Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Marco Zeppieri | -- | 2625 | 2023-09-01 07:03:27 | | | |
| 2 | Catherine Yang | Meta information modification | 2625 | 2023-09-01 07:15:23 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Agosti, E.; Panciani, P.P.; Zeppieri, M.; De Maria, L.; Pasqualetti, F.; Tel, A.; Zanin, L.; Fontanella, M.M.; Ius, T. Tumor Microenvironment and Glioblastoma Cell Interplay. Encyclopedia. Available online: https://encyclopedia.pub/entry/48727 (accessed on 27 June 2026).
Agosti E, Panciani PP, Zeppieri M, De Maria L, Pasqualetti F, Tel A, et al. Tumor Microenvironment and Glioblastoma Cell Interplay. Encyclopedia. Available at: https://encyclopedia.pub/entry/48727. Accessed June 27, 2026.
Agosti, Edoardo, Pier Paolo Panciani, Marco Zeppieri, Lucio De Maria, Francesco Pasqualetti, Alessandro Tel, Luca Zanin, Marco Maria Fontanella, Tamara Ius. "Tumor Microenvironment and Glioblastoma Cell Interplay" Encyclopedia, https://encyclopedia.pub/entry/48727 (accessed June 27, 2026).
Agosti, E., Panciani, P.P., Zeppieri, M., De Maria, L., Pasqualetti, F., Tel, A., Zanin, L., Fontanella, M.M., & Ius, T. (2023, September 01). Tumor Microenvironment and Glioblastoma Cell Interplay. In Encyclopedia. https://encyclopedia.pub/entry/48727
Agosti, Edoardo, et al. "Tumor Microenvironment and Glioblastoma Cell Interplay." Encyclopedia. Web. 01 September, 2023.
Copy Citation
The invasive nature of glioblastoma is problematic in a radical surgery approach and can be responsible for tumor recurrence. In order to create new therapeutic strategies, it is imperative to have a better understanding of the mechanisms behind tumor growth and invasion. The continuous cross-talk between glioma stem cells (GSCs) and the tumor microenvironment (TME) contributes to disease progression, which renders research in this field difficult and challenging.
tumor microenvironment
stem cells
tumor-associated macrophages
1. Introduction
Glioblastoma (GBM) is the most prevalent brain tumor in adults, and despite multimodality therapy, including maximal safe resection surgery, chemotherapy (CTX), and radiotherapy (RT), the prognosis tends to be rather unfavorable with an average overall survival (OS) typically less than 20 months [1][2][3][4]. The tumor’s site and genetic and molecular profiles, such as IDH1 mutation, p53 mutation, 1p/19q codeletion, EGFR amplification, MGMT methylation, and ATRX mutation, play a crucial role in defining its prognosis. [3][4][5][6].
A phase III trial study by Stupp et al. in 2005 [1] reported the important role of TMZ with concurrent RT followed by adjuvant treatment with TMZ in the treatment of newly diagnosed GBM, and no other CT drug has demonstrated notably better results. Additionally, although bevacizumab has U.S. Food Drug Administration (FDA) approval for recurrent GBM (rGBM), no phase III clinical trials have proved its OS benefits. Consequently, no cure for GBM exists. Elevated dismal outcomes are primarily due to the tumor’s high recurrence rate, which is closely tied to its resistance to standard therapies. Studies have proposed different therapeutic alternatives in the management of patients with GBM, however, a gold standard approach is still lacking [3][6][7].
Recurrence of the tumor in GBM individuals is one of the main causes of mortality, which could be due to the presence and persistence of cancer cells with stem cell characteristics [3][6][7][8][9]. Over the past ten years, researchers have successfully isolated glioblastoma stem cells (GSCs) from GBM and discovered their critical role in the development, maintenance, and recurrence of tumors. As a result, GSCs have become a crucial target for new treatments due to their ability to resist standard therapies and contribute to malignant relapse [3][6][10][11].
An increased number of investigations have demonstrated that subpopulations of tumor stem cells can favor resistance to therapies and cause malignant relapse, due to their capabilities of differentiation, self-renewal, growth, and progression [3][6][8][9]. The resistance mechanisms of GSCs include their quiescence, higher mitochondrial reserve, extensive DNA repair capabilities, and location in hypoxic niches [11][12][13][14][15]. Simultaneously, numerous studies have shown that tumor microenvironment (TME) and its interplay with GBM stem cells, through the release of extracellular vesicles, is gaining an important role in GBM pathogenesis and proliferation characteristics. TME has been proposed as a promising therapeutic target [9][12][16][17][18].
The GBM microenvironment is a dynamic entity comprised of several different types of cells and factors, which include tumor cells, GSCs, endothelial cells, astrocytes, microglia, stromal components, soluble factors, extracellular matrix, and other substances [9][18]. Each component plays an important and specific role in the microenvironment. It has been shown that GSCs, for example, can establish a continuous cross-talk that actively assists in the remodeling of the microenvironment. These factors can be influential in maintaining and contributing to the progression of the disease [6][8][9][11][12]. Identifying the key actors in this scenario has become increasingly challenging, as the dialogue between GSCs and the microenvironment plays a significant role in supporting GBM [9][10][11][12][13][14][15][16]. The TME also includes immune and myeloid cells. Myeloid cells are the most frequent and, among them, tumor-associated macrophages (TAMs) are the most abundant [3][6][14][17][19][20].
Characterization of chemoresistance and radioresistance has traditionally been focused on the effects of TMZ and RT on the tumor cells, while overlooking the impact on the TME. Although components of the TME have been shown to regulate cell proliferation and migration, stemness maintenance, angiogenesis, and malignant progression, their role concerning the response to TMZ and RT in GBM has been less analyzed [5][9][15][16][20][21][22][23][24].
2. Tumor Microenvironment and Glioblastoma Cell Interplay as Promoters of Therapeutic Resistance
TAMs are the major biological constituents of the GBM TME and are implicated in GBM progression, angiogenesis, and, according to recent reports, the development of resistance to adjuvant treatments [3][5][6]. GBM cells and TAMs are constantly in close cross-talk, communicating through paracrine signals, including cytokines and the extracellular production and release of exosomes, which are a novel class of extracellular vesicles that have gained enormous attention lately as facilitators of the progression of various tumors [18]. Exosomes are secreted by different cells in the body, including tumor cells and other cells in the tumor microenvironment, and are mainly used as vehicles to exchange information between cells. Their content in terms of nucleic acids (mainly DNA, mRNA, and miRNA) is dynamic and related to the cells of origin. Since each miRNA is responsible for the regulation of several mRNAs, the transport of different miRNAs from one cell to another via exosomes involves the regulation of several mRNAs in the acceptor cell and, as the final outcome, modifies its behavior. The exchange of information that follows the release of exosomes is capable of mutually modifying the phenotype of tumor cells and those in the tumor microenvironment. miRNAs and lncRNA can be transmitted via exosomes from cancer cells to other cells in the TME and vice versa, influencing tumor development, aggressiveness, and progression and TAM immunosuppressive polarization [18][21][22][23][24][25][26].
It has been currently postulated that TMZ resistance in GBM can be reflected by extracellular vesicles. Yin et al. [15] found that high serum levels of exosomes from GBM patients containing miRNA-1238 are indicative of TMZ resistance of the GBM cells. Chuang et al. [27] provided evidence that miRNA-21 and miR-416a enriched TAM M2-derived exosomes, contributing to GBM malignancy via increasing stemness and favoring the development of TMZ resistance. Witusik-Perkowska et al. [28] showed that miRNA-31, miRNA-221, miRNA-222, and miRNA-21 are expressed at higher levels in the serum of GBM patients with TMZ resistance. These miRNAs, downregulating PTEN expression in tumor cells, may reduce the sensitivity of GBM cells to TMZ. All this evidence leads to the assumption that specific miRNAs’ exosomal profiles (miRNA-31, miRNA-21, miRNA-221, miRNA-416a, miRNA-222, and miRNA-1238) in GBM patients’ serum could be considered as a possible biomarker in developing CTX protocols. Additional clinical data and studies, however, are needed to develop efficient, reliable, cost-effective, and convenient methods for identifying circulating miRNAs [15][18][21][22][23][24][25][26][27][28].
The role of miRNAs in favoring chemoresistance has been documented in current literature, however, there is less evidence regarding the role of lncRNAs. Wu et al. [29] demonstrated that lnc-RNA TALC could regulate M2 polarization and promote TMZ resistance in GBM by activating the p38 MAPK signaling pathway and promoting C5a release. In addition, Zhang et al. [23] demonstrated that exosome-mediated transfer of lncRNA 226 SBF2-AS1 from TAMs spreads TMZ resistance in GBM cells through a mechanism of upregulation of serum SBF2-AS1 levels, mediated by the transcription factor ZEB1, which binds directly to the SBF2-AS1 promoter region. Dai et al. [13] and Zheng et al. [30] showed that AHIF and lncRNAs linc-RA1 may contribute to GBM radioresistance. In detail, AHIF mediated radioresistance through VEGF-A and angiogenin in secreted exosomes [13]. On the other hand, lincRA1 stabilized the level of H2B K120 monoubiquitination (H2Bub1) by combining with H2B and inhibiting the interaction between H2Bub1 and ubiquitin-specific protease 44 (USP44), which inhibited autophagy, thus contributing to GBM radioresistance [30].
By the same mechanism, lnc-TALC also appears to favor the development of GBM radioresistance. The lncRNAs do not seem to play a role only in the development of TMZ resistance; they may also contribute to the development of GBM radioresistance. Zhang et al. [23] demonstrated that exosome-mediated transfer of lncRNA SBF2-AS1 from TAMs spreads TMZ resistance in GBM cells, while Dai et al. [13] and Zheng et al. [30] showed that lncRNAs linc-RA1 and AHIF may contribute to GBM radioresistance. This evidence suggests that the blocking of lncRNA-mediated cross-talk between GBM cells and TMAs might be a novel therapeutic strategy to address TMZ resistance and radioresistance. Moreover, specific lncRNA identifiers of an aggressive TME, predisposing to the development of TMZ resistance, can be searched for in the blood, as can miRNAs, as a biomarker of diagnosis of GBM and prognostic factor of adjuvant resistance therapies [13][23][29][30].
TAMs can be categorized into two subtypes based on their function: M1 and M2 polarized macrophages. Studies have linked M2 macrophages to increased GBM aggressiveness by secreting various molecules, such as PDGF, EGF, TGF- β1, and VEGF, to surrounding cells. However, as shown more recently, M2 macrophages may also promote the TMZ resistance and radioresistance of GBM cells [8][11][16][26][31][32]. Recent data suggest that M2 macrophages may play an important role in regulating TMZ resistance. Zhang et al. [5] and Azambuja et al. [33] showed that VEGF-dependent M2 macrophages can activate the PI3K/Akt/Nrf2 pathway to favor TMZ resistance in individuals with GBM. These results agree with previous reports that showed that upregulated VEGF levels can be important in TMZ resistance. Chuang et al. [27] showed that eliminating M2 macrophage-derived miRNA-21 exosomes can overcome GB’s TMZ resistance. M2 macrophages have been shown to have the typical surface biomarker CD163. Miyazaki et al. reported that CD163-positive M2 macrophages could also play a role in the increased immune therapy resistance in GBM cells that show TMZ resistance [34].
Accordingly, M2-polarized macrophages promote cancer stemness and chemoresistance in other types of tumors, including pancreatic cancer and thyroid cancer. Thus, in light of the numerous data reported in the literature, it is reasonable to speculate that M2 macrophages may contribute to cancer stemness, progression, aggressiveness, and TMZ resistance in GBM [35][36].
M2 macrophages appear to have a pivotal role also in the development of GBM radioresistance. Jang et al. [37] reported that the M1/M2 macrophage ratio in the TME and radiosensitivity of GBM cells are inversely associated. In this regard, short-term relapse GBMs had a significantly higher fraction of M2 macrophages after RT compared with the long-term relapse tumors detected, suggesting that M2 macrophages may play a role in radioresistance and then develop an early relapse. Furthermore, several studies have suggested that immunotherapy targeting M2 macrophages may favor the radiosensitivity of GBM cells [33][38]. These first findings highlight the primary role of M2 macrophages in the development of GBM radioresistance. The increase of the M1/M2 ratio by conversion of M2 macrophages into M1 macrophages or the destruction of M2 macrophages may, thus, represent a potential therapeutic approach for increasing GBM radiosensitivity [26][31][32][33][34][39][40].
To summarize, a high proportion of M2 macrophages’ and specific miRNAs’ (including miR-1238, miRNA-31, miRNA-221, miRNA-222, miRNA-416a, and miRNA-21) and lncRNAs (lnc-TALC, SBF2-AS1, linc-RA1, and AHIF) profiles in the TME may play a pivotal role in the acquisition of TMZ resistance and radioresistance of GBM cells. Exosomal miRNA and lncRNA levels in human serum may function not only as a potential diagnostic biomarker in GBM patients, but also as a prognostic factor for the early identification of TMZ resistant and radioresistant GBMs [31][33][34][39].
Macrophage-targeting immunotherapy has been proposed as a potential tool to improve GBM treatment strategies by preventing TAMs recruitment, repolarizing TAMs, and using immune checkpoint blockade [41]. M2-like phenotype TAMs tend to be important in the immunosuppressive TME, which can secrete immunosuppressive substances like TGF-b, IL-6, and IL-10 in GBM. The secreted pro-inflammatory cytokines include IL-12, IL-2, IFN-g, and TNF-a, which can be detected in low levels [42][43]. The M1 to M2 phenotype transition in TAMs tends to be associated with the progression of the tumor. TAMs have been shown to be important in the progression of GBM by pro-tumorigenic activities, which include GBM cell migration, proliferation, and invasion. TAMs have also been shown to play an important role in the activation of angiogenesis, generating immunosuppressive TME, and facilitating the degradation of the extracellular matrix (Figure 1) [44][45].
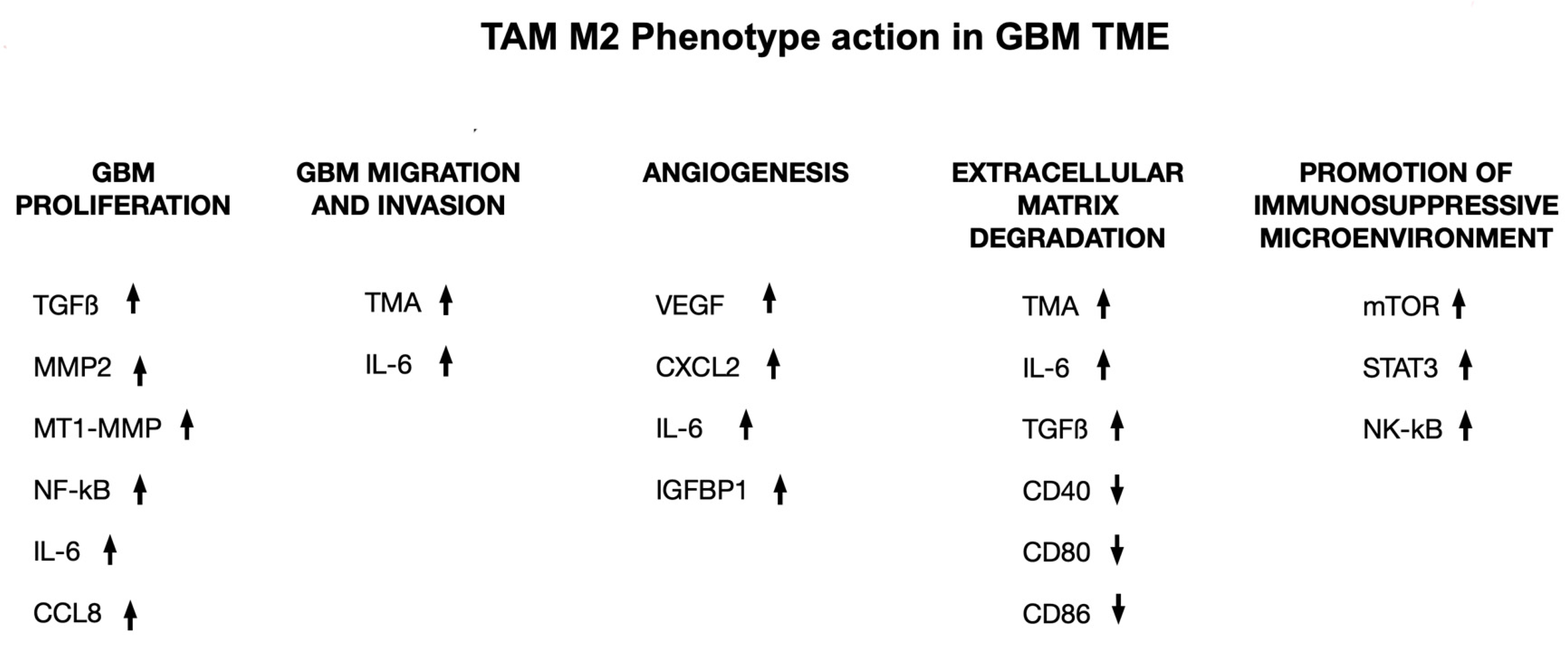
Figure 1. The transition of TAMs from M1 to M2 phenotype is linked to the advancement of tumors. TAMs are thought to drive the progression of GBM through various cytokines and factors, promoting proliferation of GBM cells, migration and invasion of GBM cells, angiogenesis within GBM, breakdown of the extracellular matrix (ECM), and an immunosuppressive TME. ↑ = increase in production; ↓ = reduction of production.
Several TAM substances, like stress-inducible protein (STI)-1, TGF-β, IL-1β, IL-6, and EGF, have been shown to enhance GBM cell invasion. Recent studies have shown that M2 macrophages play a role in promoting the vascularization of brain tumors by secreting pro-angiogenic factors like VEGF. As a result, TAM pathways have emerged as promising targets for immunotherapy. In vitro and in vivo analyses have shown that targeting these pathways can lead to reduced myeloid infiltrates, decreased tumor vascularity, and improved overall survival [45][46].
Accordingly, over the years, macrophage-targeting immunotherapy has been advocated for as a potential tool to improve GBM treatment strategies. In detail, three of the main macrophage-targeting immunotherapy strategies include: TAMs recruitment prevention, TAMs repolarization, and immune checkpoint blockade.
An effective strategy for treating GBM may involve preventing the recruitment of M2 macrophages to the GBM site, considering their role in tumor invasion and progression. Ongoing clinical trials are targeting this approach with inhibitors for the following targets: angiopoietin-2 (ANG2), CXCR4, and colony-stimulating factor 1 receptor (CSF1R), [47][48][49][50][51]. Another potential strategy is promoting a shift between M2 and M1 macrophages. CD40 and TLR agonists are currently being tested in GBM clinical trials as potential therapeutic agents targeting TAM repolarization [52][53][54][55]. In addition, immune checkpoint inhibitors such as anti-PD-1 and anti-CTLA-4 have shown success in treating melanoma and non-small cell lung cancer in recent years. Therefore, immune checkpoint blockade is a promising treatment that is also implicated in clinical trials for GBM (Table 1) [56][57].
Table 1. Review of clinical trials on immunotherapy targeting GAM in GBM (from ClinicalTrials.gov).
| Category of Immunotherapeutic Agent |
Drug Tested | Combined Therapies | Clinical Trials Registry Identifier |
Trial Phase | Patients (n) | Reported Biological Response |
|---|---|---|---|---|---|---|
| CSF1R Inhibitors | Pexidartinib | / | NCT01349036 | II | 38 | Monocyte ↓ |
| ANG2 Inhibitors | MEDI3617 | Bevacizumab | NCT01248949 | I | 13 | ORR: 0% |
| Trebananib | Bevacizumab | NCT01609790 | II | 130 | NR | |
| CXCR4 Inhibitors | Plerixafor | TMZ + RT | NCT01977677 | I/II | 29 | CXCL12 ↑ |
| Plerixafor | Bevacizumab | NCT01339039 | I | 26 | Lymphocytes ↑, Monocytes ↑ CXCL12 ↑, ANG2 ↓, sMET ↓, IL-8 ↓ | |
| CD40 Agonists | APX005M | / | NCT03389802 | I | 45 | NR |
| 2141-V11 | D2C7-IT | NCT04547777 | I | 30 | NR | |
| TLR Agonists | Poly-ICLC (TLR3 agonist) | / | NCT01188096 | II | 47 | 50% LGG respond, 25% HGG respond |
| RT | NCT00052715 | II | 31 | NR | ||
| TMZ + RT | NCT00262730 | II | 97 | NR | ||
| GAA/TT-Peptide Vaccine | NCT00795457 | I | 13 | 91% respond | ||
| Peptide Vaccines | NCT00874861 | I | 10 | 55% respond | ||
| IMA950 Peptide Vaccine | NCT01920191 | I/II | 19 | NR | ||
| Dendritic Cell Vaccine | NCT00068510 | I | 28 | TNF-a ↑, IL-6 ↑, Lymphocytes ↑ |
||
| HSPPC-96 (TLR4 agonist) |
/ | NCT02122822 | I | 20 | NR | |
| / | NCT00293423 | II | 96 | NR | ||
| CpG-ODN (TLR9 agonist) |
/ | NCT00190424 | II | 34 | No benefit | |
| PD-1 Inhibitors | Nivolumab | / | NCT02017717 | III | 529 | ORR: 7.8% |
| / | NCT02550249 | II | 29 | CXCL10 ↑, CCL4 ↑, CCL3L1 ↑ | ||
| Ipilimumab | NCT03233152 | I | 27 | No benefit | ||
| Lirilumab | NCT02813135 | II | 397 | NR | ||
| Cemiplimab | Veledimex | NCT04006119 | 40 | NR | ||
| Pembrolizumab | Bevacizumab + RT | NCT02313272 | 32 | NR | ||
| Bevacizumab | NCT02337491 | 80 | ORR:20% |
Abbreviations: ORR, objective response rate; NR, not reported; RT, radiotherapy; TMZ, temozolomide. ↑ = increase in production; ↓ = reduction of production.
3. Future Perspectives
GAMs are associated with the radio-obstruction of glioblastoma by discharging TNF α, which increments the atomic element κB (NF-κB) that is connected with substandard endurance. Different techniques have been considered to lessen the enlistment of GAMs in GBM movement, including adjusting the GAMs aggregate towards an enemy of growth M1-like microenvironment, or diminishing the M2-like cancer-advancing microenvironment (11). For GBM treatment, TLRs (toll-like receptors) could be new targets. They suppress pro-tumorigenic pathways by modulating immune responses in GBM and the innate immune system. Hence, they could address a possible new objective in GBM treatment. Polyinosinic-polycytidylic acid and poly-L-lysine (Poly ICLC—Hiltonol), a TLR3 agonist, can activate immune cells and encourage their migration into the tumor mass. In a stage I preliminary, recently analyzed GBM patients were given Poly ICLC in blend with standard consideration and customized peptides, in light of individual cancer transformations, as a growth explicit antigen immunization. Other immunostimulants, such as the TLR3 agonist Poly I, were also tested with Poly ICLC: C and TLR7/8 agonist Imiquimod in malignant growth antibody treatments on patients who went through complete growth resection of GBMs. These treatments increased OS and progression-free survival (PFS), indicating potential for future treatments [58].
References
- Stupp, R.; Mason, W.P.; van den Bent, M.J.; Weller, M.; Fisher, B.; Taphoorn, M.J.B.; Belanger, K.; Brandes, A.A.; Marosi, C.; Bogdahn, U.; et al. European Organisation for Research and Treatment of Cancer Brain Tumor and Radiotherapy Groups, National Cancer Institute of Canada Clinical Trials Group. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N. Engl. J. Med. 2005, 352, 987–996.
- Ius, T.; Pignotti, F.; Della Pepa, G.M.; La Rocca, G.; Somma, T.; Isola, M.; Battistella, C.; Gaudino, S.; Polano, M.; Dal Bo, M.; et al. A Novel Comprehensive Clinical Stratification Model to Refine Prognosis of Glioblastoma Patients Undergoing Surgical Resection. Cancers 2020, 12, 386.
- Menna, G.; Mattogno, P.P.; Donzelli, C.M.; Lisi, L.; Olivi, A.; Della Pepa, G.M. Glioma-Associated Microglia Characterization in the Glioblastoma Microenvironment through A ‘Seed-and Soil’ Approach: A Systematic Review. Brain Sci. 2022, 12, 718.
- Ius, T.; Somma, T.; Altieri, R.; Angileri, F.F.; Barbagallo, G.M.; Cappabianca, P.; Certo, F.; Cofano, F.; D’Elia, A.; Della Pepa, G.M.; et al. Is age an additional factor in the treatment of elderly patients with glioblastoma? A new stratification model: An Italian Multicenter Study. Neurosurg. Focus 2020, 49, E13.
- Zhang, G.; Tao, X.; Ji, B.; Gong, J. Hypoxia-Driven M2-Polarized Macrophages Facilitate Cancer Aggressiveness and Temozolomide Resistance in Glioblastoma. Oxid. Med. Cell. Longev. 2022, 2022, 1614336.
- Ou, A.; Yung, W.K.A.; Majd, N. Molecular mechanisms of drug resistance in glioblastoma. Int. J. Mol. Sci. 2020, 22, 6385.
- Fidoamore, A.; Cristiano, L.; Antonosante, A.; D’Angelo, M.; Di Giacomo, E.; Astarita, C.; Giordano, A.; Ippoliti, R.; Benedetti, E.; Cimini, A. Glioblastoma Stem Cells Microenvironment: The Paracrine Roles of the Niche in Drug and Radioresistance. Stem Cells Int. 2016, 2016, 6809105.
- Kenig, S.; Faoro, V.; Bourkoula, E.; Podergajs, N.; Ius, T.; Vindigni, M.; Skrap, M.; Lah, T.; Cesselli, D.; Storici, P.; et al. Topoisomerase IIβ mediates the resistance of glioblastoma stem cells to replication stress-inducing drugs. Cancer Cell Int. 2016, 16, 58.
- Manini, I.; Ruaro, M.E.; Sgarra, R.; Bartolini, A.; Caponnetto, F.; Ius, T.; Skrap, M.; Di Loreto, C.; Beltrami, A.P.; Manfioletti, G.; et al. Semaphorin-7A on Exosomes: A Promigratory Signal in the Glioma Microenvironment. Cancers 2019, 11, 758.
- Tang, X.; Zuo, C.; Fang, P.; Liu, G.; Qiu, Y.; Huang, Y.; Tang, R. Targeting Glioblastoma Stem Cells: A Review on Biomarkers, Signal Pathways and Targeted Therapy. Front. Oncol. 2021, 11, 701291.
- Andersen, R.S.; Anand, A.; Harwood, D.S.L.; Kristensen, B.W. Tumor-associated microglia and macrophages in the glioblastoma microenvironment and their implications for therapy. Cancers 2021, 13, 4255.
- Jarosz-Biej, M.; Smolarczyk, R.; Cichoń, T.; Kułach, N. Tumor microenvironment as a “game changer” in cancer radiotherapy. Int. J. Mol. Sci. 2019, 20, 3212.
- Dai, X.; Liao, K.; Zhuang, Z.; Chen, B.; Zhou, Z.; Zhou, S.; Lin, G.; Zhang, F.; Lin, Y.; Miao, Y.; et al. AHIF promotes glioblastoma progression and radioresistance via exosomes. Int. J. Oncol. 2019, 54, 261–270.
- Hide, T.; Komohara, Y.; Miyasato, Y.; Nakamura, H.; Makino, K.; Takeya, M.; Kuratsu, J.-I.; Mukasa, A.; Yano, S. Oligodendrocyte Progenitor Cells and Macrophages/Microglia Produce Glioma Stem Cell Niches at the Tumor Border. eBioMedicine 2018, 30, 94–104.
- Yin, J.; Zeng, A.; Zhang, Z.; Shi, Z.; Yan, W.; You, Y. Exosomal transfer of miR-1238 contributes to temozolomide-resistance in glioblastoma. eBioMedicine 2019, 42, 238–251.
- Caponnetto, F.; Dalla, E.; Mangoni, D.; Piazza, S.; Radovic, S.; Ius, T.; Skrap, M.; Di Loreto, C.; Beltrami, A.P.; Manini, I.; et al. The miRNA Content of Exosomes Released from the Glioma Microenvironment Can Affect Malignant Progression. Biomedicines 2020, 8, 564.
- Byrne, N.M.; Tambe, P.; Coulter, J.A. Radiation response in the tumour microenvironment: Predictive biomarkers and future perspectives. J. Pers. Med. 2021, 11, 53.
- Li, I.; Nabet, B.Y. Exosomes in the tumor microenvironment as mediators of cancer therapy resistance. Mol. Cancer 2019, 18, 32.
- Okkenhaug, K.; Graupera, M.; Vanhaesebroeck, B. Targeting PI3K in Cancer: Impact on Tumor Cells, their Protective Stroma, Angiogenesis, and Immunotherapy. Cancer Discov. 2016, 6, 1090–1105.
- Liu, X.; Huang, Y.; Qi, Y.; Wu, S.; Hu, F.; Wang, J.; Shu, K.; Zhang, H.; Bartsch, J.W.; Nimsky, C.; et al. The GBM Tumor Microenvironment as a Modulator of Therapy Response: ADAM8 Causes Tumor Infiltration of Tams through HB-EGF/EGFR-Mediated CCL2 Expression and Overcomes TMZ Chemosensitization in Glioblastoma. Cancers 2022, 14, 4910.
- Jamal, M.; Rath, B.H.; Williams, E.S.; Camphausen, K.; Tofilon, P.J. Microenvironmental regulation of glioblastoma radioresponse. Clin. Cancer Res. 2010, 16, 6049–6059.
- Hsieh, C.H.; Wu, C.P.; Lee, H.T.; Liang, J.A.; Yu, C.Y.; Lin, Y.J. NADPH oxidase subunit 4 mediates cycling hypoxia-promoted radiation resistance in glioblastoma multiforme. Free. Radic. Biol. Med. 2012, 53, 649–658.
- Zhang, Z.; Yin, J.; Lu, C.; Wei, Y.; Zeng, A.; You, Y. Exosomal transfer of long non-coding RNA SBF2-AS1 enhances chemoresistance to temozolomide in glioblastoma. J. Exp. Clin. Cancer Res. 2019, 38, 166.
- Xu, J.; Zhang, J.; Zhang, Z.; Gao, Z.; Qi, Y.; Qiu, W.; Pan, Z.; Guo, Q.; Li, B.; Zhao, S.; et al. Hypoxic glioma-derived exosomes promote M2-like macrophage polarization by enhancing autophagy induction. Cell Death Dis. 2021, 12, 373.
- Pessina, S.; Cantini, G.; Kapetis, D.; Cazzato, E.; Di Ianni, N.; Finocchiaro, G.; Pellegatta, S. The multidrug-resistance transporter Abcc3 protects NK cells from chemotherapy in a murine model of malignant glioma. Oncoimmunology 2016, 5, e1108513.
- Ansems, M.; Span, P.N. The tumor microenvironment and radiotherapy response; a central role for cancer-associated fibroblasts. Clin. Transl. Radiat. Oncol. 2020, 22, 90–97.
- Chuang, H.-Y.; Su, Y.-K.; Liu, H.-W.; Chen, C.-H.; Chiu, S.-C.; Cho, D.-Y.; Lin, S.-Z.; Chen, Y.-S.; Lin, C.-M. Preclinical evidence of STAT3 inhibitor pacritinib overcoming temozolomide resistance via downregulating miR-21-enriched exosomes from M2 glioblastoma-associated macrophages. J. Clin. Med. 2019, 8, 959.
- Witusik-Perkowska, M.; Zakrzewska, M.; Jaskolski, D.J.; Liberski, P.P.; Szemraj, J. Artificial microenvironment of in vitro glioblastoma cell cultures changes profile of miRNAs related to tumor drug resistance. OncoTargets Ther. 2019, 12, 3905–3918.
- Wu, P.; Cai, J.; Chen, Q.; Han, B.; Meng, X.; Li, Y.; Li, Z.; Wang, R.; Lin, L.; Duan, C.; et al. Lnc-TALC promotes O6-methylguanine-DNA methyltransferase expression via regulating the c-Met pathway by competitively binding with miR-20b-3p. Nat. Commun. 2019, 10, 2045.
- Zheng, J.; Wang, B.; Zheng, R.; Zhang, J.; Huang, C.; Zheng, R.; Huang, Z.; Qiu, W.; Liu, M.; Yang, K.; et al. Linc-RA1 inhibits autophagy and promotes radioresistance by preventing H2Bub1/USP44 combination in glioma cells. Cell Death Dis. 2020, 11, 758.
- Mannino, M.; Chalmers, A.J. Radioresistance of glioma stem cells: Intrinsic characteristic or property of the “microenvironment-stem cell unit”? Mol. Oncol. 2011, 5, 374–386.
- Han, X.; Xue, X.; Zhou, H.; Zhang, G. A molecular view of the radioresistance of gliomas. Oncotarget 2017, 8, 100931–100941.
- Azambuja, J.H.; da Silveira, E.F.; de Carvalho, T.R.; Oliveira, P.S.; Pacheco, S.; Couto, C.T.D.; Beira, F.T.; Stefanello, F.M.; Spanevello, R.M.; Braganhol, E. Glioma sensitive or chemoresistant to temozolomide differentially modulate macrophage protumor activities. Biochim. Biophys. Acta-Gen. Subj. 2017, 1861, 2652–2662.
- Miyazaki, T.; Ishikawa, E.; Matsuda, M.; Sugii, N.; Kohzuki, H.; Akutsu, H.; Sakamoto, N.; Takano, S.; Matsumura, A. Infiltration of CD163-positive macrophages in glioma tissues after treatment with anti-PD-L1 antibody and role of PI3Kγ inhibitor as a combination therapy with anti-PD-L1 antibody in in vivo model using temozolomide-resistant murine glioma-initiating cells. Brain Tumor Pathol. 2020, 37, 41–49.
- Pustchi, S.E.; Avci, N.G.; Akay, Y.M.; Akay, M. Astrocytes decreased the sensitivity of glioblastoma cells to temozolomide and bay 11-7082. Int. J. Mol. Sci. 2020, 21, 7154.
- Xue, B.; Xiang, W.; Zhang, Q.; Wang, H.; Zhou, Y.; Tian, H.; Abdelmaksou, A.; Xue, J.; Sun, M.; Yi, D.; et al. CD90low glioma-associated mesenchymal stromal/stem cells promote temozolomide resistance by activating FOXS1-mediated epithelial-mesenchymal transition in glioma cells. Stem Cell Res. Ther. 2021, 12, 394.
- Jang, B.S.; Kim, I.A. Relationship between Macrophage and Radiosensitivity in Human Primary and Recurrent Glioblastoma: In Silico Analysis with Publicly Available Datasets. Biomedicines 2022, 10, 292.
- Li, Z.; Meng, X.; Wu, P.; Zha, C.; Han, B.; Li, L.; Sun, N.; Qi, T.; Qin, J.; Zhang, Y.; et al. Glioblastoma Cell-Derived lncRNA-Containing Exosomes Induce Microglia to Produce Complement C5, Promoting Chemotherapy Resistance. Cancer Immunol. Res. 2021, 9, 1383–1399.
- Zhou, F.; Shi, Q.; Fan, X.; Yu, R.; Wu, Z.; Wang, B.; Tian, W.; Yu, T.; Pan, M.; You, Y.; et al. Diverse Macrophages Constituted the Glioma Microenvironment and Influenced by PTEN Status. Front. Immunol. 2022, 13, 841404.
- Alhaddad, L.; Osipov, A.N.; Leonov, S. The Molecular and Cellular Strategies of Glioblastoma and Non-Small-Cell Lung Cancer Cells Conferring Radioresistance. Int. J. Mol. Sci. 2022, 23, 13577.
- Locarno, C.V.; Simonelli, M.; Carenza, C.; Capucetti, A.; Stanzani, E.; Lorenzi, E.; Persico, P.; Della Bella, S.; Passoni, L.; Mavilio, D.; et al. Role of myeloid cells in the immunosuppressive microenvironment in gliomas. Immunobiology 2020, 225, 151853.
- Perng, P.; Lim, M. Immunosuppressive mechanisms of malignant gliomas: Parallels at non-CNS sites. Front. Oncol. 2015, 5, 153.
- Paulus, W.; Baur, I.; Huettner, C.; Schmausser, B.; Roggendorf, W.; Schlingensiepen, K.H.; Brysch, W.; SchmauBer, B. Effects of transforming growth factor-beta 1 on collagen synthesis, integrin expression, adhesion and invasion of glioma cells. J. Neuropathol. Exp. Neurol. 1995, 54, 236–244.
- Quail, D.F.; Joyce, J.A. The microenvironmental landscape of brain tumors. Cancer Cell 2017, 31, 326–341.
- Wang, G.; Zhong, K.; Wang, Z.; Zhang, Z.; Tang, X.; Tong, A.; Zhou, L. Tumor-associated microglia and macrophages in glioblastoma:From basic insights to therapeutic opportunities. Front. Immunol. 2022, 13, 964898.
- Bianconi, A.; Aruta, G.; Rizzo, F.; Salvati, L.F.; Zeppa, P.; Garbossa, D.; Cofano, F. Systematic Review on Tumor Microenvironment in Glial Neoplasm: From Understanding Pathogenesis to Future Therapeutic Perspectives. Int. J. Mol. Sci. 2022, 23, 4166.
- Butowski, N.; Colman, H.; De Groot, J.F.; Omuro, A.M.; Nayak, L.; Wen, P.Y.; Cloughesy, T.F.; Marimuthu, A.; Haidar, S.; Perry, A.; et al. Orally administered colony stimulating factor 1 receptor inhibitor Plx3397 in recurrent glioblastoma: An ivy foundation early phase clinical trials consortium phase ii study. Neuro-Oncol. 2016, 18, 557–564.
- Chen, L.; Li, J.; Wang, F.; Dai, C.; Wu, F.; Liu, X.; Li, T.; Glauben, R.; Zhang, Y.; Nie, G.; et al. Tie2 expression on macrophages is required for blood vessel reconstruction and tumor relapse after chemotherapy. Cancer Res. 2016, 76, 6828–6838.
- Hyman, D.M.; Rizvi, N.A.; Natale, R.B.; Armstrong, D.K.; Birrer, M.J.; Recht, L.; Dotan, E.; Makker, V.; Kaley, T.J.; Kuruvilla, D.; et al. Phase I study of Medi3617, a selective angiopoietin-2 inhibitor alone and combined with Carboplatin/Paclitaxel, paclitaxel, or bevacizumab for advanced solid tumors. Clin. Cancer Res. 2018, 24, 2749–2757.
- Martin-Liberal, J.; Hollebecque, A.; Aftimos, P.; Jungels, C.; Martin-Romano, P.; Rodon, J.; Kremer, J.D.; Zhang, W.; Bendell, J. First-in-Human, dose-escalation, phase 1 study of anti- Angiopoietin-2 Ly3127804 as monotherapy and in combination with ramucirumab in patients with advanced solid tumours. Br. J. Cancer 2020, 123, 1235–1243.
- Thomas, R.P.; Nagpal, S.; Iv, M.; Soltys, S.G.; Bertrand, S.; Pelpola, J.S.; Ball, R.; Yang, J.; Sundaram, V.; Chernikova, S.B.; et al. Macrophage exclusion after radiation therapy (Mert): A first in human phase I/Ii trial using a Cxcr4 inhibitor in glioblastoma. Clin. Cancer Res. 2019, 25, 6948–6957.
- van Hooren, L.; Vaccaro, A.; Ramachandran, M.; Vazaios, K.; Libard, S.; van de Walle, T.; Georganaki, M.; Huang, H.; Pietilä, I.; Lau, J.; et al. Agonistic Cd40 therapy induces tertiary lymphoid structures but impairs responses to checkpoint blockade in glioma. Nat. Commun. 2021, 12, 4127.
- Vonderheide, R.H. Cd40 agonist antibodies in cancer immunotherapy. Annu. Rev. Med. 2020, 71, 47–58.
- Deng, S.; Zhu, S.; Qiao, Y.; Liu, Y.-J.; Chen, W.; Zhao, G.; Chen, J. Recent advances in the role of toll-like receptors and tlr agonists in immunotherapy for human glioma. Protein Cell 2014, 5, 899–911.
- Jiang, Y.; Zhou, J.; Luo, P.; Gao, H.; Ma, Y.; Chen, Y.-S.; Li, L.; Zou, D.; Zhang, Y.; Jing, Z. Prosaposin promotes the proliferation and tumorigenesis of glioma through toll-like receptor 4 (Tlr4)-mediated nf-kappab signaling pathway. eBioMedicine 2018, 37, 78–90.
- Morad, G.; Helmink, B.A.; Sharma, P.; Wargo, J.A. Hallmarks of response, resistance, and toxicity to immune checkpoint blockade. Cell 2021, 184, 5309–5337.
- Hartley, G.P.; Chow, L.; Ammons, D.T.; Wheat, W.H.; Dow, S.W. Programmed cell death ligand 1 (Pd-L1) signaling regulates macrophage proliferation and activation. Cancer Immunol. Res. 2018, 6, 1260–1273.
- Xun, Y.; Yang, H.; Kaminska, B.; You, H. Toll-like receptors and toll-like receptor-targeted immunotherapy against glioma. J. Hematol. Oncol. 2021, 14, 176.
More
Information
Subjects:
Neurosciences
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
584
Revisions:
2 times
(View History)
Update Date:
01 Sep 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No