Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Dana Antonia Tapoi | -- | 2525 | 2023-11-13 11:04:58 | | | |
| 2 | Rita Xu | Meta information modification | 2525 | 2023-11-14 02:49:58 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Gosman, L.M.; Țăpoi, D.; Costache, M. Emerging Biomarkers in Cutaneous Melanoma. Encyclopedia. Available online: https://encyclopedia.pub/entry/51475 (accessed on 06 June 2026).
Gosman LM, Țăpoi D, Costache M. Emerging Biomarkers in Cutaneous Melanoma. Encyclopedia. Available at: https://encyclopedia.pub/entry/51475. Accessed June 06, 2026.
Gosman, Laura Maria, Dana-Antonia Țăpoi, Mariana Costache. "Emerging Biomarkers in Cutaneous Melanoma" Encyclopedia, https://encyclopedia.pub/entry/51475 (accessed June 06, 2026).
Gosman, L.M., Țăpoi, D., & Costache, M. (2023, November 13). Emerging Biomarkers in Cutaneous Melanoma. In Encyclopedia. https://encyclopedia.pub/entry/51475
Gosman, Laura Maria, et al. "Emerging Biomarkers in Cutaneous Melanoma." Encyclopedia. Web. 13 November, 2023.
Copy Citation
Cutaneous melanoma (CM) is an increasingly significant public health concern. Due to alarming mortality rates and escalating incidence, it is crucial to understand its etiology and identify emerging biomarkers for improved diagnosis and treatment strategies.
cutaneous melanoma
immunohistochemistry
genetic mutations
1. Introduction
Cutaneous melanoma (CM) is a particularly aggressive form of cancer that originates from melanocytes, pigment-producing cells derived from the neural crest [1]. Despite representing a mere 4% of all skin cancers, CM accounts for up to 75% of skin cancer-related deaths [2]. However, with early detection and proper intervention, over 90% of the cases could be cured [3].
The pathogenesis of CM is multifactorial, involving both genetic and environmental factors [4]. Ultraviolet radiation (UVR), either from natural light or artificial sources, is the most important environmental risk factor for CM. Additionally, individuals with lighter complexion have the highest risk of developing CM due to lower levels of melanin which make these individuals more likely to develop sunburns. A higher number of nevi are also associated with an increased risk. A familial history of CM further increases this risk, possibly due to shared sun exposure behaviors or hereditary genetic mutations [5].
Patient survival is strongly correlated with an early detection of the disease. Among various prognostic factors, the depth of invasion remains the most critical determinant of survival in numerous studies. Intraepidermal (in situ) melanomas can be cured by excision alone, and thin melanomas have minimal metastatic potential [6]. On the contrary, thick CMs still have very high mortality rates [7].
2. Melanoma Pathogenesis
Ultraviolet radiation exposure is a major risk factor for CM due to the UVR capacity to damage DNA, causing somatic mutations [8]. The exposure can be classified as intermittent or chronic, the latter being mostly occupational. Both these exposure patterns are associated with an increased risk of CM, but it appears that the risk is higher for intermittent exposure [8][9]. This may be explained by the fact that intermittently exposed individuals have lower melanin levels and are more likely to develop sunburns [10]. Nevertheless, there are cutaneous melanomas, such as acral melanomas, which arise in skin that is not exposed to UVR. In this context, according to the 2023 WHO Classification of Tumors, cutaneous melanomas are classified as melanomas arising in sun-exposed skin and melanomas arising in sun-shielded sites (Table 1) [11].
Table 1. WHO Classification of cutaneous melanomas.
| Melanomas arising in sun-exposed skin | Low CSD melanoma: SSM, low CSD nodular melanoma |
| High CSD melanoma: lentigo malignant melanoma, high CSD nodular melanoma | |
| Desmoplastic melanoma: most often associated with severely sun-damaged skin | |
| Melanomas arising in sun-shielded skin or without known UVR exposure | Spitz melanoma |
| Acral melanoma | |
| Melanoma arising in congenital nevus | |
| Melanoma arising in blue nevus |
Apart from UVR exposure, hereditary predisposition is another risk factor for cutaneous melanomas. However, familial cases encompass around 10% of all melanomas [12]. In this respect, several high-penetrance genes such as CDKN2A, CDK4, or BAP1 are the most mutated in hereditary melanomas [13]. Individuals with germline mutations in the CDKN2A, a tumor suppressor gene, have a very high lifetime risk of developing CM, this mutation being encountered in up to 40% of melanoma-prone families [12]. Nevertheless, these mutations are relatively rare and are responsible for just around 2% of all CM cases [8]. In addition to these high-penetrance genes, some medium-penetrance genes such as MITF and MC1R are also involved in hereditary CM [12]. Furthermore, MC1R can be considered a “melanoma modifier gene” as it also increases the penetrance of CDKN2A [14].
As hereditary melanomas are relatively rare, most cutaneous melanomas are characterized by a remarkably high burden of somatic genetic mutations [15][16]. Identifying these genetic mutations can serve both diagnostic and prognostic purposes. Genetic testing is particularly useful for the diagnosis of dedifferentiated CMs which lack typical morphological and immunohistochemical features. In such cases, the diagnosis can be established by identifying melanoma-specific mutations [17].
The most frequent mutations in CM affect genes involved in the aberrant activation of the RAS/RAF/MEK/ERK signaling pathway, also known as the mitogen-activated protein kinase (MAPK) pathway, and the phosphoinositol-3-kinase (PI3K)/AKT pathway [18]. These mutated genes include BRAF, NRAS, NF1, PTEN, KIT, TP53, CDKN2A, and TERT [19][20].
The MAPK pathway is involved in the transduction of extracellular signals to the nucleus, thus activating genes that regulate cell proliferation and differentiation [21][22]. This aberrant activation is responsible for several cellular dysfunctions, such as the deregulation of the cell cycle and inhibition of apoptosis [21][23][24]. MAPK is the most frequently dysregulated pathway in cutaneous melanoma [25]. Up to 90% of all melanoma cases exhibit an abnormal activation of the MAPK pathway. The second most frequently activated pathway in CM is the PI3K pathway which plays a crucial role in maintaining cellular homeostasis [26][27].
As the MAPK pathway is the most affected in CM, numerous mechanisms contribute to its abnormal signaling, including BRAF mutations [18][28]. Between 37% and 60% of cutaneous melanomas harbor a somatic mutation in this gene, with the highest frequency observed in CM associated with low CSD [29]. The majority of BRAF mutations in cutaneous melanoma are missense, resulting in amino acid substitutions at the valine 600 position. Approximately 80% are V600E mutations (glutamic acid substitution), while 5–12% are V600K mutations (lysine substitution). Less common mutations include V600D (valine to aspartic acid) or V600R (arginine substitution). Additionally, BRAF non-V600 mutations can occur in around 5% of cases [30]. The BRAF gene encodes a protein kinase with three distinct domains: two regulatory and one catalytic. The latter is involved in the phosphorylation of MEK and in maintaining the protein inactive through a hydrophobic interaction. [31]. In the BRAF V600E mutation, the hydrophobic valine residue is substituted by a polar, hydrophilic glutamic acid which induces a conformational change in the catalytic domain, resulting in a constitutively active kinase [32][33]. BRAF non-V600E mutations generally operate through a similar mechanism, enhancing BRAF kinase activity [33]. Acknowledging these mutations is clinically significant for treatment and prognosis. BRAF V600-mutated melanomas can be treated with BRAF/MEK inhibitors, with response rates higher in V600E-mutated cases compared to V600K-mutated cases. Furthermore, even though the evidence is still limited, BRAF non-V600-mutated melanomas may still benefit from BRAF/MEK inhibitors [30].
The second most prevalent cause of aberrant MAPK pathway signaling In cutaneous melanoma is attributed to activating mutations in the NRAS gene. These mutations occur in 15–30% of melanomas and are predominantly missense, most often affecting codon 61 [34][35]. These mutations perpetuate aberrant signaling through both the MAPK and PI3K pathways [18][36][37]. It is noteworthy that NRAS and BRAF mutations are generally considered to be mutually exclusive, although co-mutations have been observed in rare instances [37]. NRAS- and NRAS-BRAF-co-mutated melanomas have a less favorable prognosis than BRAF-mutated ones as there are no target therapies for NRAS mutations [17].
Neurofibromin 1 (NF1) is a tumor suppressor gene, mutated in 10–15% of CM, making it the third most common mutation in this pathology [38][39]. NF1 alterations are more frequent in melanomas associated with high CSD. These cases tend to possess a high mutational burden, including a co-occurrence of BRAF or NRAS mutations [19][40]. The NF1 protein serves as a regulator of the RAS family, attenuating downstream RAS signaling [41]. Consequently, loss-of-function mutations in NF1 result in the hyperactivation of NRAS, leading to increased signaling through both the MAPK and PI3K pathways [19][38][39][41]. Analyzing NF1 mutation status has some prognostic value even though there are no target therapies for NF1-mutated melanomas, but such cases respond favorably to immunotherapy [42]. Moreover, NF1 analysis can offer important diagnostic information as this mutation is particularly common in dedifferentiated lesions which can be difficult to diagnose otherwise [17].
The receptor tyrosine kinase KIT plays a crucial physiological role in the proliferation and survival of melanoma cells, through the PI3K and the MAPK signaling cascades. KIT mutations are found in 2–8% of melanoma cases and are more common in acral melanomas and melanomas associated with low CSD [43][44]. Recognizing these mutations is important as such cases can benefit from tyrosine kinase inhibitors [45].
Mutations in the TERT promoter confer a proliferative advantage to melanoma cells and are common in advanced disease, being associated with a less favorable prognosis. Nevertheless, this mutation could become a potential therapeutic target [46][47]. TP53-mutated melanomas are also associated with advanced disease [46]. Assessing the status of TP53 is important as these mutations have been associated with MAPK inhibitor resistance but they can also become potential therapeutic targets [48][49].
The PTEN gene, a tumor suppressor gene, is commonly dysregulated in the vertical growth phase of melanoma and in metastatic lesions, occurring in 10–30% of cutaneous melanomas [18][50]. PTEN alterations tend to be mutually exclusive with NRAS mutations but often co-occur with mutations in BRAF [51][52]. This co-occurrence has been hypothesized to increase PI3K pathway activation [51][52], mimicking the effects of an NRAS-only activation [51][53]. Additionally, PTEN loss-of-function is involved in acquired resistance to BRAF inhibitors in BRAF-mutated melanomas [54]. As mentioned before, BRAF-mutated melanomas may respond to BRAF inhibitors. However, therapeutic success is often temporary, as patients usually experience disease progression at some point or may even exhibit primary resistance to this target therapy. In this respect, acquired genetic mutations affecting the MAPK and PI3K signaling pathways play a central role in resistance to both chemotherapy and targeted therapies [55][56][57]. In this context, targeted PTEN therapy could improve the outcomes of the patients [49]. Having taken everything into consideration, due to this extraordinary genetic heterogeneity of melanomas, a multi-faced diagnostic and therapeutic approach including the identification of molecular biomarkers and genetic aberrations is imperative for optimizing patient outcomes.
3. Emerging Biomarkers in CM
Due to the extraordinarily heterogenous histopathological, immunohistochemical, and molecular landscape of CM, this disease continues to pose important challenges in terms of diagnosis and treatment. Therefore, several new biomarkers have become increasingly studied in recent years in the hope of improving the understanding of CM pathogenesis and management. Unlike immunohistochemical and genetic testing, these emerging biomarkers are expected to improve the early detection and subsequent monitoring of CM in rapid, cost-effective, and non-invasive ways as they can easily be analyzed from blood samples. Furthermore, these new biomarkers may also serve as potential therapeutic targets.
3.1. MicroRNA
MicroRNA (miRNAs) represent non-coding RNAs involved in degrading mRNAs [58]. They have been increasingly recognized as critical modulators of oncogenic processes, including various stages of cancer progression such as melanoma [59][60]. In melanomas, miRNA dysregulation is involved in promoting cell proliferation, resistance to apoptosis and invasion, angiogenesis, and metastasis [61]. Additionally, miRNAs have also been associated with resistance to BRAF and MAPK inhibitors [62][63]. In this context, due to their detectability in both intra- and extracellular compartments and their stable levels even in unfavorable conditions, miRNAs have attracted considerable attention as emerging biomarkers in oncology [64].
At present, miRNA levels can be assessed from various sources, such as resected primary or metastatic tumors, as well as arterial or venous plasma and serum. Importantly, the data derived from these different sources have shown no significant divergence. Depending on the type of cancer under investigation, abnormal miRNA expression profiles have been found to correlate with various disease stages, overall prognosis, tumor recurrence, and potential responsiveness to therapeutic interventions [59][65]. In patients with melanoma, different miRNAs can be either up- or down-regulated, and have been correlated with progression-free survival and overall survival [66][67][68]. Interestingly, serum levels of various microRNAs can discriminate between melanoma stages with increased accuracy compared to S100B or LDH [69]. For instance, miR-137, miR-148, and miR-182 downregulate MITF expression and promote tumor invasion [70]. miR-221 plasma levels are increased in melanoma patients, and are correlated with stage, recurrence, and disease progression [71]. Rigg E. et al. demonstrated that miR-146a-5p is overexpressed in melanoma brain metastases and its knockdown results in a reduction of metastatic lesions [72]. On the contrary, other types of microRNA such as mirR-211, miR-542 3p, or miR-152-3p are downregulated in invasive melanomas [73]. In vitro studies demonstrated that increasing miR-152-3p expression inhibits the proliferation and invasiveness of melanoma cells [74]. miR-542 3p is involved in epithelial-to-mesenchymal transition (ETM), and its experimental upregulation inhibited ETM and metastatic spread [75]. miR-143 also bears anti-tumoral effects as it has been linked to promoting apoptosis and inhibiting the proliferation of melanoma cells [76]. Similar anti-tumoral effects have been reported for miR-224-5p which can additionally overturn acquired resistance to BRAF inhibitors [77]. Several other microRNA seem to play a role in resistance to target or conventional chemotherapy. A downregulation of miR-7, miR-579 3p, and miR-126 3p was found in melanomas resistant to BRAF/MAPK inhibitors [78][79][80], while miR-31 downregulation is associated with chemoresistance [81]. Importantly, the experimental upregulation of miR-7 and miR-126 3p restored responses to BRAF inhibitors in melanoma cell lines [78][79].
3.2. Exosomes
Exosomes are extracellular vesicles secreted by cells, encompassing a unique molecular signature that reflects the cell type from which they originate. Given their traceable cellular origins and facile isolation, exosomes can be regarded as potential biomarkers for diagnosis and prognosis in various cancers, including melanoma [82]. They are readily available through minimally invasive methods, as they can be isolated from a variety of biological fluids such as blood, plasma, urine, and cerebrospinal fluid [83]. Exosomes extracted from melanoma cell lines have been shown to contain distinct mRNA, miRNA, and protein profiles [82][83]. Exosome analysis can offer important diagnostic and prognostic information, as various exosomal components are significantly altered in cutaneous melanomas [82][84][85]. In this respect, Surman M. et al. found increased exosome concentrations in melanoma cases but those levels were not correlated with disease stage [82]. On the contrary, Boussadia Z. et al. reported a higher exosome concentration in metastatic melanoma compared to non-metastatic cases [86]. The complex relationship between exosomal components and melanoma progression is not entirely understood, but various mechanisms have been proposed. For instance, exosomes can carry and modulate the activity of matrix metalloproteinases (MMPs), as well as alter cell adhesion and activate fibroblasts to become cancer-associated fibroblasts, thus stimulating melanoma invasiveness [82][87][88][89]. Exosomal components have also been shown to enhance metastatic potential in CM by promoting epithelial-to-mesenchymal transition (ETM) [90], angiogenesis [82][91], and lymphangiogenesis [92]. As melanoma is particularly prone to brain metastases, exosomes may also at least partially explain this characteristic by damaging endothelial cells and the blood–brain barrier, and activating glial cells [93].
Some exosomal components have also been linked to resistance to therapy in CM, and targeting these molecules may improve therapeutic response [94]. In this context, exosomes can influence the melanoma microenvironment by altering the function of lymphocytes and stimulating tumor-associated macrophages (TAMs) to become M2-polarized and secrete pro-tumorigenic cytokines [95][96][97]. These effects can affect the response to immunotherapy, and targeting TAMs could improve the outcome of melanoma patients [98]. Furthermore, exosomes can be used as a means for administering therapy [99]. In this respect, exosomes containing BRAF siRNA were shown to have increased anti-tumoral activity compared to siBRAF in melanoma cell lines [100]. Similarly, cord-blood-derived exosomes produced significant genotoxicity and a decrease in survival time for melanoma cells and lymphocytes from melanoma patients, apparently by delivering anti-oncogenic miR-7. These results are particularly important as the exosome caused no significant damage to normal lymphocytes [101]. While these findings underscore the promising role of exosomes as diagnostic, prognostic, and treatment tools in melanoma, additional research is required to comprehensively delineate their utility. Future studies may aim to validate these biomarkers in larger patient cohorts, completely elucidate the roles of exosomal components in melanoma progression, and assess the feasibility of incorporating exosomal markers into existing diagnostic, prognostic, and therapeutic frameworks.
The complex effects of exosomal components in CM pathogenesis are presented in Figure 1.
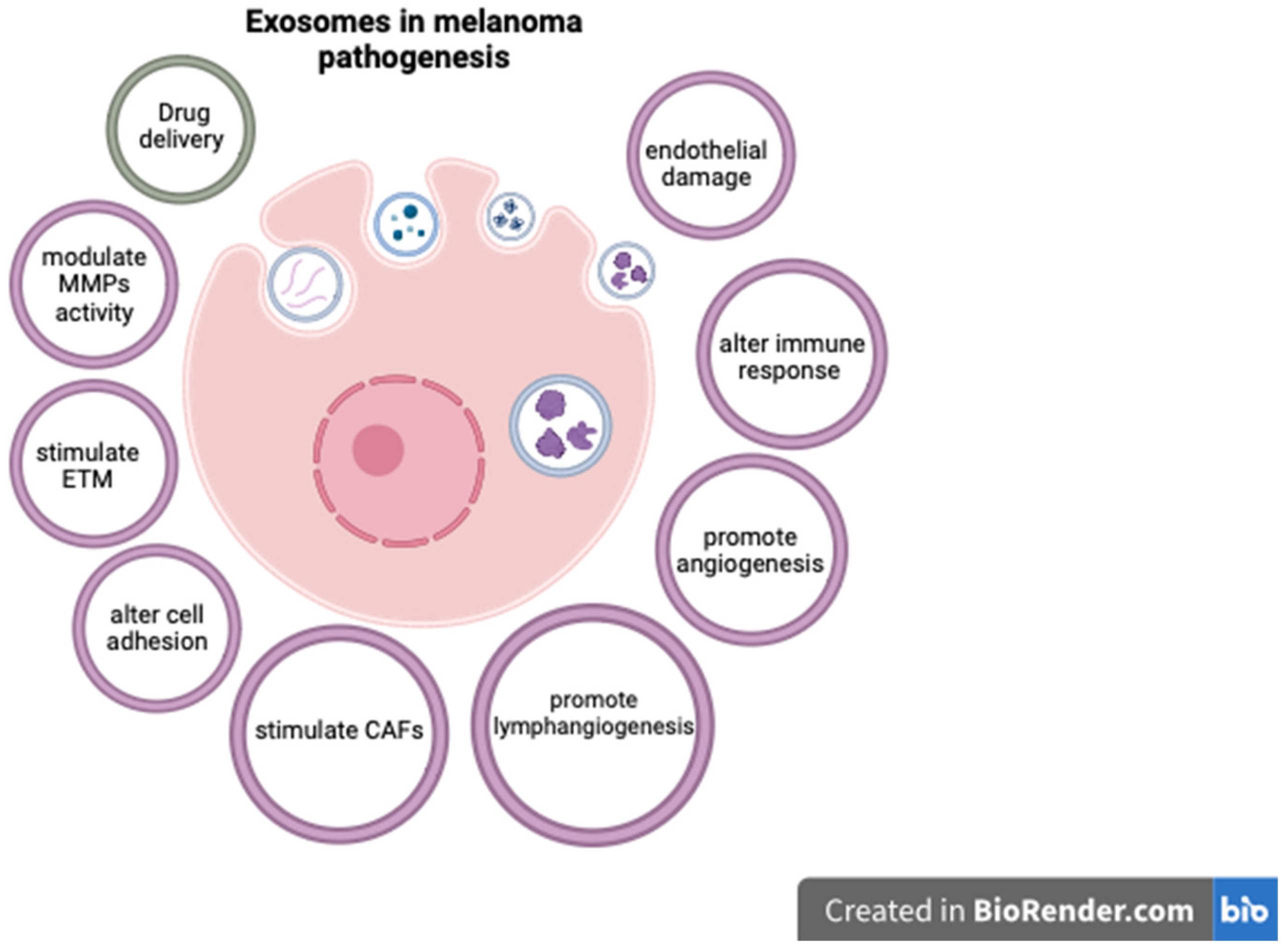
Figure 1. The role of exosomal components in CM.
References
- Leonardi, G.C.; Falzone, L.; Salemi, R.; Zanghì, A.; Spandidos, D.A.; Mccubrey, J.A.; Candido, S.; Libra, M. Cutaneous melanoma: From pathogenesis to therapy (Review). Int. J. Oncol. 2018, 52, 1071–1080.
- Davis, L.E.; Shalin, S.C.; Tackett, A.J. Current state of melanoma diagnosis and treatment. Cancer Biol. Ther. 2019, 20, 1366–1379.
- Orzan, O.A.; Șandru, A.; Jecan, C.R. Controversies in the diagnosis and treatment of early cutaneous melanoma. J. Med. Life 2015, 8, 132–141.
- Caini, S.; Gandini, S.; Sera, F.; Raimondi, S.; Fargnoli, M.C.; Boniol, M.; Armstrong, B.K. Meta-analysis of risk factors for cutaneous melanoma according to anatomical site and clinico-pathological variant. Eur. J. Cancer. 2009, 45, 3054–3063.
- Burns, D.; George, J.; Aucoin, D.; Bower, J.; Burrell, S.; Gilbert, R.; Bower, N. The Pathogenesis and Clinical Management of Cutaneous Melanoma: An Evidence-Based Review. J. Med. Imaging Radiat. Sci. 2019, 50, 460–469.e1.
- Rigel, D.S.; Carucci, J.A. Malignant melanoma: Prevention, early detection, and treatment in the 21st century. CA Cancer J. Clin. 2000, 50, 215–236.
- Țăpoi, D.A.; Derewicz, D.; Gheorghișan-Gălățeanu, A.-A.; Dumitru, A.V.; Ciongariu, A.M.; Costache, M. The Impact of Clinical and Histopathological Factors on Disease Progression and Survival in Thick Cutaneous Melanomas. Biomedicines 2023, 11, 2616.
- Cust, A.E.; Mishra, K.; Berwick, M. Melanoma—Role of the environment and genetics. Photochem. Photobiol. Sci. 2018, 17, 1853–1860.
- Gandini, S.; Sera, F.; Cattaruzza, M.S.; Pasquini, P.; Picconi, O.; Boyle, P.; Melchi, C.F. Meta-analysis of risk factors for cutaneous melanoma: II. Sun exposure. Eur. J. Cancer. 2005, 41, 45–60.
- de Gruijl, F.R. UV adaptation: Pigmentation and protection against overexposure. Exp. Dermatol. 2017, 26, 557–562.
- Elder, D.E.; Barnhill, R.Y. (Eds.) Chapter III: Melanocytic neoplasms. In WHO Classification of Tumours Editorial Board. Skin Tumours, 5th ed.; Forthcoming; International Agency for Research on Cancer: Lyon, France, 2023; Volume 12, Available online: https://tumourclassification.iarc.who.int/chapters/64 (accessed on 27 August 2023).
- Ribeiro Moura Brasil Arnaut, J.; Dos Santos Guimarães, I.; Evangelista Dos Santos, A.C.; de Moraes Lino da Silva, F.; Machado, J.R.; de Melo, A.C. Molecular landscape of Hereditary Melanoma. Crit. Rev. Oncol. Hematol. 2021, 164, 103425.
- Tímár, J.; Ladányi, A. Molecular Pathology of Skin Melanoma: Epidemiology, Differential Diagnostics, Prognosis and Therapy Prediction. Int. J. Mol. Sci. 2022, 23, 5384.
- Cakir, A.; Elcin, G.; Kilickap, S.; Gököz, Ö.; Taskiran, Z.E.; Celik, İ. Phenotypic and Genetic Features that Differ Between Hereditary and Sporadic Melanoma: Results of a Preliminary Study from a Single Center from Turkey. Dermatol. Pract. Concept. 2023, 13, e2023146.
- Vogelstein, B.; Papadopoulos, N.; Velculescu, V.E.; Zhou, S.; Diaz, L.A., Jr.; Kinzler, K.W. Cancer genome landscapes. Science 2013, 339, 1546–1558.
- Cancer Genome Atlas Network. Genomic Classification of Cutaneous Melanoma. Cell 2015, 161, 1681–1696.
- Țăpoi, D.A.; Gheorghișan-Gălățeanu, A.-A.; Dumitru, A.V.; Ciongariu, A.M.; Furtunescu, A.R.; Marin, A.; Costache, M. Primary Undifferentiated/Dedifferentiated Cutaneous Melanomas—A Review on Histological, Immunohistochemical, and Molecular Features with Emphasis on Prognosis and Treatment. Int. J. Mol. Sci. 2023, 24, 9985.
- Chappell, W.H.; Steelman, L.S.; Long, J.M.; Kempf, R.C.; Abrams, S.L.; Franklin, R.A.; Bäsecke, J.; Stivala, F.; Donia, M.; Fagone, P.; et al. Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR inhibitors: Rationale and importance to inhibiting these pathways in human health. Oncotarget 2011, 2, 135–164.
- Hodis, E.; Watson, I.R.; Kryukov, G.V.; Arold, S.T.; Imielinski, M.; Theurillat, J.P.; Nickerson, E.; Auclair, D.; Li, L.; Place, C.; et al. A landscape of driver mutations in melanoma. Cell 2012, 150, 251–263.
- Krauthammer, M.; Kong, Y.; Ha, B.H.; Evans, P.; Bacchiocchi, A.; McCusker, J.P.; Cheng, E.; Davis, M.J.; Goh, G.; Choi, M.; et al. Exome sequencing identifies recurrent somatic RAC1 mutations in melanoma. Nat. Genet. 2012, 44, 1006–1014.
- Wellbrock, C.; Karasarides, M.; Marais, R. The RAF proteins take centre stage. Nat. Rev. Mol. Cell. Biol. 2004, 5, 875–885.
- Raman, M.; Chen, W.; Cobb, M.H. Differential regulation and properties of MAPKs. Oncogene 2007, 26, 3100–3112.
- Cohen, C.; Zavala-Pompa, A.; Sequeira, J.H.; Shoji, M.; Sexton, D.G.; Cotsonis, G.; Cerimele, F.; Govindarajan, B.; Macaron, N.; Arbiser, J.L. Mitogen-actived protein kinase activation is an early event in melanoma progression. Clin. Cancer Res. 2002, 8, 3728–3733.
- Wang, Y.F.; Jiang, C.C.; Kiejda, K.A.; Gillespie, S.; Zhang, X.D.; Hersey, P. Apoptosis induction in human melanoma cells by inhibition of MEK is caspase-independent and mediated by the Bcl-2 family members PUMA, Bim, and Mcl-1. Clin. Cancer Res. 2007, 13, 4934–4942.
- Carlino, M.S.; Long, G.V.; Kefford, R.F.; Rizos, H. Targeting oncogenic BRAF and aberrant MAPK activation in the treatment of cutaneous melanoma. Crit. Rev. Oncol. Hematol. 2015, 96, 385–398.
- Yuan, T.L.; Cantley, L.C. PI3K pathway alterations in cancer: Variations on a theme. Oncogene 2008, 27, 5497–5510.
- Davies, M.A. The role of the PI3K-AKT pathway in melanoma. Cancer J. 2012, 18, 142–147.
- Gray-Schopfer, V.; Wellbrock, C.; Marais, R. Melanoma biology and new targeted therapy. Nature 2007, 445, 851–857.
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850.
- Davis, E.J.; Johnson, D.B.; Sosman, J.A.; Chandra, S. Melanoma: What do all the mutations mean? Cancer 2018, 124, 3490–3499.
- Davies, H.; Bignell, G.R.; Cox, C.; Stephens, P.; Edkins, S.; Clegg, S.; Teague, J.; Woffendin, H.; Garnett, M.J.; Bottomley, W.; et al. Mutations of the BRAF gene in human cancer. Nature 2002, 417, 949–954.
- Wan, P.T.; Garnett, M.J.; Roe, S.M.; Lee, S.; Niculescu-Duvaz, D.; Good, V.M.; Jones, C.M.; Marshall, C.J.; Springer, C.J.; Barford, D.; et al. Mechanism of activation of the RAF-ERK signaling pathway by oncogenic mutations of B-RAF. Cell 2004, 116, 855–867.
- Richtig, G.; Hoeller, C.; Kashofer, K.; Aigelsreiter, A.; Heinemann, A.; Kwong, L.N.; Pichler, M.; Richtig, E. Beyond the BRAFV600E hotspot: Biology and clinical implications of rare BRAF gene mutations in melanoma patients. Br. J. Dermatol. 2017, 177, 936–944.
- Curtin, J.A.; Fridlyand, J.; Kageshita, T.; Patel, H.N.; Busam, K.J.; Kutzner, H.; Cho, K.H.; Aiba, S.; Bröcker, E.B.; LeBoit, P.E.; et al. Distinct sets of genetic alterations in melanoma. N. Engl. J. Med. 2005, 353, 2135–2147.
- Jakob, J.A.; Bassett, R.L., Jr.; Ng, C.S.; Curry, J.L.; Joseph, R.W.; Alvarado, G.C.; Rohlfs, M.L.; Richard, J.; Gershenwald, J.E.; Kim, K.B.; et al. NRAS mutation status is an independent prognostic factor in metastatic melanoma. Cancer 2012, 118, 4014–4023.
- Giehl, K. Oncogenic Ras in tumour progression and metastasis. Biol. Chem. 2005, 386, 193–205.
- Fedorenko, I.V.; Gibney, G.T.; Smalley, K.S. NRAS mutant melanoma: Biological behavior and future strategies for therapeutic management. Oncogene 2013, 32, 3009–3018.
- Maertens, O.; Johnson, B.; Hollstein, P.; Frederick, D.T.; Cooper, Z.A.; Messiaen, L.; Bronson, R.T.; McMahon, M.; Granter, S.; Flaherty, K.; et al. Elucidating distinct roles for NF1 in melanomagenesis. Cancer Discov. 2013, 3, 338–349.
- Whittaker, S.R.; Theurillat, J.P.; Van Allen, E.; Wagle, N.; Hsiao, J.; Cowley, G.S.; Schadendorf, D.; Root, D.E.; Garraway, L.A. A genome-scale RNA interference screen implicates NF1 loss in resistance to RAF inhibition. Cancer Discov. 2013, 3, 350–362.
- Gibney, G.T.; Smalley, K.S. An unholy alliance: Cooperation between BRAF and NF1 in melanoma development and BRAF inhibitor resistance. Cancer Discov. 2013, 3, 260–263.
- Nissan, M.H.; Pratilas, C.A.; Jones, A.M.; Ramirez, R.; Won, H.; Liu, C.; Tiwari, S.; Kong, L.; Hanrahan, A.J.; Yao, Z.; et al. Loss of NF1 in cutaneous melanoma is associated with RAS activation and MEK dependence. Cancer Res. 2014, 74, 2340–2350.
- Thielmann, C.M.; Chorti, E.; Matull, J.; Murali, R.; Zaremba, A.; Lodde, G.; Jansen, P.; Richter, L.; Kretz, J.; Möller, I.; et al. NF1-mutated melanomas reveal distinct clinical characteristics depending on tumour origin and respond favourably to immune checkpoint inhibitors. Eur. J. Cancer 2021, 159, 113–124.
- Beadling, C.; Jacobson-Dunlop, E.; Hodi, F.S.; Le, C.; Warrick, A.; Patterson, J.; Town, A.; Harlow, A.; Cruz, F., 3rd; Azar, S.; et al. KIT gene mutations and copy number in melanoma subtypes. Clin. Cancer Res. 2008, 14, 6821–6828.
- Handolias, D.; Salemi, R.; Murray, W.; Tan, A.; Liu, W.; Viros, A.; Dobrovic, A.; Kelly, J.; McArthur, G.A. Mutations in KIT occur at low frequency in melanomas arising from anatomical sites associated with chronic and intermittent sun exposure. Pigment. Cell Melanoma Res. 2010, 23, 210–215.
- Pham, D.D.M.; Guhan, S.; Tsao, H. KIT and Melanoma: Biological Insights and Clinical Implications. Yonsei Med. J. 2020, 61, 562–571.
- Loras, A.; Gil-Barrachina, M.; Marqués-Torrejón, M.Á.; Perez-Pastor, G.; Martinez-Cadenas, C. UV-Induced Somatic Mutations Driving Clonal Evolution in Healthy Skin, Nevus, and Cutaneous Melanoma. Life 2022, 12, 1339.
- Wagstaff, W.; Mwamba, R.N.; Grullon, K.; Armstrong, M.; Zhao, P.; Hendren-Santiago, B.; Qin, K.H.; Li, A.J.; Hu, D.A.; Youssef, A.; et al. Melanoma: Molecular genetics, metastasis, targeted therapies, immunotherapies, and therapeutic resistance. Genes Dis. 2022, 9, 1608–1623.
- Tadijan, A.; Precazzini, F.; Hanžić, N.; Radić, M.; Gavioli, N.; Vlašić, I.; Ozretić, P.; Pinto, L.; Škreblin, L.; Barban, G.; et al. Altered Expression of Shorter p53 Family Isoforms Can Impact Melanoma Aggressiveness. Cancers 2021, 13, 5231.
- Loureiro, J.B.; Raimundo, L.; Calheiros, J.; Carvalho, C.; Barcherini, V.; Lima, N.R.; Gomes, C.; Almeida, M.I.; Alves, M.G.; Costa, J.L.; et al. Targeting p53 for Melanoma Treatment: Counteracting Tumour Proliferation, Dissemination and Therapeutic Resistance. Cancers 2021, 13, 1648.
- Wu, H.; Goel, V.; Haluska, F.G. PTEN signaling pathways in melanoma. Oncogene 2003, 22, 3113–3122.
- Tsao, H.; Goel, V.; Wu, H.; Yang, G.; Haluska, F.G. Genetic interaction between NRAS and BRAF mutations and PTEN/MMAC1 inactivation in melanoma. J. Investig. Dermatol. 2004, 122, 337–341.
- Stahl, J.M.; Cheung, M.; Sharma, A.; Trivedi, N.R.; Shanmugam, S.; Robertson, G.P. Loss of PTEN promotes tumor development in malignant melanoma. Cancer Res. 2003, 63, 2881–2890.
- Nogueira, C.; Kim, K.H.; Sung, H.; Paraiso, K.H.; Dannenberg, J.H.; Bosenberg, M.; Chin, L.; Kim, M. Cooperative interactions of PTEN deficiency and RAS activation in melanoma metastasis. Oncogene 2010, 29, 6222–6232.
- Shi, H.; Hugo, W.; Kong, X.; Hong, A.; Koya, R.C.; Moriceau, G.; Chodon, T.; Guo, R.; Johnson, D.B.; Dahlman, K.B.; et al. Acquired resistance and clonal evolution in melanoma during BRAF inhibitor therapy. Cancer Discov. 2014, 4, 80–93.
- McCubrey, J.A.; Steelman, L.S.; Kempf, C.R.; Chappell, W.H.; Abrams, S.L.; Stivala, F.; Malaponte, G.; Nicoletti, F.; Libra, M.; Bäsecke, J.; et al. Therapeutic resistance resulting from mutations in Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR signaling pathways. J. Cell. Physiol. 2011, 226, 2762–2781.
- Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Kempf, R.C.; Long, J.; Laidler, P.; Mijatovic, S.; Maksimovic-Ivanic, D.; Stivala, F.; Mazzarino, M.C.; et al. Roles of the Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR pathways in controlling growth and sensitivity to therapy-implications for cancer and aging. Aging 2011, 3, 192–1222.
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Franklin, R.A.; Montalto, G.; Cervello, M.; Libra, M.; Candido, S.; Malaponte, G.; et al. Ras/Raf/MEK/ERK and PI3K/PTEN/Akt/mTOR cascade inhibitors: How mutations can result in therapy resistance and how to overcome resistance. Oncotarget 2012, 3, 1068–1111.
- Iura, K.; Kohashi, K.; Hotokebuchi, Y.; Ishii, T.; Maekawa, A.; Yamada, Y.; Yamamoto, H.; Iwamoto, Y.; Oda, Y. Cancer-testis antigens PRAME and NY-ESO-1 correlate with tumour grade and poor prognosis in myxoid liposarcoma. J. Pathol. Clin. Res. 2015, 1, 144–159.
- Ghafouri-Fard, S.; Gholipour, M.; Taheri, M. MicroRNA Signature in Melanoma: Biomarkers and Therapeutic Targets. Front. Oncol. 2021, 11, 608987.
- Poniewierska-Baran, A.; Słuczanowska-Głąbowska, S.; Małkowska, P.; Sierawska, O.; Zadroga, Ł.; Pawlik, A.; Niedźwiedzka-Rystwej, P. Role of miRNA in Melanoma Development and Progression. Int. J. Mol. Sci. 2023, 24, 201.
- Bennett, P.E.; Bemis, L.; Norris, D.A.; Shellman, Y.G. miR in melanoma development: miRNAs and acquired hallmarks of cancer in melanoma. Physiol. Genom. 2013, 45, 1049–1059.
- Díaz-Martínez, M.; Benito-Jardón, L.; Alonso, L.; Koetz-Ploch, L.; Hernando, E.; Teixidó, J. miR-204-5p and miR-211-5p Contribute to BRAF Inhibitor Resistance in Melanoma. Cancer Res. 2018, 78, 1017–1030.
- Motti, M.L.; Minopoli, M.; Di Carluccio, G.; Ascierto, P.A.; Carriero, M.V. MicroRNAs as Key Players in Melanoma Cell Resistance to MAPK and Immune Checkpoint Inhibitors. Int. J. Mol. Sci. 2020, 21, 4544.
- Zhang, J.; Zhao, H.; Gao, Y.; Zhang, W. Secretory miRNAs as novel cancer biomarkers. Biochim. Biophys. Acta 2012, 1826, 32–43.
- Li, J.; Liu, Y.; Wang, C.; Deng, T.; Liang, H.; Wang, Y.; Huang, D.; Fan, Q.; Wang, X.; Ning, T.; et al. Serum miRNA expression profile as a prognostic biomarker of stage II/III colorectal adenocarcinoma. Sci. Rep. 2015, 5, 12921.
- Hanniford, D.; Zhong, J.; Koetz, L.; Gaziel-Sovran, A.; Lackaye, D.J.; Shang, S.; Pavlick, A.; Shapiro, R.; Berman, R.; Darvishian, F.; et al. A miRNA-Based Signature Detected in Primary Melanoma Tissue Predicts Development of Brain Metastasis. Clin. Cancer Res. 2015, 21, 4903–4912.
- Stark, M.S.; Klein, K.; Weide, B.; Haydu, L.E.; Pflugfelder, A.; Tang, Y.H.; Palmer, J.M.; Whiteman, D.C.; Scolyer, R.A.; Mann, G.J.; et al. The Prognostic and Predictive Value of Melanoma-related MicroRNAs Using Tissue and Serum: A MicroRNA Expression Analysis. EBioMedicine 2015, 2, 671–680.
- Antonova, E.; Hambikova, A.; Shcherbakov, D.; Sukhov, V.; Vysochanskaya, S.; Fadeeva, I.; Gorshenin, D.; Sidorova, E.; Kashutina, M.; Zhdanova, A.; et al. Determination of Common microRNA Biomarker Candidates in Stage IV Melanoma Patients and a Human Melanoma Cell Line: A Potential Anti-Melanoma Agent Screening Model. Int. J. Mol. Sci. 2023, 24, 9160.
- Vitiello, M.; D’Aurizio, R.; Poliseno, L. Biological role of miR-204 and miR-211 in melanoma. Oncoscience 2018, 5, 248–251.
- Varrone, F.; Caputo, E. The miRNAs Role in Melanoma and in Its Resistance to Therapy. Int. J. Mol. Sci. 2020, 21, 878.
- Qian, L.-Y.; Li, P.; He, Q.-Y.; Luo, C.-Q. Circulating miR-221 Expression Level and Prognosis of Cutaneous Malignant Melanoma. Experiment 2014, 20, 2472–2477.
- Rigg, E.; Wang, J.; Xue, Z.; Lunavat, T.R.; Liu, G.; Hoang, T.; Parajuli, H.; Han, M.; Bjerkvig, R.; Nazarov, P.V.; et al. Inhibition of extracellular vesicle-derived miR-146a-5p decreases progression of melanoma brain metastasis via Notch pathway dysregulation in astrocytes. J. Extracell. Vesicles 2023, 12, e12363.
- Bell, R.E.; Khaled, M.; Netanely, D.; Schubert, S.; Golan, T.; Buxbaum, A.; Janas, M.M.; Postolsky, B.; Goldberg, M.S.; Shamir, R.; et al. Transcription factor/microRNA axis blocks melanoma invasion program by miR-211 targeting NUAK1. J. Investig. Dermatol. 2014, 134, 441–451.
- Luan, W.; Li, R.; Liu, L.; Ni, X.; Shi, Y.; Xia, Y.; Wang, J.; Lu, F.; Xu, B. Long non-coding RNA HOTAIR acts as a competing endogenous RNA to promote malignant melanoma progression by sponging miR-152-3p. Oncotarget 2017, 8, 85401–85414.
- Rang, Z.; Yang, G.; Wang, Y.W.; Cui, F. miR-542-3p suppresses invasion and metastasis by targeting the proto-oncogene serine/threonine protein kinase, PIM1, in melanoma. Biochem. Biophys. Res. Commun. 2016, 474, 315–320.
- Nabipoorashrafi, S.A.; Shomali, N.; Sadat-Hatamnezhad, L.; Mahami-Oskouei, M.; Mahmoudi, J.; Shotorbani, B.S.; Akbari, M.; Xu, H. miR-143 acts as an inhibitor of migration and proliferation as well as an inducer of apoptosis in melanoma cancer cells in vitro. IUBMB Life 2020, 72, 2034–2044.
- Liu, Y.; Ruan, H.; Lu, F.; Peng, H.; Luan, W. miR-224-5p acts as a tumour suppressor and reverses the resistance to BRAF Inhibitor In melanoma through directly targeting PAK4 to block the MAPK pathway. Pathol. Res. Pract. 2023, 249, 154772.
- Sun, X.; Li, J.; Sun, Y.; Zhang, Y.; Dong, L.; Shen, C.; Yang, L.; Yang, M.; Li, Y.; Shen, G.; et al. miR-7 reverses the resistance to BRAFi in melanoma by targeting EGFR/IGF-1R/CRAF and inhibiting the MAPK and PI3K/AKT signaling pathways. Oncotarget 2016, 7, 53558–53570.
- Caporali, S.; Amaro, A.; Levati, L.; Alvino, E.; Lacal, P.M.; Mastroeni, S.; Ruffini, F.; Bonmassar, L.; Antonini Cappellini, G.C.; Felli, N.; et al. miR-126-3p down-regulation contributes to dabrafenib acquired resistance in melanoma by up-regulating ADAM9 and VEGF-A. J. Exp. Clin. Cancer Res. 2019, 38, 272.
- Diaz-Martinez, M.; Benito-Jardon, L.; Teixido, J. New insights in melanoma resistance to BRAF inhibitors: A role for microRNAs. Oncotarget 2018, 9, 35374–35375.
- Zheng, Y.; Sun, Y.; Liu, Y.; Zhang, X.; Li, F.; Li, L.; Wang, J. The miR-31-SOX10 axis regulates tumor growth and chemotherapy resistance of melanoma via PI3K/AKT pathway. Biochem. Biophys. Res. Commun. 2018, 503, 2451–2458.
- Surman, M.; Jankowska, U.; Wilczak, M.; Przybyło, M. Similarities and Differences in the Protein Composition of Cutaneous Melanoma Cells and Their Exosomes Identified by Mass Spectrometry. Cancers 2023, 15, 1097.
- Whiteside, T.L. Immunosuppressive functions of melanoma cell-derived exosomes in plasma of melanoma patients. Front. Cell Dev. Biol. 2023, 10, 1080925.
- Zhu, H.; Zhang, P.; Shi, J.; Kou, D.; Bai, X. Exosome-delivered circRPS5 inhibits the progression of melanoma via regulating the miR-151a/NPTX1 axis. PLoS ONE 2023, 18, e0287347.
- Shen, Y.; Li, M.; Liao, L.; Gao, S.; Wang, Y. Plasma exosome-derived connexin43 as a promising biomarker for melanoma patients. BMC Cancer 2023, 23, 242.
- Boussadia, Z.; Lamberti, J.; Mattei, F.; Pizzi, E.; Puglisi, R.; Zanetti, C.; Pasquini, L.; Fratini, F.; Fantozzi, L.; Felicetti, F.; et al. Acidic microenvironment plays a key role in human melanoma progression through a sustained exosome mediated transfer of clinically relevant metastatic molecules. J. Exp. Clin. Cancer Res. 2018, 37, 245.
- Surman, M.; Kędracka-Krok, S.; Hoja-Łukowicz, D.; Jankowska, U.; Drożdż, A.; Stępień, E.Ł.; Przybyło, M. Mass Spectrometry-Based Proteomic Characterization of Cutaneous Melanoma Ectosomes Reveals the Presence of Cancer-Related Molecules. Int. J. Mol. Sci. 2020, 21, 2934.
- Lattmann, E.; Levesque, M.P. The Role of Extracellular Vesicles in Melanoma Progression. Cancers 2022, 14, 3086.
- Strnadová, K.; Pfeiferová, L.; Přikryl, P.; Dvořánková, B.; Vlčák, E.; Frýdlová, J.; Vokurka, M.; Novotný, J.; Šáchová, J.; Hradilová, M.; et al. Exosomes produced by melanoma cells significantly influence the biological properties of normal and cancer-associated fibroblasts. Histochem. Cell Biol. 2022, 157, 153–172.
- Xiao, D.; Barry, S.; Kmetz, D.; Egger, M.; Pan, J.; Rai, S.N.; Qu, J.; McMasters, K.M.; Hao, H. Melanoma cell-derived exosomes promote epithelial-mesenchymal transition in primary melanocytes through paracrine/autocrine signaling in the tumor microenvironment. Cancer Lett. 2016, 376, 318–327.
- Biagioni, A.; Laurenzana, A.; Menicacci, B.; Peppicelli, S.; Andreucci, E.; Bianchini, F.; Guasti, D.; Paoli, P.; Serratì, S.; Mocali, A.; et al. uPAR-expressing melanoma exosomes promote angiogenesis by VE-Cadherin, EGFR and uPAR overexpression and rise of ERK1,2 signaling in endothelial cells. Cell Mol. Life Sci. 2021, 78, 3057–3072.
- García-Silva, S.; Benito-Martín, A.; Nogués, L.; Hernández-Barranco, A.; Mazariegos, M.S.; Santos, V.; Hergueta-Redondo, M.; Ximénez-Embún, P.; Kataru, R.P.; Lopez, A.A.; et al. Melanoma-derived small extracellular vesicles induce lymphangiogenesis and metastasis through an NGFR-dependent mechanism. Nat. Cancer 2021, 2, 1387–1405.
- Wang, P.; Wu, Y.; Chen, W.; Zhang, M.; Qin, J. Malignant melanoma-derived exosomes induce endothelial damage and glial activation on a human BBB chip model. Biosensors 2022, 12, 89.
- Ye, Q.; Li, Z.; Li, Y.; Li, Y.; Zhang, Y.; Gui, R.; Cui, Y.; Zhang, Q.; Qian, L.; Xiong, Y.; et al. Exosome-Derived microRNA: Implications in Melanoma Progression, Diagnosis and Treatment. Cancers 2023, 15, 80.
- Tan, Y.; Tang, F.; Li, J.; Yu, H.; Wu, M.; Wu, Y.; Zeng, H.; Hou, K.; Zhang, Q. Tumor-derived exosomes: The emerging orchestrators in melanoma. Biomed. Pharmacother. 2022, 149, 112832.
- Gerloff, D.; Lützkendorf, J.; Moritz, R.K.C.; Wersig, T.; Mäder, K.; Müller, L.P.; Sunderkötter, C. Melanoma-Derived Exosomal miR-125b-5p Educates Tumor Associated Macrophages (TAMs) by Targeting Lysosomal Acid Lipase A (LIPA). Cancers 2020, 12, 464.
- Li, X.; Liu, Y.; Yang, L.; Jiang, Y.; Qian, Q. TIM-3 shuttled by MV3 cells-secreted exosomes inhibits CD4+ T cell immune function and induces macrophage M2 polarization to promote the growth and metastasis of melanoma cells. Transl. Oncol. 2022, 18, 101334.
- Zhou, Q.; Fang, T.; Wei, S.; Chai, S.; Yang, H.; Tao, M.; Cao, Y. Macrophages in melanoma: A double-edged sword and targeted therapy strategies (Review). Exp. Ther. Med. 2022, 24, 640.
- Gu, Y.; Du, Y.; Jiang, L.; Tang, X.; Li, A.; Zhao, Y.; Lang, Y.; Liu, X.; Liu, J. αvβ3 integrin-specific exosomes engineered with cyclopeptide for targeted delivery of triptolide against malignant melanoma. J. Nanobiotechnol. 2022, 20, 384.
- Gao, H.; Lao, Y.; Zhang, J.; Ding, B. Dendritic Cell-Derived Exosomes Driven Drug Co-Delivery Biomimetic Nanosystem for Effective Combination of Malignant Melanoma Immunotherapy and Gene Therapy. Drug Des. Devel. Ther. 2023, 17, 2087–2106.
- Naeem, P.; Baumgartner, A.; Ghaderi, N.; Sefat, F.; Alhawamdeh, M.; Heidari, S.; Shahzad, F.; Swaminathan, K.; Akhbari, P.; Isreb, M.; et al. Anticarcinogenic impact of extracellular vesicles (exosomes) from cord blood stem cells in malignant melanoma: A potential biological treatment. J. Cell. Mol. Med. 2023, 27, 222–231.
More
Information
Subjects:
Dermatology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
497
Revisions:
2 times
(View History)
Update Date:
14 Nov 2023
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No