Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Tigran Harutyunyan | -- | 1125 | 2023-09-29 11:04:43 | | | |
| 2 | Wendy Huang | Meta information modification | 1125 | 2023-10-07 09:34:00 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Hovhannisyan, G.; Harutyunyan, T.; Aroutiounian, R.; Liehr, T. Sources and Biological Activity of Cell-Free DNA. Encyclopedia. Available online: https://encyclopedia.pub/entry/49793 (accessed on 26 July 2026).
Hovhannisyan G, Harutyunyan T, Aroutiounian R, Liehr T. Sources and Biological Activity of Cell-Free DNA. Encyclopedia. Available at: https://encyclopedia.pub/entry/49793. Accessed July 26, 2026.
Hovhannisyan, Galina, Tigran Harutyunyan, Rouben Aroutiounian, Thomas Liehr. "Sources and Biological Activity of Cell-Free DNA" Encyclopedia, https://encyclopedia.pub/entry/49793 (accessed July 26, 2026).
Hovhannisyan, G., Harutyunyan, T., Aroutiounian, R., & Liehr, T. (2023, September 29). Sources and Biological Activity of Cell-Free DNA. In Encyclopedia. https://encyclopedia.pub/entry/49793
Hovhannisyan, Galina, et al. "Sources and Biological Activity of Cell-Free DNA." Encyclopedia. Web. 29 September, 2023.
Copy Citation
The term cell-free DNA (cfDNA) encompasses all kinds of extracellular DNA molecules found in serum or plasma and other body fluids, and includes genomic DNA (gDNA) and mitochondrial DNA (mtDNA), as well as DNA of bacterial or viral origin. Cell-free DNA (cfDNA), freely circulating in the bloodstream, urine, and other fluids (or encapsulated in vesicles) may be derived from both normal and diseased cells. cfDNA is extremely dynamic and responsive, providing sensitive indicators of changes that are not detectable by standard clinical tests. It can be used as a reliable, safe, and objective tool to reflect disease progression and supplement clinical data in a particular patient, and thus, represents a new path in personalized medicine.
cfDNA
apoptosis
necrosis
NETosis
EVs
MNi
Erythroblast
1. Introduction
A pathology-related increase of cfDNA level was first detected in patients with systemic lupus erythematosus in 1966 [1] and cancer patients [2] in 1977. Moreover, Leon et al. [2] found a decrease of cfDNA level after anticancer therapy. The identification of placenta-derived cfDNA in maternal blood in 1997 [3] led to the establishment of NIPT cell-free mtDNA (cf-mtDNA) was first reported in serum and plasma of patients with type 2 diabetes mellitus and healthy donors in 2000. Hereby, an A to G 3243 mutation was identified in cf-mtDNA of diabetes patients but not in a control group [4].
Elevated levels of host cfDNA in the blood (plasma or serum) have been found in various viral diseases, as well as in COVID-19. Initially, an increase in cfDNA and its prognostic role was reported in patients with acute dengue virus infection [5][6]. An increase of cfDNA level in plasma during acute Puumala hantavirus infection, as well as its positive correlation with blood leukocyte concentration and length of hospital stay was found [7]. Elevated levels of plasma cfDNA have been found in patients with Hantaan virus infection [8]. The difference between cfDNA levels was revealed in mild, intermediate, and severely affected patients with Crimean–Congo hemorrhagic fever caused by a tick-borne virus (Nairovirus) [9]. cfDNA as a marker of COVID-19 diagnostics and management is currently under study [10]. In addition to freely circulating, cfDNA attached to extracellular vesicles, as well as cfDNA as part of neutrophil extracellular traps (NETs) has been studied in viral pathologies.
While cfDNA research and application had a slow start, at present, a rapid progress is observed, due to its great potential as a minimally invasive source of diagnostically relevant information, including patient-centered diagnosis, prognosis, prediction, and monitoring [11].
cfDNA origin in body fluids is mainly associated with cell death through apoptosis, necrosis, and NETosis. It can also be formed as a result of erythroblast enucleation and chromosomal instability. Furthermore, cfDNA can be present in its free form or encapsulated in extracellular vesicles (EVs). The main manifestations of the biological activity of cfDNA are regulation of the immune system, removal of damaged DNA, and intercellular communication.
2. Apoptosis, Necrosis and NETosis
Cell death results in massive release of cellular contents into the extracellular space. Increased cfDNA levels have been reported for various diseases, including viral infections, in which cell death and tissue/organ damage contribute to pathogenesis. Cell death pathways are initiated during viral attachment, entry, genome replication, and gene expression.
Apoptosis, as well as two forms of regulated necrosis—necroptosis and pyroptosis, are considered as the main ways of virus associated cell death [12][13]. cfDNA can also originate from neutrophil extracellular traps (NETs), web-like structures formed due to the programmed death of neutrophils called NETosis in response to infections or cancer [14]. Brinkmann et al. [15] for the first time reported that NETs are composed of decondensed chromatin complexed with neutrophil proteins (e.g., neutrophil elastase and myeloperoxidase) which are able to capture, neutralize, and kill microbes. NETs can contain both mtDNA and gDNA [16].
A significant increase in apoptotic signals was observed in postmortem lung sections from COVID-19 patients and the lungs of a non-human primate model infected with SARS-CoV-2 [17]. Apoptotic death of T-cells was demonstrated by André et al. [18] in severely affected COVID-19 patients. Necroptosis and pyroptosis are commonly seen in COVID-19 in conditions of low apoptosis [19]. However, the assessment of cell death in the mentioned studies is not accompanied by data on the release of cfDNA. Upcoming detailed analysis of released molecules could supplement the data on COVID-19 molecular pathology.
The main forms of cell death associated with COVID-19 (Figure 1) are summarized in the reviews of Rex et al. [13], Bader et al. [20], and Morais da Silva et al. [21].
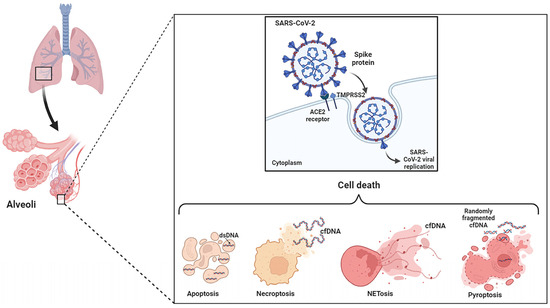
Figure 1. Main forms of cell death associated with COVID-19 and occurrence of cfDNA (This illustration was created with BioRender.com).
3. Extracellular Vesicles
EVs such as apoptotic bodies (1000–5000 nm), microvesicles (100–3000 nm), and exosomes (30–100 nm) are another source of cfDNA [22]. Thakur et al. [23] was the first to discover double-stranded gDNA (dsDNA) in exosomes from different cancer models.
Exosomes are secreted by all cell types and have been found in plasma, urine, semen, saliva, bronchial fluid, cerebrospinal fluid, breast milk, serum, amniotic fluid, synovial fluid, tears, lymph, bile, and gastric acid [22]. Whole-genome sequencing revealed that fragments of DNA in EVs (EV-DNA) can originate from any chromosome and mtDNA, which indicates a sequence-independent DNA fragment loading [24]. EVs may also contain single-stranded DNA (ssDNA) fragments [25], which can be transferred from cell to cell. Plasma cfDNA analysis in healthy donors showed that more than 93% of cfDNA is located in plasma exosomes [26]. However, cfDNA can be released into circulation through the breakdown of EVs [27].
It has been shown that cfDNA secreted in exosomes may serve to maintain tissue homeostasis by removing damaged DNA. The inhibition of exosome secretion results in the accumulation of nuclear DNA in the cytoplasm and provokes the innate immune response [28].
DNA sequences in EVs, which could be delivered from one cell to another, can regulate mRNA and protein expression, and affect the physiological function in the recipient cells [29].
4. Chromosomal Instability and Micronucleus Formation
Micronuclei (MNi) have become recognized as one of the most important biomarkers of genomic instability and source of pro-inflammatory DNA in humans [30]. Mackenzie et al. [31] reported activation of cytosolic DNA sensor cyclic GMP-AMP synthase (cGAS) that triggers a type-I interferon response in micronucleated mouse embryonic fibroblasts. The authors suggest that DNA is released from MNi due to micronuclear membrane rupture, which leads to the exposure of DNA to the cGAS. Grabuschnig et al. [25] hypothesized that MNi, with their DNA cargo, may translocate to the extracellular space and serve as sources of cfDNA; however, appropriate studies supporting this hypothesis have not yet been implemented.
5. Erythroblast Enucleation
An additional mechanism of active DNA release is nucleus exclusion from erythroblasts. Lam et al. [32] found that erythroid DNA comprised a significant proportion of the cfDNA pool in plasma of healthy individuals (about 30%). The fraction of erythroid derived cfDNA in plasma of anemia patients was 12.4%, which was significantly lower than that of the healthy controls. These findings are concordant with reduced erythropoietic activity in anemia patients.
References
- Tan, E.M.; Kunkel, H.G. Characteristics of a Soluble Nuclear Antigen Precipitating with Sera of Patients with Systemic Lupus Erythematosus. J. Immunol. 1966, 96, 464–471.
- Leon, S.A.; Ehrlich, G.E.; Shapiro, B.; Labbate, V.A. Free DNA in the Serum of Rheumatoid Arthritis Patients. J. Rheumatol. 1977, 4, 139–143.
- Dennis Lo, Y.M.; Corbetta, N.; Chamberlain, P.F.; Rai, V.; Sargent, I.L.; Redman, C.W.G.; Wainscoat, J.S. Presence of Fetal DNA in Maternal Plasma and Serum. Lancet 1997, 350, 485–487.
- Zhong, S.; Ng, M.C.Y.; Lo, Y.M.D.; Chan, J.C.N.; Johnson, P.J. Presence of Mitochondrial TRNA(Leu(UUR)) A to G 3243 Mutation in DNA Extracted from Serum and Plasma of Patients with Type 2 Diabetes Mellitus. J. Clin. Pathol. 2000, 53, 466–469.
- Ha, T.T.N.; Huy, N.T.; Murao, L.A.; Lan, N.T.P.; Thuy, T.T.; Tuan, H.M.; Nga, C.T.P.; van Tuong, V.; van Dat, T.; Kikuchi, M.; et al. Elevated Levels of Cell-Free Circulating DNA in Patients with Acute Dengue Virus Infection. PLoS ONE 2011, 6, e25969.
- Phuong, N.T.N.; Manh, D.H.; Dumre, S.P.; Mizukami, S.; Weiss, L.N.; van Thuong, N.; Ha, T.T.N.; Phuc, L.H.; van An, T.; Tieu, T.M.; et al. Plasma Cell-Free DNA: A Potential Biomarker for Early Prediction of Severe Dengue. Ann. Clin. Microbiol. Antimicrob. 2019, 18, 10.
- Outinen, T.K.; Kuparinen, T.; Jylhävä, J.; Leppänen, S.; Mustonen, J.; Mäkelä, S.; Pörsti, I.; Syrjänen, J.; Vaheri, A.; Hurme, M. Plasma Cell-Free DNA Levels Are Elevated in Acute Puumala Hantavirus Infection. PLoS ONE 2012, 7, e31455.
- Yi, J.; Zhang, Y.; Zhang, Y.; Ma, Y.; Zhang, C.; Li, Q.; Liu, B.; Liu, Z.; Liu, J.; Zhang, X.; et al. Increased Plasma Cell-Free DNA Level during HTNV Infection: Correlation with Disease Severity and Virus Load. Viruses 2014, 6, 2723–2734.
- Bakir, M.; Engin, A.; Kuskucu, M.A.; Bakir, S.; Gündag, O.; Midilli, K. Relationship of Plasma Cell-Free DNA Level with Mortality and Prognosis in Patients with Crimean-Congo Hemorrhagic Fever. J. Med. Virol. 2016, 88, 1152–1158.
- Stawski, R.; Nowak, D.; Perdas, E. Cell-Free DNA: Potential Application in COVID-19 Diagnostics and Management. Viruses 2022, 14, 321.
- Teo, Y.V.; Capri, M.; Morsiani, C.; Pizza, G.; Faria, A.M.C.; Franceschi, C.; Neretti, N. Cell-free DNA as a biomarker of aging. Aging Cell 2019, 18, e12890.
- Bai, Y.; Zheng, F.; Zhang, T.; Luo, Q.; Luo, Y.; Zhou, R.; Jin, Y.; Shan, Y.; Cheng, J.; Yang, Z.; et al. Integrating Plasma Cell-free DNA with Clinical Laboratory Results Enhances the Prediction of Critically Ill Patients with COVID-19 at Hospital Admission. Clin. Transl. Med. 2022, 12, e966.
- Danthi, P. Viruses and the Diversity of Cell Death. Annu. Rev. Virol. 2016, 3, 533–553.
- Rex, D.A.B.; Prasad, T.S.K.; Kandasamy, R.K. Revisiting Regulated Cell Death Responses in Viral Infections. Int. J. Mol. Sci. 2022, 23, 7023.
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2020, 36, 191–218.
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extracellular Traps Kill Bacteria. Science 2004, 303, 1532–1535.
- Keshari, R.S.; Jyoti, A.; Kumar, S.; Dubey, M.; Verma, A.; Srinag, B.S.; Krishnamurthy, H.; Barthwal, M.K.; Dikshit, M. Neutrophil Extracellular Traps Contain Mitochondrial as Well as Nuclear DNA and Exhibit Inflammatory Potential. Cytom. Part A 2012, 81 A, 238–247.
- Liu, Y.; Garron, T.M.; Chang, Q.; Su, Z.; Zhou, C.; Qiu, Y.; Gong, E.C.; Zheng, J.; Whitney Yin, Y.; Ksiazek, T.; et al. Cell-Type Apoptosis in Lung during Sars-Cov-2 Infection. Pathogens 2021, 10, 509.
- André, S.; Picard, M.; Cezar, R.; Roux-Dalvai, F.; Alleaume-Butaux, A.; Soundaramourty, C.; Cruz, A.S.; Mendes-Frias, A.; Gotti, C.; Leclercq, M.; et al. T Cell Apoptosis Characterizes Severe COVID-19 Disease. Cell Death Differ. 2022, 29, 1486–1499.
- Sun, C.; Han, Y.; Zhang, R.; Liu, S.; Wang, J.; Zhang, Y.; Chen, X.; Jiang, C.; Wang, J.; Fan, X.; et al. Regulated Necrosis in COVID-19: A Double-Edged Sword. Front. Immunol. 2022, 13, 917141.
- Bader, S.M.; Cooney, J.P.; Pellegrini, M.; Doerflinger, M. Programmed Cell Death: The Pathways to Severe COVID-19? Biochem. J. 2022, 479, 609–628.
- Da Silva, M.M.; de Lucena, A.S.L.; Júnior, S.d.S.L.P.; de Carvalho, V.M.F.; de Oliveira, P.S.S.; da Rosa, M.M.; Rego, M.J.B.d.M.; Pitta, M.G.d.R.; Pereira, M.C. Cell Death Mechanisms Involved in Cell Injury Caused by SARS-CoV-2. Rev. Med. Virol. 2022, 32, 3.
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727.
- Thakur, B.K.; Zhang, H.; Becker, A.; Matei, I.; Huang, Y.; Costa-Silva, B.; Zheng, Y.; Hoshino, A.; Brazier, H.; Xiang, J.; et al. Double-Stranded DNA in Exosomes: A Novel Biomarker in Cancer Detection. Cell Res. 2014, 24, 766–769.
- Grabuschnig, S.; Bronkhorst, A.J.; Holdenrieder, S.; Rodriguez, I.R.; Schliep, K.P.; Schwendenwein, D.; Ungerer, V.; Sensen, C.W. Putative Origins of Cell-Free DNA in Humans: A Review of Active and Passive Nucleic Acid Release Mechanisms. Int. J. Mol. Sci. 2020, 21, 8062.
- Lázaro-Ibáñez, E.; Lässer, C.; Shelke, G.V.; Crescitelli, R.; Jang, S.C.; Cvjetkovic, A.; García-Rodríguez, A.; Lötvall, J. DNA Analysis of Low- and High-Density Fractions Defines Heterogeneous Subpopulations of Small Extracellular Vesicles Based on Their DNA Cargo and Topology. J. Extracell. Vesicles 2019, 8, 1656993.
- Fernando, M.R.; Jiang, C.; Krzyzanowski, G.D.; Ryan, W.L. New Evidence That a Large Proportion of Human Blood Plasma Cell-Free DNA Is Localized in Exosomes. PLoS ONE 2017, 12, e0183915.
- Mondelo-Macía, P.; Castro-Santos, P.; Castillo-García, A.; Muinelo-Romay, L.; Diaz-Peña, R. Circulating Free DNA and Its Emerging Role in Autoimmune Diseases. J. Pers. Med. 2021, 11, 151.
- Takahashi, A.; Okada, R.; Nagao, K.; Kawamata, Y.; Hanyu, A.; Yoshimoto, S.; Takasugi, M.; Watanabe, S.; Kanemaki, M.T.; Obuse, C.; et al. Exosomes Maintain Cellular Homeostasis by Excreting Harmful DNA from Cells. Nat. Commun. 2017, 8, 9.
- Cai, J.; Han, Y.; Ren, H.; Chen, C.; He, D.; Zhou, L.; Eisner, G.M.; Asico, L.D.; Jose, P.A.; Zeng, C. Extracellular Vesicle-Mediated Transfer of Donor Genomic DNA to Recipient Cells Is a Novel Mechanism for Genetic Influence between Cells. J. Mol. Cell Biol. 2013, 5, 227–238.
- Fenech, M.; Knasmueller, S.; Bolognesi, C.; Holland, N.; Bonassi, S.; Kirsch-Volders, M. Micronuclei as Biomarkers of DNA Damage, Aneuploidy, Inducers of Chromosomal Hypermutation and as Sources of pro-Inflammatory DNA in Humans. Mutat. Res.-Rev. Mutat. Res. 2020, 786, 108342.
- MacKenzie, K.J.; Carroll, P.; Martin, C.A.; Murina, O.; Fluteau, A.; Simpson, D.J.; Olova, N.; Sutcliffe, H.; Rainger, J.K.; Leitch, A.; et al. CGAS Surveillance of Micronuclei Links Genome Instability to Innate Immunity. Nature 2017, 548, 461–465.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
611
Revisions:
2 times
(View History)
Update Date:
07 Oct 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No