 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Biji Thomas George | -- | 2856 | 2024-01-02 08:35:41 | | | |
| 2 | Wendy Huang | Meta information modification | 2856 | 2024-01-05 06:45:17 | | |
Video Upload Options
Overweight and obesity are terms used to describe an abnormal and excessive build-up of body fat that may have a detrimental effect on health. Body mass index (BMI) is a measure of body weight per height that is widely used to classify adults as overweight or obese. It is calculated by dividing a person’s body weight (in kilograms) by the square of their height (in meters/m2). The World Health Organization (WHO) defines overweight as a BMI of 25 kg/m2 or above and obesity as a BMI of 30 kg/m2 or above. The rising incidence of obesity has coincided with rising levels of poor reproductive outcomes. The molecular basis for the association of infertility in obese males is now being explained through various mechanisms. Insulin resistance, hyperglycemia, and changes in serum and gonadal concentrations of adipokines, like leptin, adiponectin, resistin, and ghrelin have been implicated as causes of male infertility in obese males.
1. Introduction
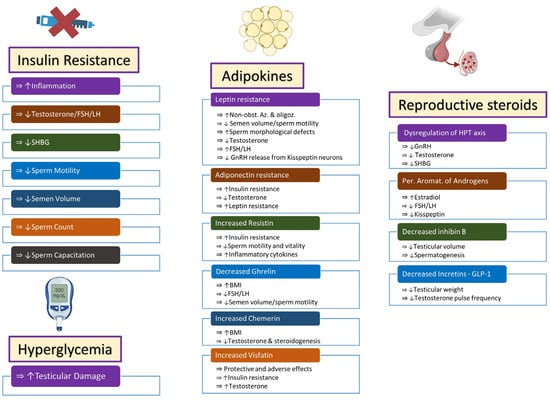
2. Insulin Resistance and Hyperglycemia
2.1. Insulin Resistance
2.2. Hyperglycemia
3. Adipokines
3.1. Leptin
3.2. Adiponectin
3.3. Resistin
3.4. Ghrelin
3.5. Chemerin
3.6. Visfatin
3.7. Apelin, Omentin, Hepcidin, and Vaspin
4. Reproductive Steroids
4.1. Hypothalamic–Pituitary–Testicular Axis
4.2. Estrogens
4.3. Inhibin B
5. Incretins
References
- Meldrum, D.R.; Morris, M.A.; Gambone, J.C. Obesity pandemic: Causes, consequences, and solutions-but do we have the will? Fertil. Steril. 2017, 107, 833–839.
- Obesity and Overweight. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 18 October 2023).
- Barbagallo, F.; Condorelli, R.A.; Mongioì, L.M.; Cannarella, R.; Cimino, L.; Magagnini, M.C.; Crafa, A.; La Vignera, S.; Calogero, A.E. Molecular Mechanisms Underlying the Relationship between Obesity and Male Infertility. Metabolites 2021, 11, 840.
- Chambers, T.J.; Richard, R.A. The impact of obesity on male fertility. Hormones 2015, 14, 563–568.
- Kahn, B.E.; Brannigan, R.E. Obesity and male infertility. Curr. Opin. Urol. 2017, 27, 441–445.
- Swan, S.H.; Elkin, E.P.; Fenster, L. The question of declining sperm density revisited: An analysis of 101 studies published 1934–1996. Environ. Health Perspect. 2000, 108, 961–966.
- Sermondade, N.; Faure, C.; Fezeu, L.; Lévy, R.; Czernichow, S. Obesity and increased risk for oligozoospermia and azoospermia. Arch. Intern. Med. 2012, 172, 440–442.
- Ellulu, M.S.; Patimah, I.; Khaza’ai, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863.
- Leisegang, K.; Sengupta, P.; Agarwal, A.; Henkel, R. Obesity and male infertility: Mechanisms and management. Andrologia 2021, 53, e13617.
- Craig, J.R.; Jenkins, T.G.; Carrell, D.T.; Hotaling, J.M. Obesity, male infertility, and the sperm epigenome. Fertil. Steril. 2017, 107, 848–859.
- Alves, M.G.; Jesus, T.T.; Sousa, M.; Goldberg, E.; Silva, B.M.; Oliveira, P.F. Male fertility and obesity: Are ghrelin, leptin and glucagon-like peptide-1 pharmacologically relevant? Curr. Pharm. Des. 2016, 22, 783–791.
- Dupont, J.; Pollet-Villard, X.; Reverchon, M.; Mellouk, N.; Levy, R. Adipokines in human reproduction. Horm. Mol. Biol. Clin. Investig. 2015, 24, 11–24.
- Kawwass, J.F.; Summer, R.; Kallen, C.B. Direct effects of leptin and adiponectin on peripheral reproductive tissues: A critical review. Mol. Hum. Reprod. 2015, 21, 617–632.
- Fushimi, H.; Horie, H.; Inoue, T.; Kameyama, M.; Kanao, K.; Ishihara, S.; Tsujimura, T.; Nunotani, H.; Minami, T.; Okazaki, Y.; et al. Low testosterone levels in diabetic men and animals: A possible role in testicular impotence. Diabetes Res. Clin. Pract. 1989, 6, 297–301.
- Carrageta, D.F.; Oliveira, P.F.; Alves, M.G.; Monteiro, M.P. Obesity and male hypogonadism: Tales of a vicious cycle. Obes. Rev. 2019, 20, 1148–1158.
- Barbagallo, F.; La Vignera, S.; Cannarella, R.; Mongioì, L.M.; Garofalo, V.; Leanza, C.; Marino, M.; Calogero, A.E.; Condorelli, R.A. Obesity and Male Reproduction: Do Sirtuins Play a Role? Int. J. Mol. Sci. 2022, 23, 973.
- Martins, A.D.; Majzoub, A.; Agawal, A. Metabolic Syndrome and Male Fertility. World J. Mens. Health 2019, 37, 113–127.
- Zańko, A.; Siewko, K.; Krętowski, A.J.; Milewski, R. Lifestyle, Insulin Resistance and Semen Quality as Co-Dependent Factors of Male Infertility. Int. J. Environ. Res. Public Health 2022, 20, 732.
- Ma, J.; Han, R.Y.; Mei, X.A.; Qi, Y.N.; Ma, J.Y.; Liu, W.J.; Wang, S.S. Correlation of insulin resistance with male reproductive hormone levels and semen parameters. Zhonghua Nan Ke Xue 2018, 24, 695–699.
- Mansour, R.; El-Faissal, Y.; Kamel, A.; Kamal, O.; Aboulserour, G.; Aboulghar, M.; Fahmy, I. Increased insulin resistance in men with unexplained infertility. Reprod. Biomed. Online 2017, 35, 571–575.
- Morrison, C.D.; Brannigan, R.E. Metabolic syndrome and infertility in men. Best Pract. Res. Clin. Obstet. Gynaecol. 2015, 29, 507–515.
- AbbasiHormozi, S.; Kouhkan, A.; Shahverdi, A.; Parikar, A.; Shirin, A.; Vesali, S. How much obesity and diabetes do impair male fertility? Reprod. Biol. Endocrinol. 2023, 21, 48.
- Hammoud, A.O.; Gibson, M.; Peterson, C.M.; Meikle, A.W.; Carrell, D.T. Impact of male obesity on infertility: A critical review of the current literature. Fertil. Steril. 2008, 90, 897–904.
- Jin, S.K.; Yang, W.X. Factors and pathways involved in capacitation: How are they regulated? Oncotarget 2017, 8, 3600–3627.
- Okabe, M. Sperm-egg interaction and fertilization: Past, present, and future. Biol. Reprod. 2018, 99, 134–146.
- Aquila, S.; Gentile, M.; Middea, E.; Catalano, S.; Andò, S. Autocrine regulation of insulin secretion in human ejaculated spermatozoa. Endocrinology 2005, 146, 552–557.
- Aitken, R.J.; Curry, B.J.; Shokri, S.; Pujianto, D.A.; Gavriliouk, D.; Gibb, Z.; Whiting, S.; Connaughton, H.S.; Nixon, B.; Salamonsen, L.A.; et al. Evidence that extrapancreatic insulin production is involved in the mediation of sperm survival. Mol. Cell. Endocrinol. 2021, 526, 111193.
- Pasquali, R.; Patton, L.; Gambineri, A. Obesity and infertility. Curr. Opin. Endocrinol. Diabetes Obes. 2007, 14, 482–487.
- Lampiao, F.; Agarwal, A.; du Plessis, S.S. Invited review The role of insulin and leptin in male reproduction. Arch. Med. Sci. Spec. Issues 2009, 5, S48–S54.
- Maresch, C.C.; Stute, D.C.; Alves, M.G.; Oliveira, P.F.; de Kretser, D.M.; Linn, T. Diabetes-induced hyperglycemia impairs male reproductive function: A systematic review. Hum. Reprod. Update 2018, 24, 86–105.
- Facondo, P.; Di Lodovico, E.; Delbarba, A.; Anelli, V.; Pezzaioli, L.C.; Filippini, E.; Cappelli, C.; Corona, G.; Ferlin, A. The impact of diabetes mellitus type 1 on male fertility: Systematic review and meta-analysis. Andrology 2022, 10, 426–440.
- Ding, G.L.; Liu, Y.; Liu, M.E.; Pan, J.X.; Guo, M.X.; Sheng, J.Z.; Huang, H.F. The effects of diabetes on male fertility and epigenetic regulation during spermatogenesis. Asian J. Androl. 2015, 17, 948–953.
- Maresch, C.C.; Stute, D.C.; Fleming, T.; Lin, J.; Hammes, H.P.; Linn, T. Hyperglycemia induces spermatogenic disruption via major pathways of diabetes pathogenesis. Sci. Rep. 2019, 9, 13074.
- Omolaoye, T.S.; du Plessis, S.S. Male infertility: A proximate look at the advanced glycation end products. Reprod. Toxicol. 2020, 93, 169–177.
- Roumaud, P.; Martin, L.J. Roles of leptin, adiponectin and resistin in the transcriptional regulation of steroidogenic genes contributing to decreased Leydig cells function in obesity. Horm. Mol. Biol. Clin. Investig. 2015, 24, 25–45.
- Tsatsanis, C.; Dermitzaki, E.; Avgoustinaki, P.; Malliaraki, N.; Mytaras, V.; Margioris, A.N. The impact of adipose tissue-derived factors on the hypothalamic-pituitary-gonadal (HPG) axis. Hormones 2015, 14, 549–562.
- Childs, G.V.; Odle, A.K.; MacNicol, M.C.; MacNicol, A.M. The Importance of Leptin to Reproduction. Endocrinology 2021, 162, bqaa204.
- Khodamoradi, K.; Parmar, M.; Khosravizadeh, Z.; Kuchakulla, M.; Manoharan, M.; Arora, H. The role of leptin and obesity on male infertility. Curr. Opin. Urol. 2020, 30, 334–339.
- Ghaderpour, S.; Ghiasi, R.; Heydari, H.; Keyhanmanesh, R. The relation between obesity, kisspeptin, leptin, and male fertility. Horm. Mol. Biol. Clin. Investig. 2021, 43, 235–247.
- Ahmad, R.; Haque, M. Obesity: A Doorway to a Molecular Path Leading to Infertility. Cureus 2022, 14, e30770.
- Isidori, A.M.; Caprio, M.; Strollo, F.; Moretti, C.; Frajese, G.; Isidori, A.; Fabbri, A. Leptin and androgens in male obesity: Evidence for leptin contribution to reduced androgen levels. J. Clin. Endocrinol. Metab. 1999, 84, 3673–3680.
- Thomas, S.; Kratzsch, D.; Schaab, M.; Scholz, M.; Grunewald, S.; Thiery, J.; Paasch, U.; Kratzsch, J. Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil. Steril. 2013, 99, 1256–1263.e3.
- Page, S.T.; Herbst, K.L.; Amory, J.K.; Coviello, A.D.; Anawalt, B.D.; Matsumoto, A.M.; Bremner, W.J. Testosterone administration suppresses adiponectin levels in men. J. Androl. 2005, 26, 85–92.
- Wilkinson, M.; Brown, R.; Imran, S.A.; Ur, E. Adipokine gene expression in brain and pituitary gland. Neuroendocrinology 2007, 86, 191–209.
- Heydari, H.; Ghiasi, R.; Ghaderpour, S.; Keyhanmanesh, R. The Mechanisms Involved in Obesity-Induced Male Infertility. Curr. Diabetes Rev. 2021, 17, 259–267.
- Wang, Y.; Cao, L.; Liu, X. Ghrelin alleviates endoplasmic reticulum stress and inflammation-mediated reproductive dysfunction induced by stress. J. Assist. Reprod. Genet. 2019, 36, 2357–2366.
- Kheradmand, A.; Dezfoulian, O.; Alirezaei, M.; Rasoulian, B. Ghrelin modulates testicular germ cells apoptosis and proliferation in adult normal rats. Biochem. Biophys. Res. Commun. 2012, 419, 299–304.
- Kluge, M.; Schussler, P.; Uhr, M.; Yassouridis, A.; Steiger, A. Ghrelin suppresses secretion of luteinizing hormone in humans. J. Clin. Endocrinol. Metab. 2007, 92, 3202–3205.
- Kluge, M.; Uhr, M.; Bleninger, P.; Yassouridis, A.; Steiger, A. Ghrelin suppresses secretion of FSH in males. Clin. Endocrinol. 2009, 70, 920–923.
- Wang, C.; Jackson, G.; Jones, T.H.; Matsumoto, A.M.; Nehra, A.; Perelman, M.A.; Swerdloff, R.S.; Traish, A.; Zitzmann, M.; Cunningham, G. Low testosterone associated with obesity and the metabolic syndrome contributes to sexual dysfunction and cardiovascular disease risk in men with type 2 diabetes. Diabetes Care 2011, 34, 1669–1675.
- Kısa, Ü.; Başar, M.M.; Şipal, T.; Ceylan, Ö.D. Ghrelin and orexin levels in infertile male: Evaluation of effects on varicocele pathophysiology, relationship seminal and hormonal parameter. Turk. J. Biochem. 2020, 45, 877–882.
- Li, L.; Huang, C.; Zhang, X.; Wang, J.; Ma, P.; Liu, Y.; Xiao, T.; Zabel, B.A.; Zhang, J.V. Chemerin-derived peptide C-20 suppressed gonadal steroidogenesis. Am. J. Reprod. Immunol. 2014, 71, 265–277.
- Li, L.; Ma, P.; Huang, C.; Liu, Y.; Zhang, Y.; Gao, C.; Xiao, T.; Ren, P.G.; Zabel, B.A.; Zhang, J.V. Expression of chemerin and its receptors in rat testes and its action on testosterone secretion. J. Endocrinol. 2014, 220, 155–163.
- Yu, M.; Yang, Y.; Huang, C.; Ge, L.; Xue, L.; Xiao, Z.; Xiao, T.; Zhao, H.; Ren, P.; Zhang, J.V. Chemerin: A Functional Adipokine in Reproductive Health and Diseases. Biomedicines 2022, 10, 1910.
- Singh, A.; Choubey, M.; Bora, P.; Krishna, A. Adiponectin and Chemerin: Contrary Adipokines in Regulating Reproduction and Metabolic Disorders. Reprod. Sci. 2018, 25, 1462–1473.
- Zorena, K.; Jachimowicz-Duda, O.; Slezak, D.; Robakowska, M.; Mrugacz, M. Adipokines and Obesity. Potential Link to Metabolic Disorders and Chronic Complications. Int. J. Mol. Sci. 2020, 21, 3570.
- Wang, W.D.; Xing, L.; Teng, J.R.; Li, S.; Mi, N.A. Effects of basal insulin application on serum visfatin and adiponectin levels in type 2 diabetes. Exp. Ther. Med. 2015, 9, 2219–2224.
- Chang, Y.C.; Chang, T.J.; Lee, W.J.; Chuang, L.M. The relationship of visfatin/pre-B-cell colony-enhancing factor/nicotinamide phosphoribosyltransferase in adipose tissue with inflammation, insulin resistance, and plasma lipids. Metabolism 2010, 59, 93–99.
- Hameed, W.; Yousaf, I.; Latif, R.; Aslam, M. Effect of visfatin on testicular steroidogenesis in purified Leydig cells. J. Ayub. Med. Coll. Abbottabad 2012, 24, 62–64.
- Elfassy, Y.; Bastard, J.P.; McAvoy, C.; Fellahi, S.; Dupont, J.; Levy, R. Adipokines in Semen: Physiopathology and Effects on Spermatozoas. Int. J. Endocrinol. 2018, 2018, 3906490.
- Wozniak, S.E.; Gee, L.L.; Wachtel, M.S.; Frezza, E.E. Adipose tissue: The new endocrine organ? A review article. Dig. Dis. Sci. 2009, 54, 1847–1856.
- Higuchi, K.; Masaki, T.; Gotoh, K.; Chiba, S.; Katsuragi, I.; Tanaka, K.; Kakuma, T.; Yoshimatsu, H. Apelin, an APJ receptor ligand, regulates body adiposity and favors the messenger ribonucleic acid expression of uncoupling proteins in mice. Endocrinology 2007, 148, 2690–2697.
- Moretti, E.; Signorini, C.; Noto, D.; Tripodi, S.A.; Menchiari, A.; Sorrentino, E.; Collodel, G. Seminal Levels of Omentin-1/ITLN1 in Inflammatory Conditions Related to Male Infertility and Localization in Spermatozoa and Tissues of Male Reproductive System. J. Inflamm. Res. 2022, 15, 2019–2031.
- de Souza Batista, C.M.; Yang, R.Z.; Lee, M.J.; Glynn, N.M.; Yu, D.Z.; Pray, J.; Ndubuizu, K.; Patil, S.; Schwartz, A.; Kligman, M.; et al. Omentin plasma levels and gene expression are decreased in obesity. Diabetes 2007, 56, 1655–1661.
- Bekri, S.; Gual, P.; Anty, R.; Luciani, N.; Dahman, M.; Ramesh, B.; Iannelli, A.; Staccini-Myx, A.; Casanova, D.; Ben Amor, I.; et al. Increased adipose tissue expression of hepcidin in severe obesity is independent from diabetes and NASH. Gastroenterology 2006, 131, 788–796.
- Sokol, R.Z. Endocrinology of male infertility: Evaluation and treatment. Semin. Reprod. Med. 2009, 27, 149–158.
- Anawalt, B.D. Approach to Male Infertility and Induction of Spermatogenesis. J. Clin. Endocrinol. Metab. 2013, 98, 3532–3542.
- Genchi, V.A.; Rossi, E.; Lauriola, C.; D’Oria, R.; Palma, G.; Borrelli, A.; Caccioppoli, C.; Giorgino, F.; Cignarelli, A. Adipose Tissue Dysfunction and Obesity-Related Male Hypogonadism. Int. J. Mol. Sci. 2022, 23, 8194.
- Salas-Huetos, A.; Maghsoumi-Norouzabad, L.; James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Becerra-Tomás, N.; Javid, A.Z.; Abed, R.; Torres, P.J.; et al. Male adiposity, sperm parameters and reproductive hormones: An updated systematic review and collaborative meta-analysis. Obes. Rev. 2021, 22, e13082.
- Giagulli, V.A.; Kaufman, J.M.; Vermeulen, A. Pathogenesis of the decreased androgen levels in obese men. J. Clin. Endocrinol. Metab. 1994, 79, 997–1000.
- Tchernof, A.; Després, J.P.; Bélanger, A.; Dupont, A.; Prud’homme, D.; Moorjani, S.; Lupien, P.J.; Labrie, F. Reduced testosterone and adrenal C19 steroid levels in obese men. Metabolism 1995, 44, 513–519.
- Navarro, V.M. Metabolic regulation of kisspeptin—The link between energy balance and reproduction. Nat. Rev. Endocrinol. 2020, 16, 407–420.
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The role of kisspeptin neurons in reproduction and metabolism. J. Endocrinol. 2018, 238, R173–R183.
- Kolb, B.A.; Stanczyk, F.Z.; Sokol, R.Z. Serum inhibin B levels in males with gonadal dysfunction. Fertil. Steril. 2000, 74, 234–238.
- Meachem, S.J.; Nieschlag, E.; Simoni, M. Inhibin B in male reproduction: Pathophysiology and clinical relevance. Eur. J. Endocrinol. 2001, 145, 561–571.
- Andersson, A.M.; Skakkebaek, N.E. Serum inhibin B levels during male childhood and puberty. Mol. Cell. Endocrinol. 2001, 180, 103–107.
- Hui, H.; Farilla, L.; Merkel, P.; Perfetti, R. The short half-life of glucagon-like peptide-1 in plasma does not reflect its long-lasting beneficial effects. Eur. J. Endocrinol. 2002, 146, 863–869.
- Cannarella, R.; Calogero, A.E.; Condorelli, R.A.; Greco, E.A.; Aversa, A.; La Vignera, S. Is there a role for glucagon-like peptide-1 receptor agonists in the treatment of male infertility? Andrology 2021, 9, 1499–1503.

