Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Maria Guadalupe Ortega Pierres | -- | 2733 | 2023-08-31 02:05:17 | | | |
| 2 | Camila Xu | Meta information modification | 2733 | 2023-08-31 04:25:41 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Argüello-García, R.; Carrero, J.C.; Ortega-Pierres, M.G. Cysteine Proteases Secreted by G. duodenalis. Encyclopedia. Available online: https://encyclopedia.pub/entry/48666 (accessed on 26 July 2026).
Argüello-García R, Carrero JC, Ortega-Pierres MG. Cysteine Proteases Secreted by G. duodenalis. Encyclopedia. Available at: https://encyclopedia.pub/entry/48666. Accessed July 26, 2026.
Argüello-García, Raúl, Julio César Carrero, M. Guadalupe Ortega-Pierres. "Cysteine Proteases Secreted by G. duodenalis" Encyclopedia, https://encyclopedia.pub/entry/48666 (accessed July 26, 2026).
Argüello-García, R., Carrero, J.C., & Ortega-Pierres, M.G. (2023, August 31). Cysteine Proteases Secreted by G. duodenalis. In Encyclopedia. https://encyclopedia.pub/entry/48666
Argüello-García, Raúl, et al. "Cysteine Proteases Secreted by G. duodenalis." Encyclopedia. Web. 31 August, 2023.
Copy Citation
Giardia duodenalis (syn. G. lamblia, G. intestinalis) is an intestinal protozoan parasite that causes giardiasis in humans and livestock, companion and wild animals. Giardiasis is a leading cause of waterborne diarrheal infections worldwide, with an estimated number of 280 million human cases per annum.
extracellular proteases
cysteine proteases
papain-like proteases
1. Introduction
Protistan parasites of the genera Giardia, Entamoeba, Cryptosporidium and Blastocystis cause intestinal infections and diseases (giardiasis, amoebiasis, cryptosporidiosis and blastocystosis) in humans and animals worldwide. Current evidence indicates that these pathogens collectively affect >1.5 billion people and cause ~300,000 deaths per annum [1][2][3]. These figures are likely to get worse due to climate change, extensive human travel and increased globalization.
All of these parasites colonize the intestinal tract and can cause acute or chronic diarrhea, malabsorption and/or bleeding, with severe sequelae, such as chronic bowel inflammation, reduced growth and development and in severe cases, death. However, infections can be self-limiting or asymptomatic, reflecting a degree of adaptation of these pathogens to their hosts and/or the effectiveness of host immune responses to suppress infections [4]. These pathogens have distinctive life cycles, which are linked to their particular tropism. For instance, Blastocystis and Giardia are extracellular parasites and Cryptosporidium is intracellular/extra cytoplasmic, whereas E. histolytica can invade tissues from the large intestine to spread (via the bloodstream) to the liver, spleen, kidney, brain and other organs, enabled by cellular structures for locomotion, attachment and penetration as well as by molecules acting on intra-, epi- or extracellular levels to promote parasite establishment and survival within the host.
Parasite structures and/or molecules that have an adverse effect on host intestinal homeostasis through mechanical, metabolic and functional damage can be virulence factors. These can include surface-anchored or secreted molecules such as toxins, lectins, cysteine-rich proteins, lytic enzymes (proteases, lipases and nucleases), metabolic enzymes and molecules with multiple functions, which are often called “moonlighting” proteins [5][6][7][8]. In the intestinal lumen and in intraepithelial compartments, intestinal protozoa and the molecular products that they release must overcome multiple barriers, such as mucus, epithelial cell surfaces (e.g., via microvilli, receptors and/or channels), intercellular junctions (tight and/or adherent junctions and desmosomes) and elements of the innate and adaptive immune systems (phagocytic/cytotoxic cells, IgA, antimicrobial peptides and cytokines). In this context, peptide hydrolases (proteases) play a key role. Seven broad groups of hydrolytic (i.e., aspartic, asparagine, cysteine, glutamic, metallo-, serine and threonine) proteases are classified in the current release (12.4) of the MEROPS database (https://www.ebi.ac.uk/merops/cgi-bin/family_index?type=P; accessed date: 2 August 2023) [9]. Cysteine proteases (CPs) are the most diverse type (families C1–C124), and tens to hundreds of CPs are encoded in the genomes of intestinal protists.
Several CPs are synthesized as inactive zymogens, which, upon proteolytic removal of an N-terminal prodomain, become catalytically competent. The catalytic mechanism of CPs involves the following steps: (a) deprotonation of a cysteine at the active site by an adjacent proton acceptor (usually histidine) that renders a thiolate-imidazolium charge, which is stabilized by a neighbor asparagine; (b) nucleophilic attack at the carbonyl carbon by the cysteine thiolate (proton acceptor), which promotes the release of the first half of the product and a thioester intermediate; (c) hydrolysis of the acyl-enzyme intermediate by activated water, which, in turn, releases the second half of the product with a terminal carboxylic acid, converting a CP into its initial state [10]. According to the MEROPS protease database, there are 14 CP groups called “clans” including nine C-type and five P-type groups. Evidence suggests that CPs of intestinal protozoa emerged from convergent evolution of the catalytic mechanism characteristic of clan CA (papain-like); in this clan, the most studied, abundant and relevant CPs are those in families C1 (cathepsin B/L-like) and C2 (calpain-like) [11][12].
2. Cysteine Proteases Secreted by G. duodenalis: An Arsenal of Cathepsin-like Enzymes with Multiple Targets
Giardia duodenalis (syn. G. lamblia, G. intestinalis) is an intestinal protozoan parasite that causes giardiasis in humans and livestock, companion and wild animals. Giardiasis is a leading cause of waterborne diarrheal infections worldwide, with an estimated number of 280 million human cases per annum. Giardiasis is included in the WHO’s Neglected Disease Initiative [13], and Giardia is considered a potential bioterrorism agent (category C) [14]. During its life cycle, Giardia alternates between the vegetative, pathogenic trophozoite and dormant, infective cyst forms. When a susceptible host ingests cysts contaminating water and food, excystation occurs during passage through the stomach into the small intestine. Emerging excyzoites rapidly divide by binary fission to give rise to trophozoites, which adhere to enterocytes via their adhesive disks and proliferate and progressively colonize the epithelial surfaces of the duodenum and midjejunum. Here, trophozoites are exposed to an alkaline pH and biliary secretions; cholesterol deprivation induces encystation, leading to the formation of cysts, which are shed in the feces.
Giardia infection can result in disease (giardiasis), which is symptomatic (acute to chronic) but can also be asymptomatic. The pathogenesis of giardiasis relates to virulence factors, which become active at the interface between trophozoites and small intestinal epithelium and are produced/released by trophozoites. These factors include cathepsin B proteases, tenascins, cystatin and a number of variant-specific surface proteins (VSPs) [15]. Of the secreted products, CP activity accounts for almost all of the parasite-derived proteolysis from supernatants recovered from trophozoite–epithelial cell interaction assays [16].
It has been reported that the genome of G. duodenalis (reference clone WBA6) encodes 25 CPs. Of these, 21 are cathepsin type: 8 cathepsin-L, 9 cathepsin-B with endopeptidase activity, 3 cathepsin-B with endopeptidase and dipeptidyl peptidase activity (associated with the presence of an ‘occluding loop’) and 1 cathepsin-C-like (the encystation-specific CP, ESCP) [17]. The repertoire of the cathepsin-like gene varies among strains and assemblages. Thus, in assemblage B, the GS strain has 17 homologs, while strain GS-B has 23; in assemblage E, the P15 strain contains 23 homologs. These proteases are activated following proteolytic removal of the N-terminal prodomain of their inactive precursors. During trophozoite growth and encystation, differential expression of cathepsin-B-like proteases occurs, including those formerly named CP2, CP4 and CP5, as well as the cathepsin-L-like protease CP17 [18], which are abundantly expressed [19]. This finding suggests that this family of proteases has diverse functional roles, depending on their location in the cellular compartments. These proteases, which are expressed and released by the parasite, have gelatinolytic and/or collagenolytic activities [16][20].
Interestingly, in G. duodenalis, some CPs are also encoded by other genes representing >230 variant surface proteins (VSPs). These VSPs are expressed as glycoproteins and/or palmitoyl-proteins, and their genes are transcriptionally regulated by an iRNA-silencing mechanism (named “all but one”) and are also involved in antigenic variation and the induction of protective immune responses. In this family, there is at least one clade of CPs with structural and/or functional similarities to cathepsins B. One of them is VSP9B10A (739 amino acids (aa) long; GL50803_101074), which can be detected in a secreted form (~75 kDa) during trophozoite–epithelial cell monolayer (IEC-6) interactions and displays proteolytic activity [21]. VSP9B10A contains a central region (aa 324 to 684) with a catalytic substrate binding pocket and calcium recognition sites of a typical cathepsin B. This atypical cathepsin is not always expressed on the trophozoite surface but has been shown to be toxic to epithelial monolayers [21], leading to a loss of cell-cell and cell-substrate contacts (Figure 1). Experimental evidence has demonstrated that VSP9B10A is a conditional virulence factor [21].
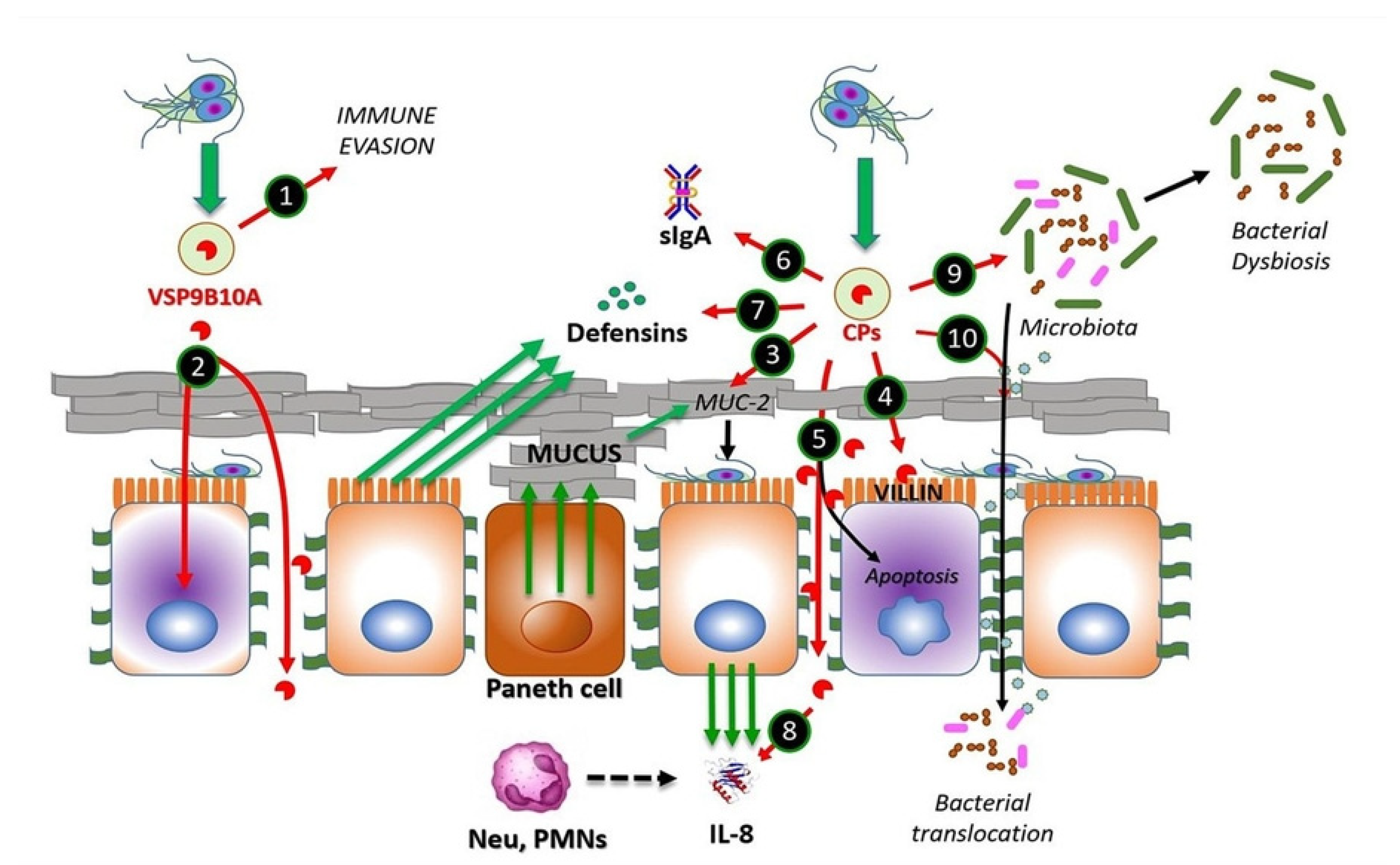
Figure 1. CPs from Giardia duodenalis trophozoites interact with multiple targets in the small intestine. Experimental models of trophozoites–epithelial cell interactions indicate that the parasite releases CPs, mainly cathepsin B-type, including Giardipain-1 (GL50803_14019), GL16160 and GL16779 and non-canonical CPs (e.g., VSP9B10A). This latter CP may serve, when expressed, to divert the immune system (1) or may cause damage to the epithelium due to a loss of cell–cell junction and cytotoxicity in the cell (2). Secreted CPs have been shown to degrade substrates, such as mucin 2, an important component of intestinal mucus (3), enabling trophozoite adhesion to microvilli. Direct damage to epithelial cell integrity by CPs may include disruption of the cytoskeletal microvillus-resident protein villin (4) and the disruption of tight junction proteins, such as ZO-1 and claudins, also involving adherens junction proteins including β-catenin or E-cadherin (5). Soluble elements of the innate immune response, including secretory IgA (6), produced by plasmatic B-cells, defensins (7) and IL-8 (8), both produced by epithelial cells, of which the latter works as neutrophil attractant, might be degraded by giardial CPs. These enzymes also provoke alterations in the microbiome of the small intestine, leading to dysbiosis (9), while bacterial translocation from the luminal to the intraepithelial compartment (10) may be promoted via the degradation of intercellular junctions by CPs as mentioned. Recent studies suggest that giardial CPs might be secreted after removal of N-terminal prodomain and inclusion into membrane-bound extracellular vesicles (EVs), mainly exosomes (green-filled circles).
Giardia CPs might be regulated at the intracellular level by cystatins, as is the case for Cryptosporidium (cryptostatin) [22]. Cystatins comprise a superfamily of proteins with a homologous sequence and structure, consisting of an antiparallel β-sheet topped with an α-helix. These function as endogenous cysteine protease inhibitors of target proteases, such as cathepsins B, C, L and H, and are currently classified into type 1 (stefins), type 2 (cystatins), type 3 (kininogens), latexins and fetuins [23]. In G. duodenalis, a stefin (GL50803_27918) was identified as a cytoplasmic protein that potently inhibits three major secreted CPs of the clan CA papain-type, namely GL50803_14019 (Giardipain-1), _16160 and _16779. Interestingly, stefin is a weak inhibitor of human cathepsin B [24] and might play a role in Giardia–epithelial cell interaction, as previous evidence revealed an upregulation of cathepsin-L-like proteins GL50803_137680 or GL_50803_3099 [25] upon the interaction of distinct strains (WB, P-1, GS/M or NF) of Giardia with ICE-6 monolayers [26]. Another interaction study [27] revealed conspicuous expression of cathepsin B-like proteases in the WB strain of Giardia (i.e., GL50803_16160, _16468 and _16779) and their homologs in the GS strain (i.e., GL50581_2946, _438 and _78), in addition to other cathepsins B (including GL50803_14019 and GL50581_2036). A proteomic study of the same strains (using the same intestinal cell line) reported a distinct repertoire of three cathepsin B-like proteases in the WB strain of Giardia (i.e., GL50803_15564, _16468 and _17516) and their closest homolog in the GS strain (GL50581_2036, _438 and _2318) [28]. This information suggests that trophozoites of the zoonotic assemblages A and B (represented by the strains WB and GS, respectively) exhibit similar CP secretion profiles upon interaction with epithelial cells.
Following secretion, these proteases become catalytically competent due to the absence of an occluding loop and the intracellular excision of prodomains. In the case of Giardipain-1 (GL50803_ 14019, formerly named CP2), its precursor (33 kDa) is 8 kDa larger, i.e., 76 amino acids (aa) longer than the mature protein (25 kDa), which possesses gelatinolytic activity and exhibits pro-apoptotic effects on IEC-6 and epithelial cell (MDCK) monolayers [29]. The CPs secreted by Giardia have different protein targets on host epithelial cells and eventually disrupt intestinal homeostasis, in turn causing small intestinal damage [30] (cf. Figure 1).
The mechanism underlying the secretion of Giardia CPs is not yet fully understood. However, there is evidence for the processing of the N-terminal prodomain of clan CA proteases, such as Giardipain-1 [18][29]. In a recent proteomic study of extracellular vesicles (EVs; <100 nm in size) released by Giardia trophozoites, cathepsin B-like proteases have been identified as cargo within exosomes [31] (Figure 1). Considering the role of EVs in the intercellular crosstalk between parasites and intestinal epithelial cells, it is likely that these vesicles may mediate the release of cargo proteins around or within epithelial cells to promote cellular and tissue damage.
One of the first barriers that trophozoites encounter is the mucus layer on the brush border of the intestinal epithelium. This layer is mainly composed of membrane-bound mucins, such as MUC4 and MUC16, and gel-forming mucins, such as MUC2, the latter of which predominates in the gastrointestinal tract [32]. Interestingly, the use of a human mucus-producing colonic cell line (LS174T) in interaction assays using excretory/secretory proteins (ESPs) from strains NF (assemblage A) and GS/M (assemblage B) revealed a concentration-dependent degradation of MUC2, which was abrogated by pre-treatment with a CP inhibitor, called E-64 [33] (Figure 1). In addition, a recent investigation [34] reported a novel mechanism by which Giardia CPs modulate goblet cell activity via the cleavage and activation of protease-activated receptor 2 (PAR-2), which can regulate mucin gene expression in intestinal goblet cells.
Another barrier against Giardia is the brush border at the apical surface of enterocytes. However, it has been shown that trophozoites adhering to the microvillus lining of the enterocytes cause damage to the intestinal epithelium, leading to structural alterations and decreased absorptive surface area [35]. In this context, villin, an actin-binding protein expressed in the microvillus layer of enterocytes, can be degraded and redistributed by the action of surface-exposed Giardia CPs (Figure 1), supported by evidence showing the intact villin structure of Caco-2 cells exposed to trophozoites pre-incubated with inhibitors, such as the membrane-permeant E-64 or ML-9 (inhibitor of myosin light chain phosphorylation, MLCK) [36].
Further damage by proteases secreted by G. duodenalis trophozoites can also occur between cells in the intestinal epithelium. This has been shown via the disruption of transmembrane, plaque and filamentous proteins intervening apical junctional complexes (AJCs), known as tight junctions (claudins, occludins and/or zonula occludens—ZO-proteins), and adherent junctions (E-cadherin and β-catenin) [29][37]. There is clear evidence that Giardia ESPs, when added to Caco-2 cells, can alter the distribution of AJC components, such as claudin-1, occludin, ZO-1 and intracellular F-actin [37]. Furthermore, when added to IEC-6 monolayers, purified secreted protease Giardipain-1 has been shown to colocalize with claudin-1 and occludin at cell junctions, affecting distribution, decreasing transepithelial electrical resistance and inducing apoptosis, leading to pathological changes in epithelial cells [29]. Other experiments have shown that recombinant forms of each of the three secreted CPs, designated GL50803_14019 (Giardipain-1), GL50803_16160 and GL50803_16779 (formerly CP3) [27], affect the distribution of AJC proteins [37] and also degrade E-cadherin and occludin, which could be reduced by inhibitor E-64d [37]. Further, the three proteases degraded claudin-1 and -4 and β-catenin, respectively, suggesting that the limited number of CPs released by trophozoites have a redundant activity on cell junction proteins (Figure 1). The proteolytic activity of secreted proteases on specific substrates was shown using recombinant proteins in a two-thioredoxin system. Activity was associated with Giardipain-1, GL50803_16160 and GL50803_16779. Similar substrate selectivities were observed for Giardipain-1 and GL50803_16160; however, these differed from GL50803_16779 [24][38].
Bioinformatics analyses performed to screen the human small intestinal proteome to determine the cleavage consensus sequence of GL50803_16779 predicted immunoglobulins (Ig), defensins (Df) and chemokines (Ck) as targets, suggesting that parasite proteases interfere with adaptive host immune responses (Figure 1) [24][37][38][39][40]. Moreover, the different proteases degraded recombinant human defensins αDf1, βDf5 and αDf6, whereas the activity of Giardipain-1 was higher than those recorded for other proteases. Giardipain-1 may participate in neutralizing the innate response induced by Paneth cell defensins [24][38]. Chemokines/cytokines produced by epithelial cells, including CXCL1–3, are upregulated upon exposure to trophozoites [39]. In this context, it has been shown that interleukin-8 (CXCL8) is degraded by Giardia CPs, thereby reducing neutrophil recruitment and inflammatory responses [40], while recombinant (and upregulated) chemokines/cytokines CXCL1–3, CXCL8, CCL2 and CCL20 were cleaved by either ESPs or recombinant GL-50803_16160 [37]. These studies suggest that CPs participate in the onset and establishment of Giardia infection and also modulate host immune responses.
In experiments in which device-isolated biofilms and microbiota-free mice were used, it was observed that secreted CPs play a role in the interplay of Giardia with the commensal microbiota communities located in the gut as biofilm-forming, planktonic bacteria. In these studies, a proinflammatory response was observed, even in the absence of Giardia, suggesting that CPs from Giardia play a role in intestinal disease, including irritable bowel ensuing infection [41][42]. Interestingly, recent experiments from researchers' group [30] have shown that, in an experimental infection system, purified Giardipain-1 induces apoptosis and the extrusion of epithelial cells at the tips of the villi in ligated intestinal loops in jirds (Meriones unguiculatus). Moreover, the infection of jirds with trophozoites expressing Giardipain-1 resulted in intestinal epithelial damage, cellular infiltration, crypt hyperplasia, goblet cell hypertrophy and edema. Pathological alterations were pronounced when jirds were infected (via gavage) with Giardia trophozoites that stably overexpressed Giardipain-1. Furthermore, Giardia colonization in jirds resulted in chronic inflammation, which might relate to dysbiosis triggered by the parasite (Figure 1). These results demonstrated that Giardipain-1-induced alterations seen in the intestinal epithelial cells and in the microbiome contribute to the pathogenesis of giardiasis and its associated complications [30]. However, experimental evidence indicates that it is possible that other major secreted proteases such as GL50803_16160 and GL50803_16779, which are structurally similar to Giardipain-1, may have similar effects during G. duodenalis infection.
References
- Ximénez, C.; Cerritos, R.; Rojas, L.; Dolabella, S.; Morán, P.; Shibayama, M.; González, E.; Valadez, A.; Hernández, E.; Valenzuela, O.; et al. Human Amebiasis: Breaking the Paradigm? Int. J. Environ. Res. Public Health 2010, 7, 1105–1120.
- Chalmers, R.M. Chapter Sixteen—Cryptosporidium. In Microbiology of Waterborne Diseases, 2nd ed.; Percival, S.L., Yates, M.V., Williams, D.W., Chalmers, R.M., Gray, N.F., Eds.; Academic Press: London, UK, 2014; pp. 287–326. ISBN 978-0-12-415846-7.
- Dogruman-Al, F.; Simsek, Z.; Boorom, K.; Ekici, E.; Sahin, M.; Tuncer, C.; Kustimur, S.; Altinbas, A. Comparison of Methods for Detection of Blastocystis Infection in Routinely Submitted Stool Samples, and Also in IBS/IBD Patients in Ankara, Turkey. PLoS ONE 2010, 5, e15484.
- Argüello-García, R.; Carrero, J.C.; Ortega-Pierres, G. Host Immune Responses Against Intestinal Unicellular Parasites and Their Role in Pathogenesis and Protection. In Reference Module in Biomedical Sciences; Elsevier: Amsterdam, The Netherlands, 2020.
- Skotarczak, B. Genetic Diversity and Pathogenicity of Blastocystis. Ann. Agric. Environ. Med. 2018, 25, 411–416.
- Arguello-Garcia, R.; Ortega-Pierres, M.G. Giardia duodenalis Virulence—“To Be, or Not To Be”. Curr. Trop. Med. Rep. 2021, 8, 246–256.
- Guérin, A.; Striepen, B. The Biology of the Intestinal Intracellular Parasite Cryptosporidium. Cell Host Microbe 2020, 28, 509–515.
- Guillén, N. Pathogenicity and Virulence of Entamoeba histolytica, the Agent of Amoebiasis. Virulence 2023, 14, 2158656.
- Rawlings, N.D.; Barrett, A.J.; Thomas, P.D.; Huang, X.; Bateman, A.; Finn, R.D. The MEROPS Database of Proteolytic Enzymes, Their Substrates and Inhibitors in 2017 and a Comparison with Peptidases in the PANTHER Database. Nucleic Acids Res. 2018, 46, D624–D632.
- Roy, M.; Rawat, A.; Kaushik, S.; Jyoti, A.; Srivastava, V.K. Endogenous Cysteine Protease Inhibitors in Upmost Pathogenic Parasitic Protozoa. Microbiol. Res. 2022, 261, 127061.
- Sajid, M.; McKerrow, J.H. Cysteine Proteases of Parasitic Organisms. Mol. Biochem. Parasitol. 2002, 120, 1–21.
- Siqueira-Neto, J.L.; Debnath, A.; McCall, L.-I.; Bernatchez, J.A.; Ndao, M.; Reed, S.L.; Rosenthal, P.J. Cysteine Proteases in Protozoan Parasites. PLoS Negl. Trop. Dis. 2018, 12, e0006512.
- Savioli, L.; Smith, H.; Thompson, A. Giardia and Cryptosporidium Join the “Neglected Diseases Initiative”. Trends Parasitol. 2006, 22, 203–208.
- CDC. Centers for Disease Control and Prevention. Bioterrorism Agents/Diseases. Available online: https://emergency.cdc.gov/agent/agentlist-category.asp (accessed on 9 August 2023).
- Emery, S.J.; Mirzaei, M.; Vuong, D.; Pascovici, D.; Chick, J.M.; Lacey, E.; Haynes, P.A. Induction of Virulence Factors in Giardia duodenalis Independent of Host Attachment. Sci. Rep. 2016, 6, 20765.
- Rodríguez-Fuentes, G.B.; Cedillo-Rivera, R.; Fonseca-Liñán, R.; Argüello-García, R.; Muñoz, O.; Ortega-Pierres, G.; Yépez-Mulia, L. Giardia duodenalis: Analysis of Secreted Proteases upon Trophozoite-Epithelial Cell Interaction in vitro. Mem. Inst. Oswaldo Cruz. 2006, 101, 693–696.
- Touz, M.C.; Nores, M.J.; Slavin, I.; Carmona, C.; Conrad, J.T.; Mowatt, M.R.; Nash, T.E.; Coronel, C.E.; Luján, H.D. The Activity of a Developmentally Regulated Cysteine Proteinase Is Required for Cyst Wall Formation in the Primitive Eukaryote Giardia lamblia. J. Biol. Chem. 2002, 277, 8474–8481.
- Ward, W.; Alvarado, L.; Rawlings, N.D.; Engel, J.C.; Franklin, C.; McKerrow, J.H. A Primitive Enzyme for a Primitive Cell: The Protease Required for Excystation of Giardia. Cell 1997, 89, 437–444.
- DuBois, K.N.; Abodeely, M.; Sajid, M.; Engel, J.C.; McKerrow, J.H. Giardia lamblia Cysteine Proteases. Parasitol. Res. 2006, 99, 313–316.
- Jimenez, J.C.; Uzcanga, G.; Zambrano, A.; Di Prisco, M.C.; Lynch, N.R. Identification and Partial Characterization of Excretory/Secretory Products with Proteolytic Activity in Giardia intestinalis. J. Parasitol. 2000, 86, 859–862.
- Cabrera-Licona, A.; Solano-González, E.; Fonseca-Liñán, R.; Bazán-Tejeda, M.L.; Raúl Argüello-García, M.L.; Bermúdez-Cruz, R.M.; Ortega-Pierres, G. Expression and Secretion of the Giardia duodenalis Variant Surface Protein 9B10A by Transfected Trophozoites Causes Damage to Epithelial Cell Monolayers Mediated by Protease Activity. Exp. Parasitol. 2017, 179, 49–64.
- Kang, J.-M.; Ju, H.-L.; Yu, J.-R.; Sohn, W.-M.; Na, B.-K. Cryptostatin, a Chagasin-Family Cysteine Protease Inhibitor of Cryptosporidium parvum. Parasitology 2012, 139, 1029–1037.
- Yan, B.; Ren, Y.; Liu, C.; Shu, L.; Wang, C.; Zhang, L. Cystatin SN in Type 2 Inflammatory Airway Diseases. J. Allergy Clin. Immunol. 2023, 151, 1191–1203.e3.
- Liu, J.; Svärd, S.G.; Klotz, C. Giardi iIntestinalis Cystatin Is a Potent Inhibitor of Papain, Parasite Cysteine Proteases and, to a Lesser Extent, Human Cathepsin B. FEBS Lett. 2019, 593, 1313–1325.
- Ringqvist, E.; Avesson, L.; Söderbom, F.; Svärd, S.G. Transcriptional Changes in Giardia during Host-Parasite Interactions. Int. J. Parasitol. 2011, 41, 277–285.
- Ma’ayeh, S.Y.; Brook-Carter, P.T. Representational Difference Analysis Identifies Specific Genes in the Interaction of Giardia duodenalis with the Murine Intestinal Epithelial Cell Line, IEC-6. Int. J. Parasitol. 2012, 42, 501–509.
- Ma’ayeh, S.Y.; Liu, J.; Peirasmaki, D.; Hörnaeus, K.; Bergström Lind, S.; Grabherr, M.; Bergquist, J.; Svärd, S.G. Characterization of the Giardia intestinalis Secretome during Interaction with Human Intestinal Epithelial Cells: The Impact on Host Cells. PLoS Negl. Trop. Dis. 2017, 11, e0006120.
- Dubourg, A.; Xia, D.; Winpenny, J.P.; Al Naimi, S.; Bouzid, M.; Sexton, D.W.; Wastling, J.M.; Hunter, P.R.; Tyler, K.M. Giardia Secretome Highlights Secreted Tenascins as a Key Component of Pathogenesis. Gigascience 2018, 7, giy003.
- Ortega-Pierres, G.; Argüello-García, R.; Laredo-Cisneros, M.S.; Fonseca-Linán, R.; Gómez-Mondragón, M.; Inzunza-Arroyo, R.; Flores-Benítez, D.; Raya-Sandino, A.; Chavez-Munguía, B.; Ventura-Gallegos, J.L.; et al. Giardipain-1, a Protease Secreted by Giardia duodenalis Trophozoites, Causes Junctional, Barrier and Apoptotic Damage in Epithelial Cell Monolayers. Int. J. Parasitol. 2018, 48, 621–639.
- Quezada-Lázaro, R.; Vázquez-Cobix, Y.; Fonseca-Liñán, R.; Nava, P.; Hernández-Cueto, D.D.; Cedillo-Peláez, C.; López-Vidal, Y.; Huerta-Yepez, S.; Ortega-Pierres, M.G. The Cysteine Protease Giardipain-1 from Giardia duodenalis Contributes to a Disruption of Intestinal Homeostasis. Int. J. Mol. Sci. 2022, 23, 13649.
- Gavinho, B.; Sabatke, B.; Feijoli, V.; Rossi, I.V.; da Silva, J.M.; Evans-Osses, I.; Palmisano, G.; Lange, S.; Ramirez, M.I. Peptidylarginine Deiminase Inhibition Abolishes the Production of Large Extracellular Vesicles from Giardia intestinalis, Affecting Host-Pathogen Interactions by Hindering Adhesion to Host Cells. Front. Cell. Infect. Microbiol. 2020, 10, 417.
- Johansson, M.E.V.; Hansson, G.C. Immunological Aspects of Intestinal Mucus and Mucins. Nat. Rev. Immunol. 2016, 16, 639–649.
- Allain, T.; Amat, C.B.; Motta, J.-P.; Manko, A.; Buret, A.G. Interactions of Giardia sp. with the Intestinal Barrier: Epithelium, Mucus, and Microbiota. Tissue Barriers 2017, 5, e1274354.
- Fekete, E.; Allain, T.; Amat, C.B.; Mihara, K.; Saifeddine, M.; Hollenberg, M.D.; Chadee, K.; Buret, A.G. Giardia duodenalis Cysteine Proteases Cleave Proteinase-Activated Receptor-2 to Regulate Intestinal Goblet Cell Mucin Gene Expression. Int. J. Parasitol. 2022, 52, 285–292.
- Troeger, H.; Epple, H.-J.; Schneider, T.; Wahnschaffe, U.; Ullrich, R.; Burchard, G.-D.; Jelinek, T.; Zeitz, M.; Fromm, M.; Schulzke, J.-D. Effect of Chronic Giardia lamblia Infection on Epithelial Transport and Barrier Function in Human Duodenum. Gut 2007, 56, 328–335.
- Bhargava, A.; Cotton, J.A.; Dixon, B.R.; Gedamu, L.; Yates, R.M.; Buret, A.G. Giardia duodenalis Surface Cysteine Proteases Induce Cleavage of the Intestinal Epithelial Cytoskeletal Protein Villin via Myosin Light Chain Kinase. PLoS ONE 2015, 10, e0136102.
- Liu, J.; Ma’ayeh, S.; Peirasmaki, D.; Lundström-Stadelmann, B.; Hellman, L.; Svärd, S.G. Secreted Giardia intestinalis Cysteine Proteases Disrupt Intestinal Epithelial Cell Junctional Complexes and Degrade Chemokines. Virulence 2018, 9, 879–894.
- Liu, J.; Fu, Z.; Hellman, L.; Svärd, S.G. Cleavage Specificity of Recombinant Giardia intestinalis Cysteine Proteases: Degradation of Immunoglobulins and Defensins. Mol. Biochem. Parasitol. 2019, 227, 29–38.
- Roxström-Lindquist, K.; Ringqvist, E.; Palm, D.; Svärd, S. Giardia lamblia-Induced Changes in Gene Expression in Differentiated Caco-2 Human Intestinal Epithelial Cells. Infect. Immun. 2005, 73, 8204–8208.
- Cotton, J.A.; Bhargava, A.; Ferraz, J.G.; Yates, R.M.; Beck, P.L.; Buret, A.G. Giardia duodenalis Cathepsin B Proteases Degrade Intestinal Epithelial Interleukin-8 and Attenuate Interleukin-8-Induced Neutrophil Chemotaxis. Infect. Immun. 2014, 82, 2772–2787.
- Beatty, J.K.; Akierman, S.V.; Motta, J.-P.; Muise, S.; Workentine, M.L.; Harrison, J.J.; Bhargava, A.; Beck, P.L.; Rioux, K.P.; McKnight, G.W.; et al. Giardia duodenalis Induces Pathogenic Dysbiosis of Human Intestinal Microbiota Biofilms. Int. J. Parasitol. 2017, 47, 311–326.
- Keselman, A.; Li, E.; Maloney, J.; Singer, S.M. The Microbiota Contributes to CD8+ T Cell Activation and Nutrient Malabsorption Following Intestinal Infection with Giardia Duodenali. Infect. Immun. 2016, 84, 2853–2860.
More
Information
Subjects:
Parasitology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
655
Revisions:
2 times
(View History)
Update Date:
31 Aug 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No