor-related protein in myasthenia gravis. Ann. Neurol. 2011, 69, 418–422.
- Tzartos, J.S.; Zhang, B.; Belimezi, M.; Ragheb, S.; Bealmear, B.; Lewis, R.A.; Xiong, W.-C.; Lisak, R.P.; Tzartos, S.J.; Mei, L. Autoantibodies to lipoprotein-related protein 4 patients with double-seronegative myasthenia gravis. Arch. Neurol. 2012, 69, 445–451.
- Shen, C.; Lu, Y.; Zhang, B.; Figueiredo, D.; Bean, J.; Jung, J.; Wu, H.; Barik, A.; Yin, D.-M.; Xiong, W.-C.; et al. Antibodies against low-density lipoprotein receptor-related protein 4 induce myasthenia gravis. J. Clin. Investig. 2013, 123, 5190–5202.
- Gasperi, C.; Melms, A.; Schoser, B.; Zhang, Y.; Meltoranta, J.; Risson, V.; Schaeffer, L.; Schalke, B.; Kröger, S. Anti-agrin autoantibodies in myasthenia gravis. Neurology 2014, 82, 1976–1983.
- Zhang, B.; Shen, C.; Bealmear, B.; Ragheb, S.; Xiong, W.-C.; Lewis, R.A.; Lisak, R.P.; Mei, L. Autoantibodies to agrin in myasthenia gravis patients. PLoS ONE 2014, 9, e91816.
- Yan, M.; Liu, Z.; Fei, E.; Chen, W.; Lai, X.; Luo, B.; Chen, P.; Jing, H.; Pan, J.-X.; Rivner, M.H.; et al. Induction of anti-agrin antibodies causes myasthenia gravis in mice. Neuroscience 2018, 373, 113–121.
- Rivner, M.H.; Quarles, B.M.; Pan, J.-X.; Yu, Z.; Howard, J.F.; Corse, A.; Dimachkie, M.M.; Jackson, C.; Vu, T.; Small, G.; et al. Clinical features of LRP4/agrin-antibody-positive myasthenia gravis: A multicenter study. Muscle Nerve 2020, 62, 333–343.
- Kaminski, H.J. Seronegative myasthenia gravis—A vanishing disorder? JAMA Neurol. 2016, 73, 1055–1056.
- Drachman, D.B. How to recognize an antibody-mediated autoimmune disease: Criteria. Res. Publ. Assoc. Res. Nerv. Ment. Dis. 1990, 68, 183–186.
- Shigemoto, K.; Kubo, S.; Maruyama, N.; Hato, N.; Yamada, H.; Jie, C.; Kobayashi, N.; Mominoki, K.; Abe, Y.; Ueda, N.; et al. Induction of myasthenia gravis by immunization against muscle-specific kinase. J. Clin. Investig. 2006, 116, 1016–1024.
- Viegas, S.; Jacobson, L.; Waters, P.; Cossins, J.; Jacob, S.; Leite, M.I.; Webster, R.; Vincent, A. Passive and active immunization models of MuSK-Ab positive myasthenia: Electrophysiological evidence for pre and postsynaptic defects. Exp. Neurol. 2012, 234, 506–512.
- Ulsoy, C.; Cavus, F.; Yilmaz, V.; Tuzun, E. Immunization with recombinantly expressed LRP4 induces experimental autoimmune myasthenia gravis in C57BL/6 mice. Immunol. Investig. 2017, 46, 490–499.
- Mori, S.; Motohashi, N.; Takashima, R.; Kishi, M.; Nishimune, H.; Shigemoto, K. Immunization of mice with LRP4 induces myasthenia similar to MuSK-associated myasthenia gravis. Exp. Neurol. 2017, 297, 158–167.
- Yu, Z.; Zhang, M.; Jing, H.; Chen, P.; Cao, R.; Pan, J.; Luo, B.; Yu, Y.; Quarles, B.M.; Xiong, W.; et al. Characterization of LRP4/Agrin antibodies from a patient with myasthenia gravis. Neurology 2021, 97, e975–e987.
- Engel, A.G.; Shen, X.-M.; Selcen, D.; Sine, S.M. Congenital myasthenic syndrome: Pathogenesis, diagnosis, and treatment. Lancet Neurol. 2015, 14, 420–434.
- Parr, J.R.; Andrew, M.J.; Finnis, M.; Beeson, D.; Vincent, A.; Jayawant, S. How common is childhood myasthenia? The UK incidence and prevalence of autoimmune and congenital myasthenia. Arch. Dis. Child. 2014, 99, 539–542.
- Mansukhani, S.A.; Bothun, E.; Diehl, N.N.; Mohney, B.G. Incidence and ocular features of pediatric myasthenias. Am. J. Ophthalmol. 2019, 200, 242–249.
- Azuma, Y.; Nakata, T.; Tanaka, M.; Shen, X.-M.; Ito, M.; Iwata, S.; Okuno, T.; Nomura, Y.; Ando, N.; Ishigaki, K.; et al. Congenital myasthenic syndrome in Japan: Ethnically unique mutations in muscle nicotinic acetylcholine receptor subunits. Neuromuscul. Disord. 2015, 25, 60–69.
- Ohno, K.; Ohkawara, B.; Shen, X.-M.; Selcen, D.; Engel, A.G. Clinical and pathologic features of congenital myasthenic syndromes caused by 35 genes—A comprehensive review. Int. J. Mol. Sci. 2023, 24, 3730.
- Hayashi, M.; Kida, K.; Yamada, I.; Morimoto, T.; Matsuda, H.; Mimaki, T.; Yabuuchi, H. Anti-acetylcholine receptor antibody in juvenile and adult myasthenia gravis. Acta Paediatr. Jpn. 1986, 28, 781–787.
- Murai, H.; Yamashita, N.; Watanabe, M.; Nomura, Y.; Motomura, M.; Yoshikawa, H.; Nakamura, Y.; Kawaguchi, N.; Onodera, H.; Araga, S.; et al. Characteristics of myasthenia gravis according to onset-age: Japanese Nationwide survey. J. Neurol. Sci. 2011, 305, 97–102.
- Zao, G.; Wang, X.; Yu, X.; Zhang, X.; Guan, Y.; Jiang, J. Clinical application of clustered-AChR for the detection SNMG. Sci. Rep. 2015, 5, 10193.
- Oda, K. Differences in acetylcholine receptor-antibody interactions between extraocular and extremity muscle fibers. Ann. N. Y. Acad. Sci. 1993, 681, 238–255.
- Kaminski, H.J.; Maas, E.; Spiegel, P.; Ruff, R.L. Why are eye muscles frequently involved in myasthenia gravis? Neurology 1990, 40, 1663–1669.
- Horton, R.M.; Manfredi, A.A.; Conti-Tronconi, B.M. The ‘embrionic’ gamma subunit of the nicotinic acetylcholine receptor is expressed in adult extraocular muscle. Neurology 1993, 43, 983–986.
- Missias, A.C.; Chu, G.C.; Klocke, B.J.; Sanes, J.R.; Merlie, J.P. Maturation of the acetylcholine receptor in skeletal muscle: Regulation of the AChR γ-to-ϵ switch. Dev. Biol. 1996, 179, 223–238.
- Oda, K. Motor innervation and acetylcholine receptor distribution of human extraocular muscle fibers. J. Neurol. Sci. 1986, 74, 125–133.
- Kaminski, H.J.; Kusner, L.L.; Block, C.H. Expression of acetylcholine receptor isoforms at extraocular muscle endplates. Investig. Ophthalmol. Vis. Sci. 1996, 37, 345–351.
- Ruff, R.L.; Lennon, V.A. How myasthenia gravis alters the safety factor for neuromuscular transmission? J. Neuroimmunol. 2008, 201–202, 13–20.
- Serra, A.; Ruff, R.; Leigh, R.J. Neuromuscular transmission failure in myasthenia gravis: Decrement of safety factor and susceptibility of extraocular muscle. Ann. N. Y. Acad. Sci. 2012, 1275, 129–135.
- Yoshikawa, H.; Adachi, Y.; Nakamura, Y.; Kuriyama, N.; Murai, H.; Nomura, Y.; Sakai, Y.; Iwasa, K.; Furukawa, Y.; Kuwabara, S.; et al. Two-step nationwide epidemiological survey of myasthenia gravis in Japan 2018. PLoS ONE 2018, 17, e0274161.
- Park, J.-S.; Eah, K.Y.; Park, J.-M. Epidemiological profile of myasthenia gravis in South Korea using the national health insurance database. Acta Neurol. Scand. 2022, 145, 633–640.
- Huang, X.; Liu, W.B.; Men, L.N.; Feng, H.Y.; Li, Y.; Luo, C.M.; Qiu, L. Clinical features of myasthenia gravis in southern China: A retrospective review of 2154 cases over 22 years. Neurol. Sci. 2013, 34, 911–917.
- Popperud, T.H.; Boldingh, M.I.; Rasmussen, M.; Kerty, E. Juvenile myasthenia gravis in Norway: Clinical characteristics, treatment, and long-term outcome in a nationwide population-based cohort. Eur. J. Pediatr. Neurol. 2017, 21, 707–714.
- Heckmann, J.M.; Europa, T.A.; Soni, A.J.; Nel, M. The epidemiology and phenotypes of ocular manifestations in childhood and juvenile myasthenia gravis: A review. Front. Neurol. 2022, 13, 834212.
- Thapa, P.; Farber, D.L. The role of the thymus in the immune response. Thorac. Surg. Clin. 2019, 29, 123–1312.
- Kondo, K.; Ohigashi, I.; Takahama, Y. Thymus machinery for T-cell selection. Int. Immunol. 2019, 31, 119–125.
- Thome, J.J.C.; Grinshpun, B.; Kumar, B.V.; Kubota, M.; Ohmura, Y.; Lerner, H.; Sempowski, G.D.; Shen, Y.; Farber, D.L. Long-term maintenance of human naïve T cell through in situ homeostasis in lymphoid tissue sites. Sci. Immunol. 2016, 1, eaah6506.
- Crooke, S.N.; Ovsyannikova, I.G.; Poland, G.A.; Kennedy, R.B. Immunosenescence and human vaccine immune responses. Immun. Ageing 2019, 16, 25.
- Berman, P.W.; Patrick, J. Experimental myasthenia gravis: A murine system. J. Exp. Med. 1980, 151, 204–223.
- Christadoss, P.; Lennon, V.A.; Krco, C.J.; David, C.S. Genetic control of experimental autoimmune myasthenia gravis in mice III. Ia molecules mediate cellular immune responsiveness to acetylcholine receptors. J. Immunol. 1982, 128, 1141–1144.
- McIntyre, K.R.; Seidman, J.G. Nucleotide sequence of mutant I-Abbm12 gene is evidence for genetic exchange between mouse immune response genes. Nature 1984, 308, 551–553.
- Christadoss, P.; Lindstrom, J.M.; Melvold, R.W.; Talal, N. Mutation at I-A beta chain prevents experimental autoimmune myasthenia gravis. Immunogenet 1985, 21, 33–38.
- Bellone, M.; Ostlie, N.; Lei, S.; Wu, X.-D.; Conti-Tronconi, B.M. The I-Abm12 mutation, which confers resistance to experimental myasthenia gravis, drastically affects the epitope repertoire of murine CD4+ cells sensitized to nicotinic acetylcholine receptor. J. Immunol. 1991, 147, 1484–1491.
- Infante, A.J.; Thompson, P.A.; Krolick, K.A.; Wall, K.A. Determinant selection in murine experimental autoimmune myasthenia gravis: Effect of the bm12 mutation on T cell recognition of acetylcholine receptor epitopes. J. Immunol. 1991, 146, 2977–2982.
- Wu, X.; Tuzun, E.; Saini, S.S.; Wang, J.; Li, J.; Aguilera-Aguirre, L.; Huda, R.; Christadoss, P. Ocular myasthenia gravis induced by human acetylcholine receptor e subunit immunization in HLA-DR3 transgenic mice. Immunol. Lett. 2015, 168, 306–3012.
- Koreth, J.; Matsuoka, K.; Kim, H.T.; McDonough, S.M.; Bindra, B.; Alyea, E.P., III; Armand, P.; Cutler, C.; Ho, V.T.; Treister, N.S.; et al. Interleukin-2 and regulatory T cells in graft-versus-host disease. N. Engl. J. Med. 2011, 365, 2055–2066.
- Namba, T.; Shapiro, M.S.; Brunner, N.G.; Grob, D. Myasthenia gravis occurring in twins. J. Neurol. Neurosurg. Psychiatr. 1971, 34, 531–534.
- O’Connell, K.; Ramdas, S.; Palace, J. Management of juvenile myasthenia gravis. Front. Neurol. 2020, 11, 743.
- Hayashi, M. Childhood myasthenia gravis in Japan: Pathophysiology and treatment options. Clin. Exp. Neuroimmunol. 2023, 14, 185–194.
- Gilhus, N.E.; Skeie, G.O.; Romi, F.; Lazaridis, K.; Zisimopoulou, P.; Tzartos, S. Myasthenia gravis—Autoantibody characteristics and their implications for therapy. Nat. Rev. Neurol. 2016, 12, 259–268.
- Hayashi, M.; Kida, K.; Yoshinaga, J. Possible distinct pathogenesis in low responder myasthenia gravis: Association of soluble interleukin-2 receptor with acetylcholine receptor antibody titre or abnormal thymus. J. Neurol. Neurosurg. Psychiatr. 1996, 61, 207–208.
- Wiesendanger, M.; D’Alessandri, A. Myasthenia gravis mit fokaler infiltration der endplattenzone. Acta Neuropathol. 1963, 2, 246–252.
- Fenichel, G.M.; Shy, G.M. Muscle biopsy experience in myasthenia gravis. Arch. Neurol. 1963, 9, 237–243.
- Fenichel, G.M. Muscle lesions in myasthenia gravis. Ann. N. Y. Acad. Sci. 1966, 135, 60–67.
- Pascuzzi, R.M.; Campa, J.F. Lymphorrhage localized to the muscle end-plate in myasthenia gravis. Arch. Pathol. Lab. Med. 1988, 112, 934–937.
- Maselli, R.A.; Richman, D.P.; Wollmann, R.L. Inflammation at the neuromuscular junction in myasthenia gravis. Neurology 1991, 41, 1497–1504.
- Nakano, S.; Engel, A.G. Myasthenia gravis: Quantitative immunocytochemical analysis of inflammatory cells and detection of complement membrane attack complex at the endplate in 30 patients. Neurology 1993, 43, 1167–1172.
- Hans, J.G.H. Oosterhuis. Animal model of myasthenia gravis. In Myasthenia Gravis; Glaser, G.H., Barbeau, A., Eds.; Clinical Neurology Neurosurgery Monographs; Churchill Livingstone: Edinburgh, UK; Melbourne, Australia; New York, NY, USA, 1984; Volume 5, pp. 131–141.
- Lindstrom, J.M.; Engel, A.G.; Seybold, M.E.; Lennon, V.A.; Lambert, E.H. Pathological mechanisms in experimental autoimmune myasthenia gravis. II. Passive transfer of experimental autoimmune myasthenia gravis in rats with anti-acetylcholine receptor antibodies. J. Exp. Med. 1976, 144, 739–753.
- Gomez, C.M.; Wollmann, R.L.; Richman, D.P. Induction of morphologic changes of both acute and chronic experimental myasthenia by monoclonal antibody directed against acetylcholine receptor. Acta Neuropathol. 1984, 63, 131–143.
- Zhou, Y.; Kaminski, H.J.; Gong, B.; Cheng, G.; Feuerman, J.M.; Kusner, L. RNA expression analysis of passive transfer myasthenia supports extraocular muscle as a unique immunological environment. Investig. Ophthalmol. Vis. Sci. 2014, 55, 4348–4359.
More
Information
Subjects:
Pathology; Neurosciences; Medicine, Research & Experimental
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
657
Revisions:
2 times
(View History)
Update Date:
24 Jan 2024
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No