 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Wei Wia | -- | 2767 | 2023-12-06 14:44:10 | | | |
| 2 | Wendy Huang | Meta information modification | 2767 | 2023-12-07 13:09:18 | | |
Video Upload Options
Peritoneal metastasis (PM) is a common mode of distant metastasis in colorectal cancer (CRC) and has a poorer prognosis compared to other metastatic sites. The formation of PM foci depends on the synergistic effect of multiple molecules and the modulation of various components of the tumor microenvironment. The current treatment of CRC-PM is based on systemic chemotherapy. However, recent developments in local therapeutic modalities, such as cytoreductive surgery (CRS) and intraperitoneal chemotherapy (IPC), have improved the survival of these patients.
1. Introduction
Colorectal cancer (CRC) is one of the most common malignancies, with about 1,881,000 new cases and 916,000 deaths from the disease worldwide every year [1], posing a serious threat to human health. Distant metastases are the leading cause of death due to treatment failure. The most likely sites for CRC to metastasize are the liver and lung [2], and the peritoneum is also a common site for CRC metastasis. Compared with the liver and lung, peritoneal metastasis (PM) in CRC cases usually means that patients lose the possibility of conversion therapy and have a worse prognosis. It has been reported that PM is discovered in approximately 13% of CRC cases, with 2% involving isolated PM and the remaining 11% being peritoneal and combined with other organs [3]. Patients who suffer from PM develop a higher proportion of BRAF mutations and worse survival. Due to the lack of effective treatment, the average survival of such patients was shorter than patients with liver-isolated metastasis (16.3 months (13.5–18.8) vs. 19.1 months (18.3–19.8)). Furthermore, PM combined with ≥2 sites of metastases has a 1.4-fold increased risk of death compared with PM alone [3].
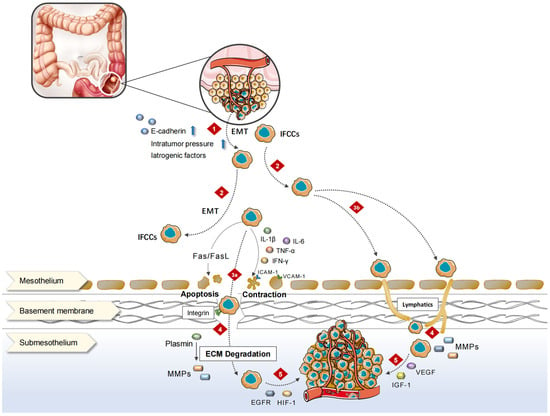
The “seed and soil” idea states that tumor cells interact with milky spots, the peritoneal surface features, to create a milieu for tumor cell colonization and dissemination [4]. Based on the generation time of the primary lesion and PM, it can be divided into synchronous PM and metachronous PM. The former is defined as PM that is detected at the time of diagnosis of CRC or observed within 30 days after primary tumor resection; the latter is defined as PM observed 30 days after radical colorectal resection [5]. The spilling of cancer cells during tumor growth causes synchronous PM. Besides the intraperitoneal free cancer cells (IFCCs) caused by tumor itself, medical variables during surgery can shed cancer cells into the peritoneal cavity, causing metachronous PM.
2. Pathophysiological Process
3. Molecular Biology Characterization
3.1. High-Frequency Mutations in PM
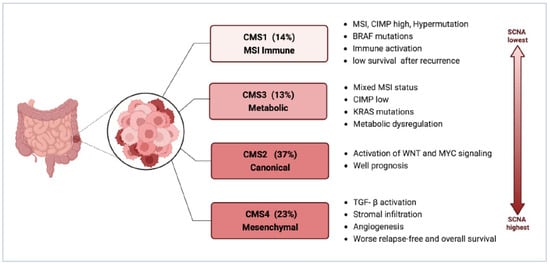
3.2. Consensus Molecular Subtypes (CMS)
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249.
- Luo, D.; Liu, Q.; Yu, W.; Ma, Y.; Zhu, J.; Lian, P.; Cai, S.; Li, Q.; Li, X. Prognostic value of distant metastasis sites and surgery in stage IV colorectal cancer: A population-based study. Int. J. Color. Dis. 2018, 33, 1241–1249.
- Franko, J.; Shi, Q.; Meyers, J.P.; Maughan, T.S.; Adams, R.A.; Seymour, M.T.; Saltz, L.; Punt, C.J.A.; Koopman, M.; Tournigand, C.; et al. Prognosis of patients with peritoneal metastatic colorectal cancer given systemic therapy: An analysis of individual patient data from prospective randomised trials from the Analysis and Research in Cancers of the Digestive System (ARCAD) database. Lancet Oncol. 2016, 17, 1709–1719.
- Fidler, I.J. The pathogenesis of cancer metastasis: The ‘seed and soil’ hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458.
- Segelman, J.; Granath, F.; Holm, T.; Machado, M.; Mahteme, H.; Martling, A. Incidence, prevalence and risk factors for peritoneal carcinomatosis from colorectal cancer. Br. J. Surg. 2012, 99, 699–705.
- Ceelen, W.P.; Bracke, M.E. Peritoneal minimal residual disease in colorectal cancer: Mechanisms, prevention, and treatment. Lancet Oncol. 2009, 10, 72–79.
- Sleeman, J.; Steeg, P.S. Cancer metastasis as a therapeutic target. Eur. J. Cancer 2010, 46, 1177–1180.
- Pandya, P.; Orgaz, J.L.; Sanz-Moreno, V. Actomyosin contractility and collective migration: May the force be with you. Curr. Opin. Cell Biol. 2017, 48, 87–96.
- Lim, S.H.; Becker, T.M.; Chua, W.; Ng, W.L.; de Souza, P.; Spring, K.J. Circulating tumour cells and the epithelial mesenchymal transition in colorectal cancer. J. Clin. Pathol. 2014, 67, 848–853.
- Sugarbaker, P.H. Observations concerning cancer spread within the peritoneal cavity and concepts supporting an ordered pathophysiology. Cancer Res. Treat. 1996, 82, 79–100.
- Mikuła-Pietrasik, J.; Sosińska, P.; Maksin, K.; Kucińska, M.G.; Piotrowska, H.; Murias, M.; Woźniak, A.; Szpurek, D.; Książek, K. Colorectal cancer-promoting activity of the senescent peritoneal mesothelium. Oncotarget 2015, 6, 29178–29195.
- Heldin, P.; Kolliopoulos, C.; Lin, C.Y.; Heldin, C.H. Involvement of hyaluronan and CD44 in cancer and viral infections. Cell Signal 2020, 65, 109427.
- Heath, R.M.; Jayne, D.G.; O’Leary, R.; Morrison, E.E.; Guillou, P.J. Tumour-induced apoptosis in human mesothelial cells: A mechanism of peritoneal invasion by Fas Ligand/Fas interaction. Br. J. Cancer 2004, 90, 1437–1442.
- Lee, I.K.; Vansaun, M.N.; Shim, J.H.; Matrisian, L.M.; Gorden, D.L. Increased metastases are associated with inflammation and matrix metalloproteinase-9 activity at incision sites in a murine model of peritoneal dissemination of colorectal cancer. J. Surg. Res. 2013, 180, 252–259.
- Kataoka, H.; Tanaka, H.; Nagaike, K.; Uchiyama, S.; Itoh, H. Role of cancer cell-stroma interaction in invasive growth of cancer cells. Hum. Cell 2003, 16, 1–14.
- Kim, T.D.; Song, K.S.; Li, G.; Choi, H.; Park, H.D.; Lim, K.; Hwang, B.D.; Yoon, W.H. Activity and expression of urokinase-type plasminogen activator and matrix metalloproteinases in human colorectal cancer. BMC Cancer 2006, 6, 211.
- Nataraj, N.B.; Marrocco, I.; Yarden, Y. Roles for growth factors and mutations in metastatic dissemination. Biochem. Soc. Trans. 2021, 49, 1409–1423.
- Manalo, D.J.; Rowan, A.; Lavoie, T.; Natarajan, L.; Kelly, B.D.; Ye, S.Q.; Garcia, J.G.; Semenza, G.L. Transcriptional regulation of vascular endothelial cell responses to hypoxia by HIF-1. Blood 2005, 105, 659–669.
- Shweiki, D.; Itin, A.; Soffer, D.; Keshet, E. Vascular endothelial growth factor induced by hypoxia may mediate hypoxia-initiated angiogenesis. Nature 1992, 359, 843–845.
- Gong, J.; Lin, Y.; Zhang, H.; Liu, C.; Cheng, Z.; Yang, X.; Zhang, J.; Xiao, Y.; Sang, N.; Qian, X.; et al. Reprogramming of lipid metabolism in cancer-associated fibroblasts potentiates migration of colorectal cancer cells. Cell Death Dis. 2020, 11, 267.
- Peng, S.; Chen, D.; Cai, J.; Yuan, Z.; Huang, B.; Li, Y.; Wang, H.; Luo, Q.; Kuang, Y.; Liang, W.; et al. Enhancing cancer-associated fibroblast fatty acid catabolism within a metabolically challenging tumor microenvironment drives colon cancer peritoneal metastasis. Mol. Oncol. 2021, 15, 1391–1411.
- Peng, S.; Li, Y.; Huang, M.; Tang, G.; Xie, Y.; Chen, D.; Hu, Y.; Yu, T.; Cai, J.; Yuan, Z.; et al. Metabolomics reveals that CAF-derived lipids promote colorectal cancer peritoneal metastasis by enhancing membrane fluidity. Int. J. Biol. Sci. 2022, 18, 1912–1932.
- The Cancer Genome Atlas Network. Comprehensive molecular characterization of human colon and rectal cancer. Nature 2012, 487, 330–337.
- Schneider, M.A.; Eden, J.; Pache, B.; Laminger, F.; Lopez-Lopez, V.; Steffen, T.; Hübner, M.; Kober, F.; Roka, S.; Campos, P.C.; et al. Mutations of RAS/RAF Proto-oncogenes Impair Survival After Cytoreductive Surgery and HIPEC for Peritoneal Metastasis of Colorectal Origin. Ann. Surg. 2018, 268, 845–853.
- Graf, W.; Cashin, P.H.; Ghanipour, L.; Enblad, M.; Botling, J.; Terman, A.; Birgisson, H. Prognostic Impact of BRAF and KRAS Mutation in Patients with Colorectal and Appendiceal Peritoneal Metastases Scheduled for CRS and HIPEC. Ann. Surg. Oncol. 2020, 27, 293–300.
- Baratti, D.; Kusamura, S.; Niger, M.; Perrone, F.; Milione, M.; Cattaneo, L.; Guaglio, M.; Bartolini, V.; Pietrantonio, F.; Deraco, M. Prognostic Impact of Primary Side and RAS/RAF Mutations in a Surgical Series of Colorectal Cancer with Peritoneal Metastases. Ann. Surg. Oncol. 2021, 28, 3332–3342.
- Stein, M.K.; Williard, F.W.; Xiu, J.; Tsao, M.W.; Martin, M.G.; Deschner, B.W.; Dickson, P.V.; Glazer, E.S.; Yakoub, D.; Shibata, D.; et al. Comprehensive tumor profiling reveals unique molecular differences between peritoneal metastases and primary colorectal adenocarcinoma. J. Surg. Oncol. 2020, 121, 1320–1328.
- Ubink, I.; van Eden, W.J.; Snaebjornsson, P.; Kok, N.F.M.; van Kuik, J.; van Grevenstein, W.M.U.; Laclé, M.M.; Sanders, J.; Fijneman, R.J.A.; Elias, S.G.; et al. Histopathological and molecular classification of colorectal cancer and corresponding peritoneal metastases. Br. J. Surg. 2018, 105, e204–e211.
- Ogino, S.; Nosho, K.; Kirkner, G.J.; Kawasaki, T.; Meyerhardt, J.A.; Loda, M.; Giovannucci, E.L.; Fuchs, C.S. CpG island methylator phenotype, microsatellite instability, BRAF mutation and clinical outcome in colon cancer. Gut 2009, 58, 90–96.
- Flood, M.P.; Jain, A.; Mitchell, C.; Hewitt, C.; Ramsay, R.; Michael, M.; Heriot, A.G.; Tie, J. The impact of molecular and mismatch repair status on the survival outcomes of surgically treated patients with colorectal peritoneal metastases. Eur. J. Surg. Oncol. 2022, 48, 2218–2225.
- Larsen, S.G.; Goscinski, M.A.; Dueland, S.; Steigen, S.E.; Hofsli, E.; Torgunrud, A.; Lund-Iversen, M.; Dagenborg, V.J.; Flatmark, K.; Sorbye, H. Impact of KRAS, BRAF and microsatellite instability status after cytoreductive surgery and HIPEC in a national cohort of colorectal peritoneal metastasis patients. Br. J. Cancer 2022, 126, 726–735.
- He, K.; Wang, Y.; Zhong, Y.; Pan, X.; Si, L.; Lu, J. KRAS Codon 12 Mutation is Associated with More Aggressive Invasiveness in Synchronous Metastatic Colorectal Cancer (mCRC): Retrospective Research. OncoTargets Ther. 2020, 13, 12601–12613.
- Arjona-Sanchez, A.; Rodriguez-Ortiz, L.; Baratti, D.; Schneider, M.A.; Gutiérrez-Calvo, A.; García-Fadrique, A.; Tuynman, J.B.; Cascales-Campos, P.A.; Martín, V.C.; Morales, R.; et al. RAS Mutation Decreases Overall Survival After Optimal Cytoreductive Surgery and Hyperthermic Intraperitoneal Chemotherapy of Colorectal Peritoneal Metastasis: A Modification Proposal of the Peritoneal Surface Disease Severity Score. Ann. Surg. Oncol. 2019, 26, 2595–2604.
- Breuer, E.; Hebeisen, M.; Schneider, M.A.; Roth, L.; Pauli, C.; Frischer-Ordu, K.; Eden, J.; Pache, B.; Steffen, T.; Hübner, M.; et al. Site of Recurrence and Survival After Surgery for Colorectal Peritoneal Metastasis. J. Natl. Cancer Inst. 2021, 113, 1027–1035.
- Tran, B.; Kopetz, S.; Tie, J.; Gibbs, P.; Jiang, Z.Q.; Lieu, C.H.; Agarwal, A.; Maru, D.M.; Sieber, O.; Desai, J. Impact of BRAF mutation and microsatellite instability on the pattern of metastatic spread and prognosis in metastatic colorectal cancer. Cancer 2011, 117, 4623–4632.
- Kim, C.G.; Ahn, J.B.; Jung, M.; Beom, S.H.; Kim, C.; Kim, J.H.; Heo, S.J.; Park, H.S.; Kim, J.H.; Kim, N.K.; et al. Effects of microsatellite instability on recurrence patterns and outcomes in colorectal cancers. Br. J. Cancer 2016, 115, 25–33.
- Tonello, M.; Baratti, D.; Sammartino, P.; Di Giorgio, A.; Robella, M.; Sassaroli, C.; Framarini, M.; Valle, M.; Macrì, A.; Graziosi, L.; et al. Microsatellite and RAS/RAF Mutational Status as Prognostic Factors in Colorectal Peritoneal Metastases Treated with Cytoreductive Surgery and Hyperthermic Intraperitoneal Chemotherapy (HIPEC). Ann. Surg. Oncol. 2022, 29, 3405–3417.
- Christensen, T.D.; Palshof, J.A.; Larsen, F.O.; Poulsen, T.S.; Høgdall, E.; Pfeiffer, P.; Jensen, B.V.; Yilmaz, M.K.; Nielsen, D. Associations between primary tumor RAS, BRAF and PIK3CA mutation status and metastatic site in patients with chemo-resistant metastatic colorectal cancer. Acta Oncol. 2018, 57, 1057–1062.
- Peerenboom, R.; Dhiman, A.; Witmer, H.D.D.; Spurr, L.F.; Polite, B.; Eng, O.S.; Shergill, A.; Turaga, K.K. PI3K Pathway Alterations in Peritoneal Metastases are Associated with Earlier Recurrence in Patients with Colorectal Cancer Undergoing Optimal Cytoreductive Surgery. Ann. Surg. Oncol. 2023, 30, 3114–3122.
- Mlecnik, B.; Bindea, G.; Kirilovsky, A.; Angell, H.K.; Obenauf, A.C.; Tosolini, M.; Church, S.E.; Maby, P.; Vasaturo, A.; Angelova, M.; et al. The tumor microenvironment and Immunoscore are critical determinants of dissemination to distant metastasis. Sci. Transl. Med. 2016, 8, 327ra326.
- Yaeger, R.; Chatila, W.K.; Lipsyc, M.D.; Hechtman, J.F.; Cercek, A.; Sanchez-Vega, F.; Jayakumaran, G.; Middha, S.; Zehir, A.; Donoghue, M.T.A.; et al. Clinical Sequencing Defines the Genomic Landscape of Metastatic Colorectal Cancer. Cancer Cell 2018, 33, 125–136.
- Siesing, C.; Petersson, A.; Ulfarsdottir, T.; Chattopadhyay, S.; Nodin, B.; Eberhard, J.; Brändstedt, J.; Syk, I.; Gisselsson, D.; Jirström, K. Delineating the intra-patient heterogeneity of molecular alterations in treatment-naïve colorectal cancer with peritoneal carcinomatosis. Mod. Pathol. 2022, 35, 979–988.
- Diep, C.B.; Teixeira, M.R.; Thorstensen, L.; Wiig, J.N.; Eknaes, M.; Nesland, J.M.; Giercksky, K.E.; Johansson, B.; Lothe, R.A. Genome characteristics of primary carcinomas, local recurrences, carcinomatoses, and liver metastases from colorectal cancer patients. Mol. Cancer 2004, 3, 6.
- Kleivi, K.; Lind, G.E.; Diep, C.B.; Meling, G.I.; Brandal, L.T.; Nesland, J.M.; Myklebost, O.; Rognum, T.O.; Giercksky, K.E.; Skotheim, R.I.; et al. Gene expression profiles of primary colorectal carcinomas, liver metastases, and carcinomatoses. Mol. Cancer 2007, 6, 2.
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356.
- Song, N.; Pogue-Geile, K.L.; Gavin, P.G.; Yothers, G.; Kim, S.R.; Johnson, N.L.; Lipchik, C.; Allegra, C.J.; Petrelli, N.J.; O'Connell, M.J.; et al. Clinical Outcome From Oxaliplatin Treatment in Stage II/III Colon Cancer According to Intrinsic Subtypes: Secondary Analysis of NSABP C-07/NRG Oncology Randomized Clinical Trial. JAMA Oncol. 2016, 2, 1162–1169.
- Laoukili, J.; Constantinides, A.; Wassenaar, E.C.E.; Elias, S.G.; Raats, D.A.E.; van Schelven, S.J.; van Wettum, J.; Volckmann, R.; Koster, J.; Huitema, A.D.R.; et al. Peritoneal metastases from colorectal cancer belong to Consensus Molecular Subtype 4 and are sensitised to oxaliplatin by inhibiting reducing capacity. Br. J. Cancer 2022, 126, 1824–1833.
- Becht, E.; de Reyniès, A.; Giraldo, N.A.; Pilati, C.; Buttard, B.; Lacroix, L.; Selves, J.; Sautès-Fridman, C.; Laurent-Puig, P.; Fridman, W.H. Immune and Stromal Classification of Colorectal Cancer Is Associated with Molecular Subtypes and Relevant for Precision Immunotherapy. Clin. Cancer Res. 2016, 22, 4057–4066.
- Trinh, A.; Trumpi, K.; De Sousa, E.M.F.; Wang, X.; de Jong, J.H.; Fessler, E.; Kuppen, P.J.; Reimers, M.S.; Swets, M.; Koopman, M.; et al. Practical and Robust Identification of Molecular Subtypes in Colorectal Cancer by Immunohistochemistry. Clin. Cancer Res. 2017, 23, 387–398.
- Barriuso, J.; Nagaraju, R.T.; Belgamwar, S.; Chakrabarty, B.; Burghel, G.J.; Schlecht, H.; Foster, L.; Kilgour, E.; Wallace, A.J.; Braun, M.; et al. Early Adaptation of Colorectal Cancer Cells to the Peritoneal Cavity Is Associated with Activation of “Stemness” Programs and Local Inflammation. Clin. Cancer Res. 2021, 27, 1119–1130.

