Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Alberto Pietrantoni | -- | 5116 | 2023-11-06 23:50:10 | | | |
| 2 | Lindsay Dong | Meta information modification | 5116 | 2023-11-08 02:21:51 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Serioli, S.; Agostini, L.; Pietrantoni, A.; Valeri, F.; Costanza, F.; Chiloiro, S.; Buffoli, B.; Piazza, A.; Poliani, P.L.; Peris-Celda, M.; et al. Aggressive PitNETs and Potential Target Therapies. Encyclopedia. Available online: https://encyclopedia.pub/entry/51219 (accessed on 25 June 2026).
Serioli S, Agostini L, Pietrantoni A, Valeri F, Costanza F, Chiloiro S, et al. Aggressive PitNETs and Potential Target Therapies. Encyclopedia. Available at: https://encyclopedia.pub/entry/51219. Accessed June 25, 2026.
Serioli, Simona, Ludovico Agostini, Alberto Pietrantoni, Federico Valeri, Flavia Costanza, Sabrina Chiloiro, Barbara Buffoli, Amedeo Piazza, Pietro Luigi Poliani, Maria Peris-Celda, et al. "Aggressive PitNETs and Potential Target Therapies" Encyclopedia, https://encyclopedia.pub/entry/51219 (accessed June 25, 2026).
Serioli, S., Agostini, L., Pietrantoni, A., Valeri, F., Costanza, F., Chiloiro, S., Buffoli, B., Piazza, A., Poliani, P.L., Peris-Celda, M., Iavarone, F., Gaudino, S., Gessi, M., Schinzari, G., Mattogno, P.P., Giampietro, A., De Marinis, L., Pontecorvi, A., Fontanella, M.M., ...Doglietto, F. (2023, November 06). Aggressive PitNETs and Potential Target Therapies. In Encyclopedia. https://encyclopedia.pub/entry/51219
Serioli, Simona, et al. "Aggressive PitNETs and Potential Target Therapies." Encyclopedia. Web. 06 November, 2023.
Copy Citation
Pituitary adenomas, namely pituitary neuroendocrine tumors (PitNETs), are a heterogeneous group of neoplasms deriving from the neuroendocrine cell of the adeno-pituitary and represent 15% of all intracranial tumors. Evidence has suggested the importance of epigenetic modifications in influencing PitNET pathophysiology and clinical behavior. Indeed, most pituitary tumors are sporadic without any specific genetic driver mutations. Furthermore, a better understanding of epigenetic modifications in PitNETs is paramount to developing novel therapeutic strategies.
aggressive PitNETs
gene expression
invasive PitNETs
molecular pathway
precision medicine
target therapy
1. Introduction
Pituitary adenomas, namely pituitary neuroendocrine tumors (PitNETs), are a heterogeneous group of neoplasms deriving from the neuroendocrine cell of the adeno-pituitary and represent 15% of all intracranial tumors [1][2]. Although most PitNETs are benign and respond to conventional treatment with an excellent outcome, up to 40% of cases display aggressive behavior, with an unpredictable and rapid evolution affecting the patient’s overall quality of life [3][4]. For this subgroup, the term “aggressive pituitary adenoma” was used; in 2017, the acronym PitNETs was suggested for all pituitary adenomas, highlighting the need to determine the benign or aggressive nature of each one of them and underscoring the heterogeneity of the biological and clinical behavior of these lesions [5]. According to the European Society of Endocrinology (ESE), aggressive PitNETs are characterized by propagation to extrasellar regions (traditionally defined in the parasellar area according to Knosp’s modified classification), rapid tumor growth, high hormonal serum levels, and resistance to standard treatments (medical therapy, surgery, or radiotherapy), determining high morbidity and mortality [6]. The prognosis of these tumors strictly depends on their biology, and the search for reliable clinical and pathological markers as predictors of tumor behavior continues.
The need to identify efficient treatments for managing aggressive PitNETs has increasingly emerged. Temozolomide was the first chemotherapy drug to be used as monotherapy and combined with radiotherapy or 5-fluorouracil, with variable results depending on the type of PitNET and previous treatments [7][8][9][10]. Recently, gene alterations, anomalous expression of specific molecular pathways, and regulatory molecules have become objects of interest in discovering new target therapies thanks to the technological improvements in molecular biology, genetics, and bioinformatics. Different studies have brought new knowledge for future treatment options, laying the foundations for precision or personalized medicine. Emerging therapies such as immune checkpoint inhibitors (ICIs), vascular endothelial growth factor (VEGF) receptor therapy, tyrosine kinase inhibitors (TKIs), mammalian target of rapamycin (mTOR) inhibitors, and peptide receptor radionuclide therapy (PRRT) have been tested on a limited number of patients, with variable results [6][11][12][13].
2. Aggressive PitNETs and Potential Target Therapies
2.1. Tumor Microenvironment
The tumor microenvironment (TME) is characterized by the interplay between the host and tumor cells. TME comprises non-tumor cells, vascularization, extracellular matrix, cytokines, and enzymes (Figure 1) [11][14]. The immune infiltration is eterogenous among the different subtypes of PitNET. As the TME modulates tumorigenic mechanisms such as cell proliferation, invasiveness, and angiogenesis, it may help explain PitNETs’ heterogeneous behavior [11][15]. The TME study in PitNETs has been expanding over the last few years, leading to the identification of new biomarkers for prognostic stratification and the development of novel therapies.
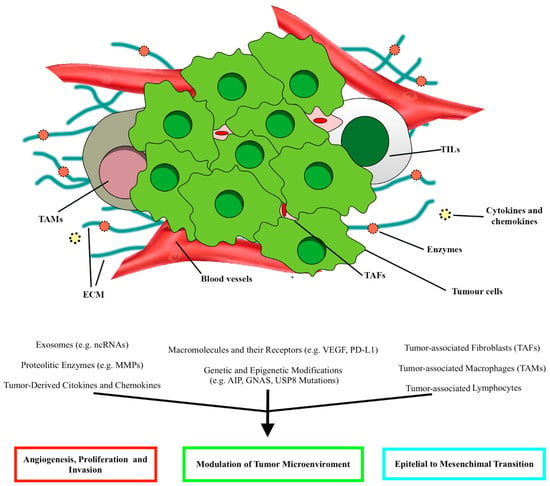
Figure 1. Schematic illustration of the main elements constituting the tumor microenvironment (TME) in PitNETs and the relative effects, according to Marques and Korbonits [16]. ECM, extracellular matrix; TAFs, tumor-associated fibroblasts; TALs, tumor-associated lymphocytes; TAMs, tumor-associated macrophages.
2.1.1. Macrophages
Macrophages, distinguished in M1 and M2 subpopulations, are immune cells infiltrating mainly sparsely granulated somatotropinomas, null-cell adenomas, and gonadotropinomas.
While M1-macrophages are characteristic of normal pituitary, the predominant population in PitNETs are M2-macrophages, characterized by pro-tumorigenic effects and associated with worse outcomes [15]. They showed a pro-angiogenic role in estrogen-induced rat prolactinomas and a tumorigenic function in diethylstilbestrol-induced rat prolactinomas [14][16].
In somatotropinomas, higher amounts of tumor-infiltrating CD68+ macrophages, widely present in sparsely granulated forms, correlated positively with tumor volume and higher expression of Ki-67 and MMPs, cavernous sinus invasion [17]. High amounts of macrophages were also revealed in AIP mutation-positive somatotrophinomas [16].
2.1.2. Lymphocytes
The role of lymphocytic infiltrate in pituitary tumors has been described, including CD3+, CD45+, cytotoxic CD8+ T cells, and CD4+ T helper, in particular in functioning pituitary tumors, and, to a lesser extent, also B cells, neutrophils, and mast cells [18]. Some recent studies reported significant infiltrates of natural killer (NK) cells and FOXP3+ T cells in the microenvironment of PIT1-positive pituitary tumors [19].
Different lymphocyte subtypes may have potentially anti- or pro-tumor effects. In pituitary tumors with a Ki-67 ≥ 3%, lower CD8:CD4 and CD8:FOXP3 T cell ratios and higher levels of macrophages, T helper, FOXP3+, and B cells were reported [16]. Invasiveness has been associated with a higher CD8+ T cell count and a higher FOXP3:CD8 cell ratio, especially in non-functioning pituitary adenomas (NF-PitNETs) [20].
2.1.3. Stromal Cells
Stromal cells include tumor-associated fibroblasts, myoepithelial cells, and pericytes and can enhance tumor cell proliferation and invasiveness. Three subtypes of tumor-associated fibroblasts have been described: antigen-presenting fibroblasts, inflammatory fibroblasts, and myofibroblasts [19]. Tumor-associated fibroblasts seem to play a crucial role in the progression of PIT1-positive and corticotropic tumors. In invasive PitNETs, fibroblasts are characterized by higher expression of both α-smooth muscle actin (α-SMA) and VEGF, positively influencing the proliferation of GH3 pituitary tumor cells [21].
A potential association between IL-6 and CCL2, secreted by tumor-associated fibroblasts, and high Ki-67 levels was found, speculating how tumor proliferation in PitNETs may be influenced by fibroblasts [16][22].
2.1.4. Folliculo-Stellate Cells
Follicle-stellate cells are identified in most PitNETs, and through their ability to release growth factors and cytokines, they maintain a balance between the different cell types and perform immune functions [23]. Recently, an association between increased S100B+ folliculo-stellate cells and lower tumor proliferation was positively related to the expression of estrogen receptor-α and FSH in gonadotropinomas [24]. Higher growth hormone levels were secreted in another study on somatotropinomas with scattered folliculo-stellate cells [25].
2.1.5. Cytokines, Chemokines, and Growth Factors
Cytokines, growth factors, and chemokines, produced by tumor cells and those surrounding the tumor, can trigger cellular defense mechanisms and regulate tumor progression. Cytokines and growth factors, such as CXCL12, CCL5, CCL17, IL-8, IL-6, IL-1, IL-2, IL-17, tumor necrosis factor-α, and vascular endothelial growth factor (VEGF), can affect tumorigenic mechanisms in pituitary neoplasms [16]. Chemokines, such as CXCL8 (or interleukin (IL)-8), CCL2, CCL3, and CCL4, are secreted by pituitary tumors and non-tumor cells, such as macrophages, lymphocytes, or fibroblasts, and modulate the microenvironment composition [16][20].
The increased expression of immune checkpoints, such as the programmed death ligand-1 (PD-L1) and the cytotoxic T-lymphocyte-associated protein 4 (CTLA-4) ligands CD80 and CD86, has been associated with aggressiveness in proliferative pituitary tumors [16].
The pituitary tumors characterized by elevated expression of PD-L1 presented prevalent immune infiltrates of CD4+, CD8+, and FOXP3+ T cells, highlighting the ability of pituitary tumor-infiltrating immune cells to modulate the expression of immune checkpoint regulators [16]. Moreover, increased expression of the aforementioned CD163 was positively associated with higher expression of PD-L1, PD-L2, and lymphocyte activating 3 (LAG3) [16][26]. The increased presence of macrophages, CD4+, CD8+, and CD45+ T cells was related to the expression of V-domain immunoglobulin suppressor of T cell activation (VISTA) and LAG3 [26].
2.1.6. The Role of Immunotherapy in Pituitary Tumors: Response and Outcomes
The efforts of the last decades in understanding TME paved the way for new therapeutic options, such as immune checkpoint inhibitors (ICIs). The growing association between the aggressive nature of PitNETs subgroups unresponsive to conventional treatments and the expression of immune checkpoints has increasingly determined the use of drugs targeting CTLA-4 (ipilimumab) or PD-1/PD-L1 (nivolumab or pembrolizumab) [27]. Only a few cases of pituitary tumors treated with ICIs have been reported. Using these drugs in single (such as pembrolizumab) or combined formulations (ipilimumab and nivolumab) allowed a partial radiological response to be obtained in five cases of pituitary carcinoma [6][28][29]. In comparison, in two other cases, the disease remained stable [6][28][29]. Maintenance therapy with nivolumab was used in those cases of pituitary carcinoma with an initial response to ipilimumab and nivolumab, showing a radiological response. In the case of disease progression, the carcinoma was retreated with ipilimumab and nivolumab without success, while in the others, a radiological response was documented [29]. Progression-free survival (4 to 42 months) was found in three carcinomas treated with pembrolizumab [29].
The latest efforts aimed at determining which patients could benefit from immunotherapy or how to improve/enhance the therapeutic response for each patient, such as the combined effect of checkpoint inhibitors with peptide receptor radionuclide therapy [30], hypothesize that radiation may influence the TME, further sensitizing the pituitary tumor to the therapeutic response of immunotherapy.
Concerning the individual components of the TME, a tumor-infiltrating immune profile has been associated with poorer outcomes and recurrence in PitNETs [6]. Moreover, the identification of CD8+ T cells could predict the response to first-generation somatostatin analogs, regardless of tumor characteristics (hormone levels, size, and invasion) and patient age, as demonstrated by Chiloiro et al., where somatotropinomas with high levels of CD8+ and CD138+ lymphocytes responded to first-generation somatostatin analogs [31].
In addition, traditional therapies can influence TME. Somatostatin and its analogs are characterized not only by their anti-tumor action but also by the possibility of influencing the pituitary tumor microenvironment, affecting fibroblasts, and inhibiting the release of cytokines, growth factors, and VEGF from pituitary tumor cells [16][22][32][33]. The immunomodulation of dopamine with DAs, thanks to the presence of receptors on endothelial cells, macrophages, and lymphocytes, has also been described, highlighting the inhibitory effect of cytokine and growth factor secretion, blocking chemotaxis, and inducing apoptosis, especially during the management of prolactinomas and somatotropinomas [34][35][36].
2.2. PI3K/Akt/mTOR and RAS/MEK/ERK Pathways
The PI3K/Akt/mTOR and RAS/MEK/ERK pathways are fundamental for cell survival, proliferation, migration, regulation of protein, lipid, and nucleic acid metabolism, senescence, and autophagy. They are triggered by the bond between extracellular ligands and receptor-linked tyrosine kinases (RTK); this bond triggers a cascade of phosphorylation-type reactions that ultimately activate or deactivate various substrates, including transcription factors. This bond, for example, can activate the phosphatidyl-inositol-3-kinase (PI3K), which in turn stimulates the formation of mTOR Complex 1 (mTORC1) and 2 (mTORC2), two protein complexes through which mTOR can operate its effects. Growth factors binding their RTKs also trigger a protein of the Ras family (like H-Ras or K-Ras), which prompts a cascade of sequential phosphorylation-type events that activate RAF, then MEK1/2, then ERK1/2. These kinases interact with various enzymes and transcription factors and also converge on the mTOR pathway. The PI3K/Akt/mTOR and RAF/MEK/ERK pathways are involved in many human neoplasms [37][38][39].
EGFR signaling, which involves the mTOR pathway, is expressed in these tumors, and it is associated with tumor proliferation, invasive behavior, lower total resection, and epithelial-to-mesenchymal transition [40][41][42]; this latter is also mediated by the ADAM12 metalloprotease [42]. IGF-1 signaling also exerts a mitogenic effect through the PI3K/mTOR/Akt pathway [43][44].
Expressing platelet-derived growth factor (PDGF) and its receptor by folliculostellate cells stimulates pituitary cell proliferation through the PI3K/mTOR/Akt pathway [45].
Mutation of the proto-oncogene PI3KCA (which encodes a subunit of PI3K) was found in 2.3% to 12.1% of tumor series, while amplification of the same gene was documented in 21.2% to 28% of cases [46][47]. Mutations of PTEN do not seem to occur frequently [48], but it is downregulated in PitNETs [49]. In its phosphorylated form, Akt is more expressed in PitNETs than in normal pituitary tissue [48][50][51][52][53][54][55], especially in recurrent tumors [51].
The mTOR molecule was shown to be expressed in PitNET cells [56] alongside two mTOR-related proteins, RAPTOR (part of mTORC1) and RICTOR (part of mTORC2) [49], with RAPTOR expression being associated with CS invasion. Unlike normal pituitary tissue, downstream effectors of mTOR, like phospho-S6 protein and phospho-4EBP1 protein, are increased in PitNETs [54][57]. DEPTOR, a down-regulator of mTOR, is underexpressed in PitNETs [58].
The RAF/RAS/MEK/ERK pathway, and especially the ERK molecule, is also overexpressed in PitNETs [42][43][50][51][56][57][59][60][61][62] and was shown to be regulated by the PI3K/Akt/mTOR pathway [62]. ERK also mediates the signaling of different growth factors, like EGFR [40] and IGF-1 [44], in pituitary tissue. The transcript of BRAF is overexpressed in pituitary adenomas compared to normal pituitary [63], and the BRAFV600E mutation was found in 16.5% of corticotroph adenomas [64].
In summary, current evidence points to overexpression and hyperactivation of crucial molecules of the PI3K/Akt/mTOR and RAF/MEK/ERK pathways in PitNETs.
Rapamycin (also known as Sirolimus) and its analog Everolimus (also known as RAD001) inhibit mTOR directly by binding the FKBP12 protein, forming a complex that interacts with the mTOR molecule and prevents it from forming mTORC1 and mTORC2. These drugs can reduce the number of viable PitNET cells, their proliferation, and the phosphorylation of downstream mTOR effectors [53][55][65][66][67], lower prolactin secretion, decrease mTOR phosphorylation, enhance the radiotherapy response, and block IGF-I proliferative and anti-apoptotic effects [65][67]. The anti-proliferative effects of Everolimus are enhanced by the co-treatment with Pasireotide [67], and a similar effect was observed for Rapamycin with Octreotide [68].
Inhibitors of PI3K, like the pan-PI3K inhibitor NVP-BKM120 (Buparlisib) and the specific PI3K-alpha inhibitor NVP-BYL719 (Alpelisib), have shown a dose-dependent inhibition of cell viability of PitNETs and display a synergistic effect when combined with Everolimus [66]. Inhibition of PI3K with the chemical compound LY294002 reduces PitNET cell growth, increases the pro-apoptotic activity of Bcl2-associated death promoter, decreases the anti-apoptotic effect of IGF-1, and decreases phosphorylation of PI3K and Akt [44][54][69][70][71].
Dual PI3K-mTOR inhibitors like NVP-BEZ235 (Dactolisib) seem more effective than Everolimus in reducing the cell viability of PitNETs [72]. NVP-BEZ235 treatment decreases Akt and S6 phosphorylation and triggers apoptosis [72][73]. Another dual inhibitor, XL765 (Voxtalisib), enhances the effects of temozolomide against PitNET cells [73].
Gefitinib, an anti-EGFR tyrosine kinase inhibitor, reverses the epithelial-to-mesenchymal phenotype, decreases invasiveness, and reduces the proliferation of PitNETs [42]. In another study, Gefitinib resulted in tumor shrinkage and a reduction in peripheral hormone levels by around 30% in a mouse model. Gefitinib treatment in mice decreased ERK1/2 phosphorylation, followed by downregulation of tumor prolactin mRNA [40].
Among other lesser-known drugs, the Akt inhibitor MK-2206 was shown to reduce the phosphorylation of Akt. The HIV protease inhibitor Nelfinavir radiosensitizes PA cell lines in vitro, and the underlying mechanism seems to involve the mTOR pathway [74].
Corticocotroph PitNET cells harboring the BRAFV600E mutation undergo a more significant reduction in hormone secretion when treated with BRAF inhibitor Vemurafenib, compared to tumor cells with wild-type BRAF [64].
In summary, drugs targeting the PI3K/Akt/mTOR pathway seem capable of interfering with PitNET growth, survival, and hormone secretion and enhancing the effects of other therapeutic strategies like radiotherapy, somatostatin analogs, and temozolomide. Still, clinical studies are required to study their effectiveness further.
2.3. Receptors
2.3.1. Somatostatin and Dopamine Receptors
Somatostatin receptors (SSTR) and dopamine receptors (DRD) represent a staple in PitNETs therapy, and their agonists are commonly used in clinical practice, especially in PIT-1-positive tumors (GH and PRL-positive subtypes), except corticotrophs and gonadotrophs. In particular, in Cushing’s patients, SSTR5, SSTR2, and SSTR3 are predominantly expressed, representing three of the four target receptors of pasireotide [75][76].
A novel pathway has been described by Peverelli et al. that might explain SSTR2 anti-neoplastic activity and its role in PitNETs pathophysiology via the activation of the Rhoa/ROCK pathway and the consequent cofilin phosphorylation; this way, cofilin is unable to bind to actin, and SSTR2 activation inhibits cytoskeleton remodeling and cell migration [77]. The exact mechanism for the DRD2 isoform was described by Peverelli et al., who also correlated low cofilin phosphorylation with invasion in aggressive PitNETs [78]. New molecules have been proposed, such as BIM23120 (a selective SSTR2 agonist) and BIM53097 (a selective DRD2 agonist), showing promising results in reducing somatotroph cell migration and proliferation and inducing apoptosis.
SSTR5 is highly expressed in NF-, ACTH-, and GH-PitNETs and correlates with recurrence [76]. When analyzed in association with DRDs, it has been shown to form chimeric receptors, particularly with DRD2 [79][80], and the chimeric receptor SSTR5/DRD2 has been linked with inferior dimension and grade [80]. Its targeting via BIM23A760 (a selective agonist) reduced GH and PRL production in vitro; in GH tumors, it reduced cell viability and proliferation and stimulated apoptosis [79].
Other SSTRs were less investigated, given their lower representation. The data above have revived interest in these receptor subtypes, and there is evidence that SSTR1 might represent an indicator for response to medical treatment and a prognostic factor: its overexpression has been described in recurrent ACTH-secreting tumors [76], and low levels are linked with remission after first surgery [81].
As for SSTR3, Lee et al. suggested its anti-neoplastic role is due to its implication in the MAPK pathway: SSTR3 activated tyr-phosphatases, ultimately inhibiting MAPK and consequently activating p53 and the caspases, promoting apoptosis [82]. SSTR3 is highly expressed in SF-1 positive PitNETs, even recurring ones [82], while it is reduced in recurring NF PitNETs [76].
2.3.2. Peptide Receptor Radionuclide Therapy
Peptide receptor radionuclide therapy (PRRT) is an innovative therapeutic option traditionally used as a second-line treatment for advanced (metastatic or inoperable) neuroendocrine tumors (NETs), characterized by the expression of the SSTRs, especially the SSTR2 [83][84]. The applicability of such treatment is closely related to the expression of somatostatin receptors, which can be evaluated through functional imaging such as 68Ga-DOTA peptide PET/CT, octreoscan, 111In-octreotide-scintigraphy, or 99m-EDDA-HYNIC-tyr3-octreotide scintigraphy [10][85][86][87][88][89][90][91][92][93][94][95][96].
2.3.3. Transforming Growth Factor Receptor
Transforming growth factor beta (TGF-β) signaling is related to numerous biological processes involved with cell proliferation, differentiation, apoptosis, and EMT. Its action is mediated by TGF-β receptor complexes (TGF-β RI and RII) that phosphorylate Smad2 and Smad3, which in turn form trimers with Smad4 and translocate to the nucleus, regulating the expression of genes controlling the cell cycle [97]. TGF-β has been investigated in different types of cancer, and it can act as either a suppressor or an inhibitor of tumor development, depending on the tumor and stage [98]. Given its intricate relationship with tumorigenesis, new data are being implemented to uncover its role in PitNETs.
Some authors have described that TGF-β is elevated in aggressive PRL-secreting tumors, both locally [99] and in the serum (and serum TGF-β is directly correlated with tumor dimensions and aggressivity) [100]. Jiang et al. correlated TGF-β levels with microvascular density in the neoplastic tissue [99].
On the contrary, when analyzing the TGF-β pathway, Ying-Hao et al. reported an inverse association between TGF-β RII expression and invasiveness, thus documenting it as a tumor suppressor. No significant difference was highlighted for TGF-β RI [101].
2.3.4. Fibroblast Growth Factor Receptors
Fibroblast growth factor receptors (FGFR) are tyrosin-kinase receptors, and their activation mediates cell proliferation, migration, and apoptosis [102]. The involved pathways include MAPK and phosphatidylinositol-3-kinase. Ptd-FGFR4 is the N-terminally truncated isoform of FGFR4, characterized by cytoplasmic localization and constitutive phosphorylation [103]. Analyzed against common biomarkers of aggression and invasiveness, ptd-FGFR4 showed a direct correlation with Ki-67 [104]. Accordingly, Brito et al. documented that all patients with recurrent Cushing’s disease had high levels of FGFR4 mRNA expression [105].
2.3.5. Folate Receptor
Folate receptors (FR) are glycosylphosphatidylinositol-anchored membrane proteins involved in the absorption of folic acid, essential for cell proliferation. Among the three human isoforms (α, β, and γ), FRα is overexpressed in some forms of cancer, such as ovarian and cervical carcinomas, and, although less commonly, in lung and breast cancers. At the same time, it is poorly expressed in normal tissues [106]. FRα is strongly upregulated in NF-PitNETs but is absent or downregulated in functioning PitNETs or normal pituitary glands [107][108][109][110].
In a clinical study on 56 NF-PitNETs, a pre-operative SPECT/CT image after 99mTc-EC20-folate administration documented a sensitivity of 81% and a specificity of 83%, highlighting the possibility of an appropriate selection of patients who could take advantage of this treatment [111].
2.3.6. Estrogen Modulators
The relationship between estrogen hormones (ES) and their receptors (ESR) plays a crucial role in the pathogenesis of PitNETs. In normal human tissue, ERα (also called ER1, ESR1, and NR3A1) and Erβ (known as ER2, ESR2, and NR3A2) are nuclear receptor isoforms that respond to 17β-estradiol (E2), leading to cell proliferation and differentiation, respectively [112][113]. Among the best-known ERα receptors, ERα66 and its variant ERα36, which are situated in both the cytoplasm and plasma membrane, are mostly found [114].
Studies performed on lactotroph pituitary adenomas highlighted aggressive behavior, especially in the male gender, when the expression of ERα was reduced [115][116][117][118][119][120][121], probably associated also with the expression of genes situated on chromosome X (CTAG2, FGF13, and VEGF) that influence the ER pathway [115]. Recently, Mahboobifard et al. found a decreased expression of ERα66 and ERα36 in PRL-PitNETs compared with normal pituitary tissue. Furthermore, invasiveness was associated with low levels of ERα36 and ERα66, while an increased Ki67 index was related to decreased ERα36 expression. A significant inverse association between ERα66 with dopamine-agonist resistance and tumor size was also documented [122].
The association between ER and invasiveness is still debated [115][123][124]: Zhou et al. found a significantly higher expression of nuclear ERα staining in invasive NF-PitNETs (especially in females) than non-invasive ones. In contrast, ERβ staining decreased in invasive NF-PitNETs. Other studies found lower ERα mRNA levels in non-invasive prolactinomas but also significantly lower levels of ERα in invasive pituitary tumors [119][121][122][125].
2.3.7. Wnt/β-Catenin and E-Cadherin
The Wnt/β-catenin pathway, also known as the “canonical” Wnt pathway, has been linked to many human diseases, cancerous and non-cancerous [126]. Once activated by the canonical pathway, β-catenin translocates into the nucleus. It induces genes involved with cell proliferation and migration (e.g., c-myc, MMP) [126].
Moreover, β-catenin intervenes in cell-cell adhesion by forming a heterodimer with E-cadherin (which, in turn, inactivates the mitogenic function of β-catenin when complexed together); the E-cadherin/β-catenin anchoring complex is known to maintain epithelial cell differentiation and adhesion, and its dysregulation has been recognized as a promoter of epithelial–mesenchymal transition in many cancers [127].
The evidence suggests pituitary cells lose their normal differentiation and transition to a tumorous phenotype when membranous E-cadherin is downregulated. However, therapeutic options are not available yet. E-cadherin might represent a molecular therapy target in the future.
B-catenin shows a multimodal pattern: when analyzed alone, nuclear accumulation of B-catenin positively correlates with PitNETs aggressivity [128][129]. On the other hand, when E-cadherin is considered, B-catenin downregulation is linked to aggressive behavior and recurrence [130], further suggesting that E-cadherin serves as a proto-oncogene in PitNETs. Temozolomide has effectively reduced B-catenin activation and nuclear translocation, impairing cells vitality and promoting their apoptosis. It also reduced prolactin production in PRL-secreting adenomas in animal models [131].
2.3.8. Galectin-3
Galectin-3 (Gal-3) is a β-galactoside-binding lectin expressed in various types of cancers and plays a vital role in PitNET cell proliferation [132][133]. An increased Gal-3 expression in PRL− and ACTH-PitNETs has been described, but why and how it is involved in pituitary tumor progression is still unclear. Gal-3 levels further increase in the progression from PRL and ACTH-secreting adenomas to carcinomas; gene methylation plays a role in Gal-3 expression, and RUNX1 and RUNX2 transcription factors seem to target its gene directly, enhancing its expression [132].
2.4. Matrix MetalloProteinases
MMPs are zinc-containing calcium-dependent endopeptidases paramount in extracellular matrix (ECM) degradation and remodeling by acting on several substrates. The increasing evidence of the importance of ECM remodeling in tumor invasion supports the relevance of those enzymes in promoting PitNET local invasiveness.
MMP-9 has been widely studied as it acts by notably degrading type IV collagen, which represents the main component of the basal membrane and the medial wall of the CS. In 1996, for the first time, Kawamoto et al. observed MMP-9 expression to be significantly increased in invasive PitNETs when compared to non-invasive ones [134].
Moreover, they also found that the expression of MMP-9 was strongly associated with the recurrence-free interval, suggesting that patients with high MMP-9 expression may need particularly close clinical and radiologic follow-up after surgery [135]. Gong et al. found increased MMP-9 mRNA expression in the invasive PitNETs [136]. However, several studies also showed no correlation between MMP-2 and MMP-9 expression, as previously reported by other groups [137][138].
Protein kinase C (PKC) is a ubiquitous family of enzymes involved in several cellular pathways, such as mediating cell growth and tumor invasion by activating MMP-9. Hussaini et al. demonstrated increased expression and activity of MMP-9 in invasive NF-PitNETs and PKC-activated HP75 cell lines [137]. PKC activated MMP-9 in a highly cell-type-specific manner. Moreover, some studies found that invasive PitNETs were characterized by point mutations of PKC-α and higher overall PKC activity and expression [139][140]. The effect of several biological agents on MMP-9 activity and expression by either positively or negatively influencing PKC overall activity has been tested in different tumor models.
2.5. Angiogenesis
2.5.1. Vascular Endothelial Growth Factor
Vascular endothelial growth factor (VEGF), especially the subtype VEGF-A, represents a fundamental mediator of vasculogenesis, angiogenesis, vascular permeability, cell survival, and migration [141]. The expression of VEGF is regulated by several factors, such as hypoxia-inducible factor (HIF), epidermal growth factor (EGF), and platelet-derived growth factor (PDGF). In tumorigenesis, the upregulation of VEGF and its receptors is commonly found in most tumors, impacting their invasiveness, the formation of metastasis, and cell survival through the induction of bcl-2 expression [141][142]. Therefore, anti-VEGF therapy has been progressively introduced and approved as a secondary-line therapy [143]. The available data on VEGF expression for managing aggressive PitNETs are conflicting. Some authors identified overexpression of mRNA VEGF in invasive tumors and carcinomas, especially in the elderly, and NF-PitNETs. At the same time, other studies did not find a significant correlation between VEGF expression and tumor behavior [117][144][145][146][147].
The results from in vitro experiments have shown that the anti-VEGF-A mAb G6-31 inhibited tumor growth and decreased serum prolactin levels, especially in dopamine-agonist-resistant prolactinomas [148]. Zhou et al. experimented with Cabozantinib, an inhibitor of VEGFR2, on RBΔ19 mice, prolonging mean survival in a dose-dependent manner (15 mg/kg and 30 mg/kg) [149]. On prolactinomas, Axitinib, a VEGFR-selective tyrosine kinase inhibitor, stopped tumor progression, decreased vascular density, and, through the combination of bromocriptine, reduced tumor bleeding and normalized the vessel architecture [150].
2.5.2. Endocan
Endocan, also known as endothelial cell-specific molecule-1 or ESM-1, is a soluble chondroitin/dermatan sulfate marker of neoangiogenesis closely related to VEGF and FGF expression. Its role in physiological processes is well known and has an essential effect on inflammation and tumor pathogenesis [151]. In the normal pituitary gland, only a few endocrine cells expressed endocan, while in pituitary tumors, the data about its expression are inconsistent [152][153][154][155].
Even today, the role of Endocan in target therapy is still to be defined, especially for pituitary pathology. Studies on other tumors have shown that it is possible to block endocan expression by acting on the VEGF pathway through tyrosine kinase inhibitors or VEGF antibodies [156]. Recently, monoclonal antibodies, direct synthetic peptides, and endocan silencing have been developed, promoting apoptosis, tumor volume reduction, and cell migration in several tumors [157][158].
2.6. Genetic Aspects in Aggressive PitNETs
2.6.1. Germline Mutations
Pituitary tumorigenesis may also be caused by germline mutations that are part of a syndromic presentation or in isolation. The first identified gene underlying familial isolated pituitary adenoma (FIPA) is AIP (Aryl hydrocarbon receptor interacting protein), which accounts for about 10–20% of FIPA kindreds [159]. It encodes a co-chaperone protein, which acts as a tumor suppressor, especially versus GH and PRL PitNETs. The prevalence of AIP mutations in patients with sporadic pituitary tumors is exceedingly rare, and it is higher among acromegalic patients and patients with less than 30 years of age at diagnosis [160]. Some data suggest that low AIP expression is a better marker of invasiveness in somatotropinomas than the Ki-67 labeling index and p53 [161].
2.6.2. Somatic Mutations
Despite applying novel exome and whole genome sequencing techniques in the study of PitNETs, a few recurrent mutated genes have been reported [162][163]. Efforts in this respect will be paramount in developing novel therapeutic strategies and additional treatments for this kind of tumor. No apparent recurrent driver alterations have been identified for PitNETs, apart from GNAS and USP8 mutations in GH− and ACTH-PitNETs, respectively.
GNAS
GNAS is a complex imprinted gene characterized by several promoters. One of its products, the stimulatory α subunit of G-protein, is paramount in regulating the enzymatic activity of adenylyl cyclase. Tumor cell proliferation in PIT1 might be associated with GNAS copy number gain [164]. At the same time, mutations affecting GNAS codons 201 or 227 may lead to prolonged GTPase activity and increased cAMP levels, playing a role in GH secretion in acromegalic patients [165].
Increasing evidence suggests that GNAS-mutated tumors are more likely to be smaller, non-invasive, and better respond to SSAs [165][166][167][168], despite some authors showing no differences in tumor extension or response to SSAs between mutated and non-mutated tumors [169].
USP8, also known as ubiquitin-specific protease Y (UBPY), is a deubiquitinating enzyme displaying elevated activity toward EGFR and other receptor tyrosine kinases (RTKs) with similar signaling cascades [170][171]. USP8 gain-of-function mutations protected EGFR from lysosomal degradation, thereby increasing its expression. Interestingly, USP8 mutations have been only documented in sporadic Cushing’s disease. The most common somatic mutation involved exon 14, which encoded a putative binding domain [172].
USP8-mutated tumors are associated with an earlier onset and a higher risk of recurrence [173]. Retrospectively analyzing their cohort of patients, Albani et al. reported that the incidence of relapse over a 10-year follow-up in patients who obtained biochemical remission after surgery was higher in patients with tumors with the USP8 mutation [174]. In another cohort of pediatric patients with Cushing’s disease, recurrences have been documented in tumors with USP8 mutations [175].
2.7. Future Perspectives in PitNETs: The Role of Epigenetic
2.7.1. DNA Methylation
The development of new DNA methylation detection and analysis techniques has provided novel information on the pathogenesis of pituitary tumors. In 2012, the first genome-wide analysis of the DNA methylome in PitNETs examined more than 14,000 genes [176]. Since its early applications, methylome profiling has appeared helpful in understanding PitNET pathophysiology. However, up to date, global DNA methylation profiling has failed to predict clinically significant differences among pituitary tumors [177].
DNA methylation is crucial in PitNETs pathophysiology, mainly driving the loss or reduced expression of tumor suppressor genes (TSGs) [178]. Many of those genes have been shown to harbor C-phosphate-G (CpG) island hypermethylation, determining their silencing. Ling et al. identified 34 CpGs on 17 genes, characterized as being hypomethylated in invasive NF-PitNETs compared to non-invasive ones. Interestingly, among differentially methylated genes FLT1 (Fms-related receptor tyrosine kinase 1) and SLIT3 (slit guidance ligand 3), most of them were associated with invasion and cell migration [179].
Interestingly, a single miR may have different target genes. Duan et al. showed that the ectopic expression of miR-137, which is reduced in pituitary tumor tissues compared to normal controls, interfered with the proliferation and invasion of pituitary tumor cells via targeting AKT2 (AKT serine/threonine kinase 2) [180]. Moreover, Wei et al. reported miR-137 to target EGFR, thus inhibiting cell proliferation of GH3 cells and inducing apoptosis and G1-phase arrest. According to the authors, miR-137 mimic and AZD9291 (Osimertinib, an oral third-generation EGFR tyrosine kinase inhibitor)-mediated additive effects in GH3 cells [41].
2.8. Molecular Target Therapy and Clinical Prospects
Molecular assessments, such as those described in the present work, could represent the key factor for the treatment of those subtypes of aggressive PitNETs that are not responsive to conventional therapies. In this view, the 2017 and 2022 WHO classifications of endocrine tumors tried to define and identify this subgroup of neoplasia [181][182]. In particular, the 2017 WHO classifications defined as “high-risk categories“ prolactinomas in males, poorly granulated somatotrophs, silent corticotrophs, Crooke cells, PIT1-positive silent plurihormonal PitNETs, and high Ki67% index PitNETs [181]. The recent update of the 2022 WHO PitNETs classification added the role of tumor cell lineages, distinguishing between immature and mature tumors of the PIT1 lineage and defining the inclusion features of the PIT1 lineage “family” [182].
References
- Melmed, S. Pituitary-Tumor Endocrinopathies. N. Engl. J. Med. 2020, 382, 937–950.
- Saeger, W.; Lüdecke, D.K.; Buchfelder, M.; Fahlbusch, R.; Quabbe, H.J.; Petersenn, S. Pathohistological classification of pituitary tumors: 10 years of experience with the German Pituitary Tumor Registry. Eur. J. Endocrinol. 2007, 156, 203–216.
- Trouillas, J.; Jaffrain-Rea, M.L.; Vasiljevic, A.; Raverot, G.; Roncaroli, F.; Villa, C. How to Classify the Pituitary Neuroendocrine Tumors (PitNET)s in 2020. Cancers 2020, 12, 514.
- Serioli, S.; Doglietto, F.; Fiorindi, A.; Biroli, A.; Mattavelli, D.; Buffoli, B.; Ferrari, M.; Cornali, C.; Rodella, L.; Maroldi, R.; et al. Pituitary Adenomas and Invasiveness from Anatomo-Surgical, Radiological, and Histological Perspectives: A Systematic Literature Review. Cancers 2019, 11, 1936.
- Asa, S.L.; Casar-Borota, O.; Chanson, P.; Delgrange, E.; Earls, P.; Ezzat, S.; Grossman, A.; Ikeda, H.; Inoshita, N.; Karavitaki, N.; et al. From pituitary adenoma to pituitary neuroendocrine tumor (PitNET): An International Pituitary Pathology Club proposal. Endocr. Relat. Cancer 2017, 24, C5–C8.
- Raverot, G.; Ilie, M.D.; Lasolle, H.; Amodru, V.; Trouillas, J.; Castinetti, F.; Brue, T. Aggressive pituitary tumors and pituitary carcinomas. Nat. Rev. Endocrinol. 2021, 17, 671–684.
- Raverot, G.; Burman, P.; McCormack, A.; Heaney, A.; Petersenn, S.; Popovic, V.; Trouillas, J.; Dekkers, O.M. European Society of Endocrinology. Clinical Practice Guidelines for the management of aggressive pituitary tumors and carcinomas. Eur. J. Endocrinol. 2018, 178, G1–G24.
- Ilie, M.D.; Jouanneau, E.; Raverot, G. Aggressive Pituitary Adenomas and Carcinomas. Endocrinol. Metab. Clin. N. Am. 2020, 49, 505–515.
- Ji, Y.; Vogel, R.I.; Lou, E. Temozolomide treatment of pituitary carcinomas and atypical adenomas: Systematic review of case reports. Neurooncol. Pract. 2016, 3, 188–195.
- Lasolle, H.; Cortet, C.; Castinetti, F.; Cloix, L.; Caron, P.; Delemer, B.; Desailloud, R.; Jublanc, C.; Lebrun-Frenay, C.; Sadoul, J.L.; et al. Temozolomide treatment can improve overall survival in aggressive pituitary tumors and pituitary carcinomas. Eur. J. Endocrinol. 2017, 176, 769–777.
- Chiloiro, S.; De Marinis, L. The immune microenviroment in somatotropinomas: From biology to personalized and target therapy. Rev. Endocr. Metab. Disord. 2023, 24, 283–295.
- Ilie, M.D.; Vasiljevic, A.; Bertolino, P.; Raverot, G. Biological and Therapeutic Implications of the Tumor Microenvironment in Pituitary Adenomas. Endocr. Rev. 2023, 44, 297–311.
- Yamamoto, M.; Nakao, T.; Ogawa, W.; Fukuoka, H. Aggressive Cushing’s Disease: Molecular Pathology and Its Therapeutic Approach. Front. Endocrinol. 2021, 12, 650791.
- Ilie, M.D.; Vasiljevic, A.; Raverot, G.; Bertolino, P. The Microenvironment of Pituitary Tumors—Biological and Therapeutic Implications. Cancers 2019, 11, 1605.
- Zhang, J.; Gu, C.; Song, Q.; Zhu, M.; Xu, Y.; Xiao, M.; Zheng, W. Identifying cancer-associated fibroblasts as emerging targets for hepatocellular carcinoma. Cell Biosci. 2020, 10, 127.
- Marques, P.; Korbonits, M. Tumor microenvironment and pituitary tumor behaviour. J. Endocrinol. Investig. 2023, 46, 1047–1063.
- Principe, M.; Chanal, M.; Ilie, M.D.; Ziverec, A.; Vasiljevic, A.; Jouanneau, E.; Hennino, A.; Raverot, G.; Bertolino, P. Immune landscape of pituitary tumors reveals association between macrophages and gonadotroph tumor invasion. J. Clin. Endocrinol. Metab. 2020, 105, dgaa520.
- Zhou, W.; Zhang, C.; Zhang, D.; Peng, J.; Ma, S.; Wang, X.; Guan, X.; Li, P.; Li, D.; Jia, G.; et al. Comprehensive analysis of the immunological landscape of pituitary adenomas: Implications of immunotherapy for pituitary adenomas. J. Neurooncol. 2020, 149, 473–487.
- Lyu, L.; Jiang, Y.; Ma, W.; Li, H.; Liu, X.; Li, L.; Shen, A.; Yu, Y.; Jiang, S.; Li, H.; et al. Single-cell sequencing of PIT1- positive pituitary adenoma highlights the pro-tumor microenvironment mediated by IFN-gamma-induced tumor-associated fbroblasts remodelling. Br. J. Cancer 2023, 128, 1117–1133.
- Marques, P.; Barry, S.; Carlsen, E.; Collier, D.; Ronaldson, A.; Awad, S.; Dorward, N.; Grieve, J.; Mendoza, N.; Muquit, S.; et al. Chemokines modulate the tumor microenvironment in pituitary neuroendocrine tumors. Acta Neuropathol. Commun. 2019, 7, 172.
- Lv, L.; Zhang, S.; Hu, Y.; Zhou, P.; Gao, L.; Wang, M.; Sun, Z.; Chen, C.; Yin, S.; Wang, X.; et al. Invasive pituitary adenomaderived tumor-associated fibroblasts promote tumor progression both in vitro and in vivo. Exp. Clin. Endocrinol. Diabetes 2018, 126, 213–221.
- Marques, P.; Barry, S.; Carlsen, E.; Collier, D.; Ronaldson, A.; Awad, S.; Dorward, N.; Grieve, J.; Mendoza, N.; Muquit, S.; et al. Pituitary tumor fibroblast derived cytokines influence tumor aggressiveness. Endocr. Relat. Cancer 2019, 26, 853–865.
- Devnath, S.; Inoue, K. An insight to pituitary folliculo-stellate cells. J. Neuroendocr. 2008, 20, 687–691.
- Ilie, M.D.; Vasiljevic, A.; Chanal, M.; Gadot, N.; Chinezu, L.; Jouanneau, E.; Hennino, A.; Raverot, G.; Bertolino, P. Intratumoral spatial distribution of S100B + folliculostellate cells is associated with proliferation and expression of FSH and ERalpha in gonadotroph tumors. Acta Neuropathol. Commun. 2022, 10, 18.
- Voit, D.; Saeger, W.; Lüdecke, D.K. Folliculo-stellate cells in pituitary adenomas of patients with acromegaly. Pathol. Res. Pract. 1999, 195, 143–147.
- Mei, Y.; Bi, W.L.; Agolia, J.; Hu, C.; Larsen, A.M.G.; Meredith, D.M.; Al Abdulmohsen, S.; Bale, T.; Dunn, G.P.; Abedalthagafi, M.; et al. Immune profiling of pituitary tumors reveals variations in immune infiltration and checkpoint molecule expression. Pituitary 2021, 24, 359–373.
- Voellger, B.; Zhang, Z.; Benzel, J.; Wang, J.; Lei, T.; Nimsky, C.; Bartsch, J.W. Targeting Aggressive Pituitary Adenomas at the Molecular Level—A Review. J. Clin. Med. 2021, 11, 124.
- Duhamel, C.; Ilie, M.D.; Salle, H.; Nassouri, A.S.; Gaillard, S.; Deluche, E.; Assaker, R.; Mortier, L.; Cortet, C.; Raverot, G. Immunotherapy in Corticotroph and Lactotroph Aggressive Tumors and Carcinomas: Two Case Reports and a Review of the Literature. J. Pers. Med. 2020, 10, 88.
- Majd, N.; Waguespack, S.G.; Janku, F.; Fu, S.; Penas-Prado, M.; Xu, M.; Alshawa, A.; Kamiya-Matsuoka, C.; Raza, S.M.; McCutcheon, I.E.; et al. Efficacy of pembrolizumab in patients with pituitary carcinoma: Report of four cases from a phase II study. J. Immunother. Cancer 2020, 8, e001532.
- Lin, A.L.; Tabar, V.; Young, R.J.; Cohen, M.; Cuaron, J.; Yang, T.J.; Rosenblum, M.; Rudneva, V.A.; Geer, E.B.; Bodei, L. Synergism of Checkpoint Inhibitors and Peptide Receptor Radionuclide Therapy in the Treatment of Pituitary Carcinoma. J. Endocr. Soc. 2021, 5, bvab133.
- Chiloiro, S.; Giampietro, A.; Gessi, M.; Lauretti, L.; Mattogno, P.P.; Cerroni, L.; Carlino, A.; De Alessandris, Q.G.; Olivi, A.; Rindi, G.; et al. CD68+ and CD8+ immune cells are associated with the growth pattern of somatotroph tumors and response to first generation somatostatin analogs. J. Neuroendocr. 2023, 35, e13263.
- Thiele, J.O.; Lohrer, P.; Schaaf, L.; Feirer, M.; Stummer, W.; Losa, M.; Lange, M.; Tichomirowa, M.; Arzt, E.; Stalla, G.K.; et al. Functional in vitro studies on the role and regulation of interleukin-6 in human somatotroph pituitary adenomas. Eur. J. Endocrinol. 2003, 149, 455–461.
- Zatelli, M.C.; Piccin, D.; Vignali, C.; Tagliati, F.; Ambrosio, M.R.; Bondanelli, M.; Cimino, V.; Bianchi, A.; Schmid, H.A.; Scanarini, M.; et al. Pasireotide, a multiple somatostatin receptor subtypes ligand, reduces cell viability in non-functioning pituitary adenomas by inhibiting vascular endothelial growth factor secretion. Endocr. Relat. Cancer 2007, 14, 91–102.
- Beck, G.C.H.; Brinkkoetter, P.; Hanusch, C.; Schulte, J.; van Ackern, K.; van der Woude, F.J.; Yard, B.A. Clinical review: Immunomodulatory effects of dopamine in general inflammation. Crit. Care 2004, 8, 485–491.
- Casanueva, F.F.; Molitch, M.E.; Schlechte, J.A.; Abs, R.; Bonert, V.; Bronstein, M.D.; Brue, T.; Cappabianca, P.; Colao, A.; Fahlbusch, R.; et al. Guidelines of the Pituitary Society for the diagnosis and management of prolactinomas. Clin. Endocrinol. 2006, 65, 265–273.
- Recouvreux, M.V.; Camilletti, M.A.; Rifkin, D.B.; Díaz-Torga, G. The pituitary TGFbeta1 system as a novel target for the treatment of resistant prolactinomas. J. Endocrinol. 2016, 228, R73–R83.
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976.
- McCubrey, J.A.; Steelman, L.S.; Chappell, W.H.; Abrams, S.L.; Wong, E.W.; Chang, F.; Lehmann, B.; Terrian, D.M.; Milella, M.; Tafuri, A.; et al. Roles of the Raf/MEK/ERK pathway in cell growth, malignant transformation and drug resistance. Biochim. Biophys. Acta 2007, 1773, 1263–1284.
- Guo, Y.J.; Pan, W.W.; Liu, S.B.; Shen, Z.F.; Xu, Y.; Hu, L.L. ERK/MAPK signalling pathway and tumorigenesis. Exp. Ther. Med. 2020, 19, 1997–2007.
- Vlotides, G.; Siegel, E.; Donangelo, I.; Gutman, S.; Ren, S.G.; Melmed, S. Rat Prolactinoma cell growth regulation by Epidermal Growth Factor receptor ligands. Cancer Res. 2008, 68, 6377–6386.
- Wei, D.; Yu, Z.; Cheng, Y.; Jiawei, H.; Jian, G.; Hua, G.; Guilan, D. Dysregulated miR-137 and its target EGFR contribute to the progression of pituitary adenomas. Mol. Cell Endocrinol. 2021, 520, 111083.
- Wei, D.; Yu, Z.; Cheng, Y.; Jiawei, H.; Jian, G.; Hua, G.; Guilan, D. ADAM12 induces EMT and promotes cell migration, invasion and proliferation in pituitary adenomas via EGFR/ERK signaling pathway. Biomed. Pharmacother. 2018, 97, 1066–1077.
- Rose, A.; Froment, P.; Perrot, V.; Quon, M.J.; LeRoith, D.; Dupont, J. The Luteinizing Hormone-releasing Hormone Inhibits the Anti-apoptotic Activity of Insulin-like Growth Factor-1 in Pituitary T3 Cells by Protein Kinase C-mediated Negative Regulation of Akt. J. Biol. Chem. 2004, 279, 52500–52516.
- Fernández, M.; Sánchez-Franco, F.; Palacios, N.; Sánchez, I.; Fernández, C.; Cacicedo, L. IGF-I inhibits apoptosis through the activation of the phosphatidylinositol 3-kinase/Akt pathway in pituitary cells. J. Mol. Endocrinol. 2004, 33, 155–163.
- Kowarik, M.; Onofri, C.; Colaco, T.; Stalla, G.K.; Renner, U. Platelet-derived Growth Factor (PDGF) and PDGF Receptor Expression and Function in Folliculostellate Pituitary Cells. Exp. Clin. Endocrinol. Diabetes 2010, 118, 113–120.
- Murat, C.B.; Braga, P.B.; Fortes, M.A.; Bronstein, M.D.; Corrêa-Giannella, M.L.; Giorgi, R.R. Mutation and genomic amplification of the PIK3CA proto-oncogene in pituitary adenomas. Braz. J. Med. Biol. Res. 2012, 45, 851–855.
- Lin, Y.; Jiang, X.; Shen, Y.; Li, M.; Ma, H.; Xing, M.; Lu, Y. Frequent mutations and amplifications of the PIK3CA gene in pituitary tumors. Endocr. Relat. Cancer 2009, 16, 301–310.
- Musat, M.; Korbonits, M.; Kola, B.; Borboli, N.; Hanson, M.R.; Nanzer, A.M.; Grigson, J.; Jordan, S.; Morris, D.G.; Gueorguiev, M.; et al. Enhanced protein kinase B/Akt signalling in pituitary tumors. Endocr. Relat. Cancer 2005, 12, 423–433.
- Jia, W.; Sanders, A.J.; Jia, G.; Liu, X.; Lu, R.; Jiang, W.G. Expression of the mTOR pathway regulators in human pituitary adenomas indicates the clinical course. Anticancer Res. 2013, 33, 3123–3131.
- Xu, Q.; Yu, Z.X.; Xie, Y.L.; Bai, L.; Liang, S.R.; Ji, Q.H.; Zhou, J. MicroRNA-137 inhibits pituitary prolactinoma proliferation by targeting AKT2. J. Endocrinol. Investig. 2023, 46, 1145–1154.
- Noh, T.W.; Jeong, H.J.; Lee, M.K.; Kim, T.S.; Kim, S.H.; Lee, E.J. Predicting Recurrence of Nonfunctioning Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2009, 94, 4406–4413.
- Zhou, K.; Zhang, T.; Fan, Y.; Serick Du, G.; Wu, P.; Geng, D. MicroRNA-106b promotes pituitary tumor cell proliferation and invasion through PI3K/AKT signaling pathway by targeting PTEN. Tumor Biol. 2016, 37, 13469–13477.
- Zhang, D. Effect of Everolimus in Treatment of Aggressive Prolactin-Secreting Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2019, 104, 1929–1936.
- Lu, C.; Willingham, M.C.; Furuya, F.; Cheng, S.Y. Activation of Phosphatidylinositol 3-Kinase Signaling Promotes Aberrant Pituitary Growth in a Mouse Model of Thyroid-Stimulating Hormone-Secreting Pituitary Tumors. Endocrinology 2008, 149, 3339–3345.
- Gorshtein, A.; Rubinfeld, H.; Kendler, E.; Theodoropoulou, M.; Cerovac, V.; Stalla, G.K.; Cohen, Z.R.; Hadani, M.; Shimon, I. Mammalian target of rapamycin inhibitors rapamycin and RAD001 (everolimus) induce anti-proliferative effects in GH-secreting pituitary tumor cells in vitro. Endocr. Relat. Cancer 2009, 16, 1017–1027.
- Sajjad, E.A.; Zieliński, G.; Maksymowicz, M.; Hutnik, Ł.; Bednarczuk, T.; Włodarski, P. mTOR is Frequently Active in GH-Secreting Pituitary Adenomas without Influencing their Morpho-pathological Features. Endocr. Pathol. 2013, 24, 11–19.
- Dworakowska, D.; Wlodek, E.; Leontiou, C.A.; Igreja, S.; Cakir, M.; Teng, M.; Prodromou, N.; Góth, M.I.; Grozinsky-Glasberg, S.; Gueorguiev, M.; et al. Activation of RAF/MEK/ERK and PI3K/AKT/mTOR pathways in pituitary adenomas and their effects on downstream effectors. Endocr. Relat. Cancer 2009, 16, 1329–1338.
- Yao, H.; Tang, H.; Zhang, Y.; Zhang, Q.F.; Liu, X.Y.; Liu, Y.T.; Gu, W.T.; Zheng, Y.Z.; Shang, H.B.; Wang, Y.; et al. DEPTOR inhibits cell proliferation and confers sensitivity to dopamine agonist in pituitary adenoma. Cancer Lett. 2019, 459, 135–144.
- Jian, F.; Chen, Y.; Ning, G.; Fu, W.; Tang, H.; Chen, X.; Zhao, Y.; Zheng, L.; Pan, S.; Wang, W.; et al. Cold inducible RNA binding protein upregulation in pituitary corticotroph adenoma induces corticotroph cell proliferation via Erk signaling pathway. Oncotarget 2016, 7, 9175–9187.
- Cheng, S.Q.; Fan, H.Y.; Xu, X.; Gao, W.W.; Lv, S.G.; Ye, M.H.; Wu, M.J.; Shen, X.L.; Cheng, Z.J.; Zhu, X.G.; et al. Over-expression of LRIG1 Suppresses Biological Function of Pituitary Adenoma via Attenuation of PI3K/AKT and Ras/Raf/ERK Pathways In Vivo and In Vitro. J. Huazhong Univ. Sci. Technol. Med. Sci. 2016, 36, 558–563.
- Liu, J.; Wang, J.; Tian, W.; Xu, Y.; Li, R.; Zhao, K.; You, C.; Zhu, Y.; Bartsch, J.W.; Niu, H.; et al. PDCD10 promotes the aggressive behaviors of pituitary adenomas by up-regulating CXCR2 and activating downstream AKT/ERK signaling. Aging 2022, 14, 6066–6080.
- Romano, D.; Pertuit, M.; Rasolonjanahary, R.; Barnier, J.V.; Magalon, K.; Enjalbert, A.; Gerard, C. Regulation of the RAP1/RAF-1/Extracellularly Regulated Kinase-1/2 Cascade and Prolactin Release by the Phosphoinositide 3-Kinase/AKT Pathway in Pituitary Cells. Endocrinology 2006, 147, 6036–6045.
- Ewing, I.; Pedder-Smith, S.; Franchi, G.; Ruscica, M.; Emery, M.; Vax, V.; Garcia, E.; Czirják, S.; Hanzély, Z.; Kola, B.; et al. A mutation and expression analysis of the oncogene BRAF in pituitary adenomas. Clin. Endocrinol. 2007, 66, 348–352.
- Chen, J. Identification of recurrent USP48 and BRAF mutations in Cushing’s disease. Nat. Commun. 2018, 9, 3171.
- Sukumari-Ramesh, S.; Singh, N.; Dhandapani, K.M.; Vender, J.R. mTOR inhibition reduces cellular proliferation and sensitizes pituitary adenoma cells to ionizing radiation. Surg. Neurol. Int. 2011, 2, 22.
- Pivonello, C.; Patalano, R.; Solari, D.; Auriemma, R.S.; Frio, F.; Vitulli, F.; Grasso, L.F.S.; Di Cera, M.; De Martino, M.C.; Cavallo, L.M.; et al. Effect of combined treatment with a pan-PI3K inhibitor or an isoform-specific PI3K inhibitor and everolimus on cell proliferation in GH-secreting pituitary tumor in an experimental setting. Endocrine 2018, 62, 663–680.
- Zatelli, M.C.; Minoia, M.; Filieri, C.; Tagliati, F.; Buratto, M.; Ambrosio, M.R.; Lapparelli, M.; Scanarini, M.; Degli Uberti, E.C. Effect of Everolimus on Cell Viability in Nonfunctioning Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2010, 95, 968–976.
- Cerovac, V.; Monteserin-Garcia, J.; Rubinfeld, H.; Buchfelder, M.; Losa, M.; Florio, T.; Paez-Pereda, M.; Stalla, G.K.; Theodoropoulou, M. The Somatostatin Analogue Octreotide Confers Sensitivity to Rapamycin Treatment on Pituitary Tumor Cells. Cancer Res. 2010, 70, 666–674.
- Chanal, M.; Chevallier, P.; Raverot, V.; Fonteneau, G.; Lucia, K.; Garcia, J.L.M.; Rachwan, A.; Jouanneau, E.; Trouillas, J.; Honnorat, J.; et al. Differential Effects of PI3K and Dual PI3K/mTOR Inhibition in Rat Prolactin-Secreting Pituitary Tumors. Mol. Cancer Ther. 2016, 15, 1261–1270.
- Wang, J.; Zhang, J.; Ma, D.; Li, X. The Potential Role of CERS1 in Autophagy Through PI3K/AKT Signaling Pathway in Hypophysoma. Technol. Cancer Res. Treat. 2020, 19, 1533033820977536.
- Lee, M.; Theodoropoulou, M.; Graw, J.; Roncaroli, F.; Zatelli, M.C.; Pellegata, N.S. Levels of p27 Sensitize to Dual PI3K/mTOR Inhibition. Mol. Cancer Ther. 2011, 10, 1450–1459.
- Lee, M.; Wiedemann, T.; Gross, C.; Leinhäuser, I.; Roncaroli, F.; Braren, R.; Pellegata, N.S. Targeting PI3K/mTOR Signaling Displays Potent Antitumor Efficacy against Nonfunctioning Pituitary Adenomas. Clin. Cancer Res. 2015, 21, 3204–3215.
- Dai, C.; Zhang, B.; Liu, X.; Ma, S.; Yang, Y.; Yao, Y.; Feng, M.; Bao, X.; Li, G.; Wang, J.; et al. Inhibition of PI3K/AKT/mTOR Pathway Enhances Temozolomide-Induced Cytotoxicity in Pituitary Adenoma Cell Lines in Vitro and Xenografted Pituitary Adenoma in Female Nude Mice. Endocrinology 2013, 154, 1247–1259.
- Zeng, J.; See, A.P.; Aziz, K.; Thiyagarajan, S.; Salih, T.; Gajula, R.P.; Armour, M.; Phallen, J.; Terezakis, S.; Kleinberg, L.; et al. Nelfinavir induces radiation sensitization in pituitary adenoma cells. Cancer Biol. Ther. 2011, 12, 657–663.
- Reubi, J.C.; Waser, B.; Schaer, J.C.; Laissue, J.A. Somatostatin receptor sst1-sst5 expression in normal and neoplastic human tissues using receptor autoradiography with subtype-selective ligands. Eur. J. Nucl. Med. 2001, 28, 836–846.
- Behling, F.; Honegger, J.; Skardelly, M.; Gepfner-Tuma, I.; Tabatabai, G.; Tatagiba, M.; Schittenhelm, J. High Expression of Somatostatin Receptors 2A, 3, and 5 in Corticotroph Pituitary Adenoma. Int. J. Endocrinol. 2018, 2018, 1763735.
- Peverelli, E.; Giardino, E.; Treppiedi, D.; Catalano, R.; Mangili, F.; Locatelli, M.; Lania, A.G.; Arosio, M.; Spada, A.; Mantovani, G. A novel pathway activated by somatostatin receptor type 2 (SST2): Inhibition of pituitary tumor cell migration and invasion through cytoskeleton protein recruitment. Int. J. Cancer 2018, 142, 1842–1852.
- Peverelli, E.; Giardino, E.; Treppiedi, D.; Locatelli, M.; Vaira, V.; Ferrero, S.; Bosari, S.; Lania, A.G.; Spada, A.; Mantovani, G. Dopamine receptor type 2 (DRD2) inhibits migration and invasion of human tumorous pituitary cells through ROCK-mediated cofilin inactivation. Cancer Lett. 2016, 381, 279–286.
- Ibáñez-Costa, A.; López-Sánchez, L.M.; Gahete, M.D.; Rivero-Cortés, E.; Vázquez-Borrego, M.C.; Gálvez, M.A.; de la Riva, A.; Venegas-Moreno, E.; Jiménez-Reina, L.; Moreno-Carazo, A.; et al. BIM-23A760 influences key functional endpoints in pituitary adenomas and normal pituitaries: Molecular mechanisms underlying the differential response in adenomas. Sci. Rep. 2017, 7, 42002.
- De Bruin, C.; Pereira, A.M.; Feelders, R.A.; Romijn, J.A.; Roelfsema, F.; Sprij-Mooij, D.M.; van Aken, M.O.; van der Lelij, A.J.; de Herder, W.W.; Lamberts, S.W.; et al. Coexpression of dopamine and somatostatin receptor subtypes in corticotroph adenomas. J. Clin. Endocrinol. Metab. 2009, 94, 1118–1124.
- Luque, R.M.; Ibáñez-Costa, A.; Neto, L.V.; Taboada, G.F.; Hormaechea-Agulla, D.; Kasuki, L.; Venegas-Moreno, E.; Moreno-Carazo, A.; Gálvez, M.Á.; Soto-Moreno, A.; et al. Truncated somatostatin receptor variant sst5TMD4 confers aggressive features (proliferation, invasion and reduced octreotide response) to somatotropinomas. Cancer Lett. 2015, 359, 299–306.
- Lee, M.; Lupp, A.; Mendoza, N.; Martin, N.; Beschorner, R.; Honegger, J.; Schlegel, J.; Shively, T.; Pulz, E.; Schulz, S.; et al. SSTR3 is a putative target for the medical treatment of gonadotroph adenomas of the pituitary. Endocr. Relat. Cancer 2015, 22, 111–119.
- Hofland, J.; Brabander, T.; Verburg, F.A.; Feelders, R.A.; de Herder, W.W. Peptide Receptor Radionuclide Therapy. J. Clin. Endocrinol. Metab. 2022, 107, 3199–3208.
- Gonzalez, P.; Debnath, S.; Chen, Y.A.; Hernandez, E.; Jha, P.; Dakanali, M.; Hsieh, J.T.; Sun, X. A Theranostic Small-Molecule Prodrug Conjugate for Neuroendocrine Prostate Cancer. Pharmaceutics 2023, 15, 481.
- Gupta, S.K.; Singla, S.; Damle, N.A.; Agarwal, K.; Bal, C. Diagnosis of Men-I Syndrome on (68)Ga-DOTANOC PET-CT and Role of Peptide Receptor Radionuclide Therapy with (177)Lu-DOTATATE. Int. J. Endocrinol. Metab. 2012, 10, 629–633.
- Baldari, S.; Ferraù, F.; Alafaci, C.; Herberg, A.; Granata, F.; Militano, V.; Salpietro, F.M.; Trimarchi, F.; Cannavò, S. First demonstration of the effectiveness of peptide receptor radionuclide therapy (PRRT) with 111In-DTPA-octreotide in a giant PRL-secreting pituitary adenoma resistant to conventional treatment. Pituitary 2012, 15 (Suppl. S1), S57–S60.
- Giuffrida, G.; Ferraù, F.; Laudicella, R.; Cotta, O.R.; Messina, E.; Granata, F.; Angileri, F.F.; Vento, A.; Alibrandi, A.; Baldari, S.; et al. Peptide receptor radionuclide therapy for aggressive pituitary tumors: A monocentric experience. Endocr. Connect. 2019, 8, 528–535.
- Priola, S.M.; Esposito, F.; Cannavò, S.; Conti, A.; Abbritti, R.V.; Barresi, V.; Baldari, S.; Ferraù, F.; Germanò, A.; Tomasello, F.; et al. Aggressive Pituitary Adenomas: The Dark Side of the Moon. World Neurosurg. 2017, 97, 140–155.
- Kovács, G.L.; Góth, M.; Rotondo, F.; Scheithauer, B.W.; Carlsen, E.; Saadia, A.; Hubina, E.; Kovács, L.; Szabolcs, I.; Nagy, P.; et al. ACTH-secreting Crooke cell carcinoma of the pituitary. Eur. J. Clin. Investig. 2013, 43, 20–26.
- Komor, J.; Reubi, J.C.; Christ, E.R. Peptide receptor radionuclide therapy in a patient with disabling non-functioning pituitary adenoma. Pituitary 2014, 17, 227–231.
- Maclean, J.; Aldridge, M.; Bomanji, J.; Short, S.; Fersht, N. Peptide receptor radionuclide therapy for aggressive atypical pituitary adenoma/carcinoma: Variable clinical response in preliminary evaluation. Pituitary 2014, 17, 530–538.
- Bengtsson, D.; Schrøder, H.D.; Andersen, M.; Maiter, D.; Berinder, K.; Rasmussen, U.F.; Rasmussen, Å.K.; Johannsson, G.; Hoybye, C.; van der Lely, A.J.; et al. Long-term outcome and MGMT as a predictive marker in 24 patients with atypical pituitary adenomas and pituitary carcinomas given treatment with temozolomide. J. Clin. Endocrinol. Metab. 2015, 100, 1689–1698.
- Burman, P.; Trouillas, J.; Losa, M.; McCormack, A.; Petersenn, S.; Popovic, V.; Theodoropoulou, M.; Raverot, G.; Dekkers, O.M.; ESE Survey Collaborators. Aggressive pituitary tumors and carcinomas, characteristics and management of 171 patients. Eur. J. Endocrinol. 2022, 187, 593–605.
- Novruzov, F.; Aliyev, A.; Wan, M.Y.S.; Syed, R.; Mehdi, E.; Aliyeva, I.; Giammarile, F.; Bomanji, J.B.; Kayani, I. The value of Ga-DOTA-TATE PET/CT in diagnosis and management of suspected pituitary tumors. Eur. J. Hybrid Imaging 2021, 5, 10.
- Waligórska-Stachura, J.; Gut, P.; Sawicka-Gutaj, N.; Liebert, W.; Gryczyńska, M.; Baszko-Błaszyk, D.; Blanco-Gangoo, A.R.; Ruchała, M. Growth hormone-secreting macroadenoma of the pituitary gland successfully treated with the radiolabeled somatostatin analog (90)Y-DOTATATE: Case report. J. Neurosurg. 2016, 125, 346–349.
- Assadi, M.; Nemati, R.; Shooli, H.; Rekabpour, S.J.; Nabipour, I.; Jafari, E.; Gholamrezanezhad, A.; Amini, A.; Ahmadzadehfar, H. An aggressive functioning pituitary adenoma treated with peptide receptor radionuclide therapy. Eur. J. Nucl. Med. Mol. Imaging 2020, 47, 1015–1016.
- Shi, Y.; Massagué, J. Mechanisms of TGF-beta signaling from cell membrane to the nucleus. Cell 2003, 113, 685–700.
- Massagué, J. TGFbeta in Cancer. Cell 2008, 134, 215–230.
- Jiang, M.; Mou, C.Z.; Han, T.; Wang, M.; Yang, W. Thrombospondin-1 and transforming growth factor-β1 levels in prolactinoma and their clinical significance. J. Int. Med. Res. 2012, 40, 1284–1294.
- Elenkova, A.; Atanassova, I.; Kirilov, G.; Vasilev, V.; Kalinov, K.; Zacharieva, S. Transforming growth factor β1 is not a reliable biomarker for valvular fibrosis but could be a potential serum marker for invasiveness of prolactinomas (pilot study). Eur. J. Endocrinol. 2013, 169, 299–306.
- Gu, Y.H.; Feng, Y.G. Down-regulation of TGF-β RII expression is correlated with tumor growth and invasion in non-functioning pituitary adenomas. J. Clin. Neurosci. 2018, 47, 264–268.
- Eswarakumar, V.P.; Lax, I.; Schlessinger, J. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor. Rev. 2005, 16, 139–149.
- Giraldi, F.P.; Moro, M.; Cavagnini, F.; Study Group on the Hypothalamo-Pituitary-Adrenal Axis of the Italian Society of Endocrinology. Gender-related differences in the presentation and course of Cushing’s disease. J. Clin. Endocrinol. Metab. 2003, 88, 1554–1558.
- Qian, Z.R.; Sano, T.; Asa, S.L.; Yamada, S.; Horiguchi, H.; Tashiro, T.; Li, C.C.; Hirokawa, M.; Kovacs, K.; Ezzat, S. Cytoplasmic expression of fibroblast growth factor receptor-4 in human pituitary adenomas: Relation to tumor type, size, proliferation, and invasiveness. J. Clin. Endocrinol. Metab. 2004, 89, 1904–1911.
- Brito, L.P.; Lerário, A.M.; Bronstein, M.D.; Soares, I.C.; Mendonca, B.B.; Fragoso, M.C. Influence of the fibroblast growth factor receptor 4 expression and the G388R functional polymorphism on Cushing’s disease outcome. J. Clin. Endocrinol. Metab. 2010, 95, E271–E279.
- Nawaz, F.Z.; Kipreos, E.T. Emerging roles for folate receptor FOLR1 in signaling and cancer. Trends Endocrinol. Metab. 2022, 33, 159–174.
- Evans, C.O.; Reddy, P.; Brat, D.J.; O’Neill, E.B.; Craige, B.; Stevens, V.L.; Oyesiku, N.M. Differential expression of folate receptor in pituitary adenomas. Cancer Res. 2003, 63, 4218–4224.
- Ding, Z.; Li, B.; Wang, Q.; Miao, Y.; Lu, X. Increase in folate receptor alpha expression in nonfunctional pituitary adenomas. Turk. Neurosurg. 2015, 25, 298–304.
- Liu, X.; Ma, S.; Yao, Y.; Li, G.; Feng, M.; Deng, K.; Dai, C.; Cai, F.; Li, Y.; Zhang, B.; et al. Differential expression of folate receptor alpha in pituitary adenomas and its relationship to tumor behavior. Neurosurgery 2012, 70, 1274–1280, discussion 1280.
- Dai, C.; Cai, F.; Hwang, K.C.; Zhou, Y.; Zhang, Z.; Liu, X.; Ma, S.; Yang, Y.; Yao, Y.; Feng, M.; et al. Folate receptor-mediated boron-10 containing carbon nanoparticles as potential delivery vehicles for boron neutron capture therapy of nonfunctional pituitary adenomas. Sci. China Life Sci. 2013, 56, 163–173.
- Galt, J.R.; Halkar, R.K.; Evans, C.O.; Osman, N.A.; LaBorde, D.; Fox, T.H.; Faraj, B.A.; Kumar, K.; Wang, H.; Oyesiku, N.M. In vivo assay of folate receptors in nonfunctional pituitary adenomas with 99mTc-folate SPECT/CT. J. Nucl. Med. 2010, 51, 1716–1723.
- Paterni, I.; Granchi, C.; Katzenellenbogen, J.A.; Minutolo, F. Estrogen receptors alpha (ERα) and beta (ERβ): Subtype-selective ligands and clinical potential. Steroids 2014, 90, 13–29.
- Yaşar, P.; Ayaz, G.; User, S.D.; Güpür, G.; Muyan, M. Molecular mechanism of estrogen-estrogen receptor signaling. Reprod. Med. Biol. 2016, 16, 4–20.
- Wang, Z.; Zhang, X.; Shen, P.; Loggie, B.W.; Chang, Y.; Deuel, T.F. A variant of estrogen receptor-, hER-36: Transduction of estrogen- and antiestrogen-dependent membrane-initiated mitogenic signaling. Proc. Natl. Acad. Sci. USA 2006, 103, 9063–9068.
- Gao, H.; Xue, Y.; Cao, L.; Liu, Q.; Liu, C.; Shan, X.; Wang, H.; Gu, Y.; Zhang, Y. ESR1 and its antagonist fulvestrant in pituitary adenomas. Mol. Cell. Endocrinol. 2017, 443, 32–41.
- Wierinckx, A.; Delgrange, E.; Bertolino, P.; François, P.; Chanson, P.; Jouanneau, E.; Lachuer, J.; Trouillas, J.; Raverot, G. Sex-Related Differences in Lactotroph Tumor Aggressiveness Are Associated with a Specific Gene-Expression Signature and Genome Instability. Front. Endocrinol. 2018, 9, 706.
- Trouillas, J.; Delgrange, E.; Wierinckx, A.; Vasiljevic, A.; Jouanneau, E.; Burman, P.; Raverot, G. Clinical, Pathological, and Molecular Factors of Aggressiveness in Lactotroph Tumors. Neuroendocrinology 2019, 109, 70–76.
- Delgrange, E.; Vasiljevic, A.; Wierinckx, A.; François, P.; Jouanneau, E.; Raverot, G.; Trouillas, J. Expression of estrogen receptor alpha is associated with prolactin pituitary tumor prognosis and supports the sex-related difference in tumor growth. Eur. J. Endocrinol. 2015, 172, 791–801.
- Bima, C.; Chiloiro, S.; Giampietro, A.; Gessi, M.; Mattogno, P.P.; Lauretti, L.; Anile, C.; Rindi, G.; Pontecorvi, A.; De Marinis, L.; et al. Galectin-3 and Estrogen Receptor Alpha as Prognostic Markers in Prolactinoma: Preliminary Results from a Pilot Study. Front. Endocrinol. 2021, 12, 684055.
- Burdman, J.A.; Pauni, M.; Sereno, G.M.H.; Bordón, A.E. Estrogen receptors in human pituitary tumors. Horm. Metab. Res. 2008, 40, 524–527.
- Li, C.Z.; Gui, S.B.; Zong, X.Y.; Zhang, Y.Z. The Expression of Estrogen Receptor Subtypes in Prolactinomas and Their Relationship to Tumor Biological Behavior. Biomed. Env. Sci. 2015, 28, 820–822.
- Mahboobifard, F.; Bidari-Zerehpoosh, F.; Davoudi, Z.; Panahi, M.; Dargahi, L.; Pourgholami, M.H.; Sharifi, G.; Izadi, N.; Jorjani, M. Expression patterns of ERα66 and its novel variant isoform ERα36 in lactotroph pituitary adenomas and associations with clinicopathological characteristics. Pituitary 2020, 23, 232–245.
- Zhou, W.; Song, Y.; Xu, H.; Zhou, K.; Zhang, W.; Chen, J.; Qin, M.; Yi, H.; Gustafsson, J.A.; Yang, H.; et al. In nonfunctional pituitary adenomas, estrogen receptors and slug contribute to the development of invasiveness. J. Clin. Endocrinol. Metab. 2011, 96, E1237–E1245.
- Pereira-Lima, J.F.; Marroni, C.P.; Pizarro, C.B.; Barbosa-Coutinho, L.M.; Ferreira, N.P.; Oliveira, M.C. Immunohistochemical detection of estrogen receptor alpha in pituitary adenomas and its correlation with cellular replication. Neuroendocrinology 2004, 79, 119–124.
- Manoranjan, B.; Salehi, F.; Scheithauer, B.W.; Rotondo, F.; Kovacs, K.; Cusimano, M.D. Estrogen receptors alpha and beta immunohistochemical expression: Clinicopathological correlations in pituitary adenomas. Anticancer. Res. 2010, 30, 2897–2904.
- Liu, J.; Xiao, Q.; Xiao, J.; Niu, C.; Li, Y.; Zhang, X.; Zhou, Z.; Shu, G.; Yin, G. Wnt/β-catenin signalling: Function, biological mechanisms, and therapeutic opportunities. Signal Transduct. Target. Ther. 2022, 7, 3.
- Tian, X.; Liu, Z.; Niu, B.; Zhang, J.; Tan, T.K.; Lee, S.R.; Zhao, Y.; Harris, D.C.; Zheng, G. E-cadherin/β-catenin complex and the epithelial barrier. J. Biomed. Biotechnol. 2011, 2011, 567305.
- Liu, C.; Wu, Y.; Yu, S.; Bai, J.; Li, C.; Wu, D.; Zhang, Y. Increased β catenin and c-myc expression predict aggressive growth of non-functioning pituitary adenomas: An assessment using a tissue microarray-based approach. Mol. Med. Rep. 2017, 15, 1793–1799.
- Wu, Y.; Bai, J.; Hong, L.; Liu, C.; Yu, S.; Yu, G.; Zhang, Y. Low expression of secreted frizzled-related protein 2 and nuclear accumulation of β-catenin in aggressive nonfunctioning pituitary adenoma. Oncol. Lett. 2016, 12, 199–206.
- Zhou, K.; Jin, H.; Luo, Y. Expression and significance of E-cadherin and β-catenins in pituitary adenoma. Int. J. Surg. Pathol. 2013, 21, 363–367.
- Demarchi, G.; Valla, S.; Perrone, S.; Chimento, A.; Bonadeo, N.; Vitale, D.L.; Spinelli, F.M.; Cervio, A.; Sevlever, G.; Alaniz, L.; et al. β-Catenin is reduced in membranes of human prolactinoma cells and it is inhibited by temozolomide in prolactin secreting tumor models. Tumor Biol. 2022, 44, 85–105.
- Righi, A.; Jin, L.; Zhang, S.; Stilling, G.; Scheithauer, B.W.; Kovacs, K.; Lloyd, R.V. Identification and consequences of galectin-3 expression in pituitary tumors. Mol. Cell Endocrinol. 2010, 326, 8–14.
- Riss, D.; Jin, L.; Qian, X.; Bayliss, J.; Scheithauer, B.W.; Young, W.F., Jr.; Vidal, S.; Kovacs, K.; Raz, A.; Lloyd, R.V. Differential expression of galectin-3 in pituitary tumors. Cancer Res. 2003, 63, 2251–2255.
- Kawamoto, H.; Kawamoto, K.; Mizoue, T.; Uozumi, T.; Arita, K.; Kurisu, K. Matrix metalloproteinase-9 secretion by human pituitary adenomas detected by cell immunoblot analysis. Acta Neurochir. 1996, 138, 1442–1448.
- Liu, X.; Feng, M.; Zhang, Y.; Dai, C.; Sun, B.; Bao, X.; Deng, K.; Yao, Y.; Wang, R. Expression of Matrix Metalloproteinase-9, Pituitary Tumor Transforming Gene, High Mobility Group A 2, and Ki-67 in Adrenocorticotropic Hormone-Secreting Pituitary Tumors and Their Association with Tumor Recurrence. World Neurosurg. 2018, 113, e213–e221.
- Gong, J.; Zhao, Y.; Abdel-Fattah, R.; Amos, S.; Xiao, A.; Lopes, M.B.; Hussaini, I.M.; Laws, E.R. Matrix metalloproteinase-9, a potential biological marker in invasive pituitary adenomas. Pituitary 2008, 11, 37–48.
- Hussaini, I.M.; Trotter, C.; Zhao, Y.; Abdel-Fattah, R.; Amos, S.; Xiao, A.; Agi, C.U.; Redpath, G.T.; Fang, Z.; Leung, G.K.; et al. Matrix metalloproteinase-9 is differentially expressed in nonfunctioning invasive and noninvasive pituitary adenomas and increases invasion in human pituitary adenoma cell line. Am. J. Pathol. 2007, 170, 356–365.
- Beaulieu, E.; Kachra, Z.; Mousseau, N.; Delbecchi, L.; Hardy, J.; Béliveau, R. Matrix metalloproteinases and their inhibitors in human pituitary tumors. Neurosurgery 1999, 45, 1432–1441.
- Alvaro, V.; Lévy, L.; Dubray, C.; Roche, A.; Peillon, F.; Quérat, B.; Joubert, D. Invasive human pituitary tumors express a point-mutated alpha-protein kinase-C. J. Clin. Endocrinol. Metab. 1993, 77, 1125–1129.
- Alvaro, V.; Touraine, P.; Vozari, R.R.; Bai-Grenier, F.; Birman, P.; Joubert, D. Protein kinase C activity and expression in normal and adenomatous human pituitaries. Int. J. Cancer 1992, 50, 724–730.
- Roskoski, R., Jr. VEGF receptor protein-tyrosine kinases: Structure and regulation. Biochem. Biophys. Res. Commun. 2008, 375, 287–291.
- Nör, J.E.; Christensen, J.; Mooney, D.J.; Polverini, P.J. Vascular endothelial growth factor (VEGF)-mediated angiogenesis is associated with enhanced endothelial cell survival and induction of Bcl-2 expression. Am. J. Pathol. 1999, 154, 375–384.
- Dai, C.; Liang, S.; Sun, B.; Li, Y.; Kang, J. Anti-VEGF Therapy in Refractory Pituitary Adenomas and Pituitary Carcinomas: A Review. Front. Oncol. 2021, 11, 773905.
- Niveiro, M.; Aranda, F.I.; Peiró, G.; Alenda, C.; Picó, A. Immunohistochemical analysis of tumor angiogenic factors in human pituitary adenomas. Hum. Pathol. 2005, 36, 1090–1095.
- Yilmaz, M.; Vural, E.; Koc, K.; Ceylan, S. Cavernous sinus invasion and effect of immunohistochemical features on remission in growth hormone secreting pituitary adenomas. Turk. Neurosurg. 2015, 25, 380–388.
- Borg, S.A.; Kerry, K.E.; Royds, J.A.; Battersby, R.D.; Jones, T.H. Correlation of VEGF production with IL1 alpha and IL6 secretion by human pituitary adenoma cells. Eur. J. Endocrinol. 2005, 152, 293–300.
- Ghadir, M.; Khamseh, M.E.; Panahi-Shamsabad, M.; Ghorbani, M.; Akbari, H.; Mehrjardi, A.Z.; Honardoost, M.; Jafar-Mohammadi, B. Cell proliferation, apoptosis, and angiogenesis in non-functional pituitary adenoma: Association with tumor invasiveness. Endocrine 2020, 69, 596–603.
- Korsisaari, N.; Ross, J.; Wu, X.; Kowanetz, M.; Pal, N.; Hall, L.; Eastham-Anderson, J.; Forrest, W.F.; Van Bruggen, N.; Peale, F.V.; et al. Blocking vascular endothelial growth factor-A inhibits the growth of pituitary adenomas and lowers serum prolactin level in a mouse model of multiple endocrine neoplasia type 1. Clin. Cancer Res. 2008, 14, 249–258.
- Zhou, J.; Hu, Y.; Zhu, W.; Nie, C.; Zhao, W.; Faje, A.T.; Labelle, K.E.; Swearingen, B.; Lee, H.; Hedley-Whyte, E.T.; et al. Sprouting Angiogenesis in Human Pituitary Adenomas. Front. Oncol. 2022, 12, 875219.
- Chauvet, N.; Romanò, N.; Lafont, C.; Guillou, A.; Galibert, E.; Bonnefont, X.; Le Tissier, P.; Fedele, M.; Fusco, A.; Mollard, P.; et al. Complementary actions of dopamine D2 receptor agonist and anti-vegf therapy on tumoral vessel normalization in a transgenic mouse model. Int. J. Cancer 2017, 140, 2150–2161.
- Pan, K.F.; Yang, Y.C.; Lee, W.J.; Hua, K.T.; Chien, M.H. Proteoglycan Endocan: A multifaceted therapeutic target in Cancer. Biochim. Biophys. Acta Rev. Cancer 2022, 1877, 188672.
- Matano, F.; Yoshida, D.; Ishii, Y.; Tahara, S.; Teramoto, A.; Morita, A. Endocan, a new invasion and angiogenesis marker of pituitary adenomas. J. Neuro Oncol. 2014, 117, 485–491.
- Miao, Y.; Zong, M.; Jiang, T.; Yuan, X.; Guan, S.; Wang, Y.; Zhou, D. A comparative analysis of ESM-1 and vascular endothelial cell marker (CD34/CD105) expression on pituitary adenoma invasion. Pituitary 2016, 19, 194–201.
- Cornelius, A.; Cortet-Rudelli, C.; Assaker, R.; Kerdraon, O.; Gevaert, M.H.; Prévot, V.; Lassalle, P.; Trouillas, J.; Delehedde, M.; Maurage, C.A. Endothelial expression of endocan is strongly associated with tumor progression in pituitary adenoma. Brain Pathol. 2012, 22, 757–764.
- Wang, S.; Wu, Z.; Wei, L.; Zhang, J. Endothelial cell-specific molecule-1 as an invasiveness marker for pituitary null cell adenoma. BMC Endocr. Disord. 2019, 19, 90.
- Roudnicky, F.; Poyet, C.; Wild, P.; Krampitz, S.; Negrini, F.; Huggenberger, R.; Rogler, A.; Stöhr, R.; Hartmann, A.; Provenzano, M.; et al. Endocan is upregulated on tumor vessels in invasive bladder cancer where it mediates VEGF-A-induced angiogenesis. Cancer Res. 2013, 73, 1097–1106.
- Tsai, H.F.; Chang, Y.C.; Li, C.H.; Chan, M.H.; Chen, C.L.; Tsai, W.C.; Hsiao, M. Type V collagen alpha 1 chain promotes the malignancy of glioblastoma through PPRC1-ESM1 axis activation and extracellular matrix remodeling. Cell Death Discov. 2021, 7, 313.
- Sun, L.; Sun, C.; Sun, J.; Yang, W. Downregulation of ENDOCAN in myeloid leukemia cells inhibits proliferation and promotes apoptosis by suppressing nuclear factor κB activity. Mol. Med. Rep. 2019, 19, 3247–3254.
- Vierimaa, O.; Georgitsi, M.; Lehtonen, R.; Vahteristo, P.; Kokko, A.; Raitila, A.; Tuppurainen, K.; Ebeling, T.M.; Salmela, P.I.; Paschke, R.; et al. Pituitary adenoma predisposition caused by germline mutations in the AIP gene. Science 2006, 312, 1228–1230.
- Re, A.; Ferraù, F.; Cafiero, C.; Spagnolo, F.; Barresi, V.; Romeo, D.P.; Ragonese, M.; Grassi, C.; Pontecorvi, A.; Farsetti, A.; et al. Somatic Deletion in Exon 10 of Aryl Hydrocarbon Receptor Gene in Human GH-Secreting Pituitary Tumors. Front. Endocrinol. 2020, 11, 591039.
- De Pinho, L.K.J.; Neto, L.V.; Wildemberg, L.E.A.; Gasparetto, E.L.; Marcondes, J.; de Almeida Nunes, B.; Takiya, C.M.; Gadelha, M.R. Low aryl hydrocarbon receptor-interacting protein expression is a better marker of invasiveness in somatotropinomas than Ki-67 and p53. Neuroendocrinology 2011, 94, 39–48.
- Neou, M.; Villa, C.; Armignacco, R.; Jouinot, A.; Raffin-Sanson, M.L.; Septier, A.; Letourneur, F.; Diry, S.; Diedisheim, M.; Izac, B.; et al. Pangenomic Classification of Pituitary Neuroendocrine Tumors. Cancer Cell 2020, 37, 123–134.e5.
- Sapkota, S.; Horiguchi, K.; Tosaka, M.; Yamada, S.; Yamada, M. Whole-Exome Sequencing Study of Thyrotropin-Secreting Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2017, 102, 566–575.
- Zhang, F.; Zhang, Q.; Zhu, J.; Yao, B.; Ma, C.; Qiao, N.; He, S.; Ye, Z.; Wang, Y.; Han, R.; et al. Integrated proteogenomic characterization across major histological types of pituitary neuroendocrine tumors. Cell Res. 2022, 32, 1047–1067.
- Landis, C.A.; Harsh, G.; Lyons, J.; Davis, R.L.; McCormick, F.; Bourne, H.R. Clinical characteristics of acromegalic patients whose pituitary tumors contain mutant Gs protein. J. Clin. Endocrinol. Metab. 1990, 71, 1416–1420.
- Buchfelder, M.; Fahlbusch, R.; Merz, T.; Symowski, H.; Adams, E.F. Analysis of GNAS mutations in 60 growth hormone secreting pituitary tumors: Correlation with clinical and pathological characteristics and surgical outcome based on highly sensitive GH and IGF-I criteria for remission. Pituitary 2007, 10, 275–282.
- Buchfelder, M.; Fahlbusch, R.; Merz, T.; Symowski, H.; Adams, E.F. Clinical correlates in acromegalic patients with pituitary tumors expressing GSP oncogenes. Pituitary 1999, 1, 181–185.
- Efstathiadou, Z.A.; Bargiota, A.; Chrisoulidou, A.; Kanakis, G.; Papanastasiou, L.; Theodoropoulou, A.; Tigas, S.K.; Vassiliadi, D.A.; Alevizaki, M.; Tsagarakis, S. Impact of gsp mutations in somatotroph pituitary adenomas on growth hormone response to somatostatin analogs: A meta-analysis. Pituitary 2015, 18, 861–867.
- Foltran, R.K.; Amorim, P.V.G.H.; Duarte, F.H.; Grande, I.P.P.; Freire, A.C.T.B.; Frassetto, F.P.; Dettoni, J.B.; Alves, V.A.; Castro, I.; Trarbach, E.B.; et al. Study of major genetic factors involved in pituitary tumorigenesis and their impact on clinical and biological characteristics of sporadic somatotropinomas and non-functioning pituitary adenomas. Braz. J. Med. Biol. Res. 2018, 51, e7427.
- Niendorf, S.; Oksche, A.; Kisser, A.; Löhler, J.; Prinz, M.; Schorle, H.; Feller, S.; Lewitzky, M.; Horak, I.; Knobeloch, K.P. Essential role of ubiquitin-specific protease 8 for receptor tyrosine kinase stability and endocytic trafficking in vivo. Mol. Cell. Biol. 2007, 27, 5029–5039.
- Byun, S.; Lee, S.Y.; Lee, J.; Jeong, C.H.; Farrand, L.; Lim, S.; Reddy, K.; Kim, J.Y.; Lee, M.H.; Lee, H.J.; et al. USP8 is a novel target for overcoming gefitinib resistance in lung cancer. Clin. Cancer Res. 2013, 19, 3894–3904.
- Ma, Z.Y.; Song, Z.J.; Chen, J.H.; Wang, Y.F.; Li, S.Q.; Zhou, L.F.; Mao, Y.; Li, Y.M.; Hu, R.G.; Zhang, Z.Y.; et al. Recurrent gain-of-function USP8 mutations in Cushing’s disease. Cell Res. 2015, 25, 306–317.
- Perez-Rivas, L.G.; Theodoropoulou, M.; Ferraù, F.; Nusser, C.; Kawaguchi, K.; Stratakis, C.A.; Faucz, F.R.; Wildemberg, L.E.; Assié, G.; Beschorner, R.; et al. The Gene of the Ubiquitin-Specific Protease 8 Is Frequently Mutated in Adenomas Causing Cushing’s Disease. J. Clin. Endocrinol. Metab. 2015, 100, E997–E1004.
- Albani, A.; Pérez-Rivas, L.G.; Dimopoulou, C.; Zopp, S.; Colón-Bolea, P.; Roeber, S.; Honegger, J.; Flitsch, J.; Rachinger, W.; Buchfelder, M.; et al. The USP8 mutational status may predict long-term remission in patients with Cushing’s disease. Clin. Endocrinol. 2018, 89, 454–458.
- Faucz, F.R.; Tirosh, A.; Tatsi, C.; Berthon, A.; Hernández-Ramírez, L.C.; Settas, N.; Angelousi, A.; Correa, R.; Papadakis, G.Z.; Chittiboina, P.; et al. Somatic USP8 Gene Mutations Are a Common Cause of Pediatric Cushing Disease. J. Clin. Endocrinol. Metab. 2017, 102, 2836–2843.
- Duong, C.V.; Emes, R.D.; Wessely, F.; Yacqub-Usman, K.; Clayton, R.N.; Farrell, W.E. Quantitative, genome-wide analysis of the DNA methylome in sporadic pituitary adenomas. Endocr. Relat. Cancer 2012, 19, 805–816.
- Kober, P.; Boresowicz, J.; Rusetska, N.; Maksymowicz, M.; Goryca, K.; Kunicki, J.; Bonicki, W.; Siedlecki, J.A.; Bujko, M. DNA methylation profiling in nonfunctioning pituitary adenomas. Mol. Cell Endocrinol. 2018, 473, 194–204.
- Dudley, K.J.; Revill, K.; Clayton, R.N.; Farrell, W.E. Pituitary tumors: All silent on the epigenetics front. J. Mol. Endocrinol. 2009, 42, 461–468.
- Ling, C.; Pease, M.; Shi, L.; Punj, V.; Shiroishi, M.S.; Commins, D.; Weisenberger, D.J.; Wang, K.; Zada, G. A pilot genome-scale profiling of DNA methylation in sporadic pituitary macroadenomas: Association with tumor invasion and histopathological subtype. PLoS ONE 2014, 9, e96178.
- Duan, J.; Lu, G.; Li, Y.; Zhou, S.; Zhou, D.; Tao, H. miR-137 functions as a tumor suppressor gene in pituitary adenoma by targeting AKT2. Int. J. Clin. Exp. Pathol. 2019, 12, 1557–1564.
- Lloyd, R.V.; Osamura, R.Y.; Klöppel, G.; Rosai, J. Chapter 01: Tumours of the pituitary gland. In WHO Classification of Tumours of Endocrine Organs, 4th ed.; International Agency for Research on Cancer: Lyon, France, 2017; Volume 10, pp. 12–44.
- Mete, O.; Osamura, R.Y.; Asa, L.S. Chapter 02: Pituitary Tumours. In WHO Classification of Tumours Editorial Board. Endocrine and Neuroendocrine Tumours, 5th ed.; International Agency for Research on Cancer: Lyon, France, 2022; Volume 10. Available online: https://publications.iarc.fr (accessed on 30 June 2023).
More
Information
Subjects:
Medicine, Research & Experimental
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
557
Revisions:
2 times
(View History)
Update Date:
08 Nov 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No