 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Gera Neufeld | -- | 2612 | 2023-08-23 23:01:11 | | | |
| 2 | Peter Tang | Meta information modification | 2612 | 2023-08-24 04:41:18 | | |
Video Upload Options
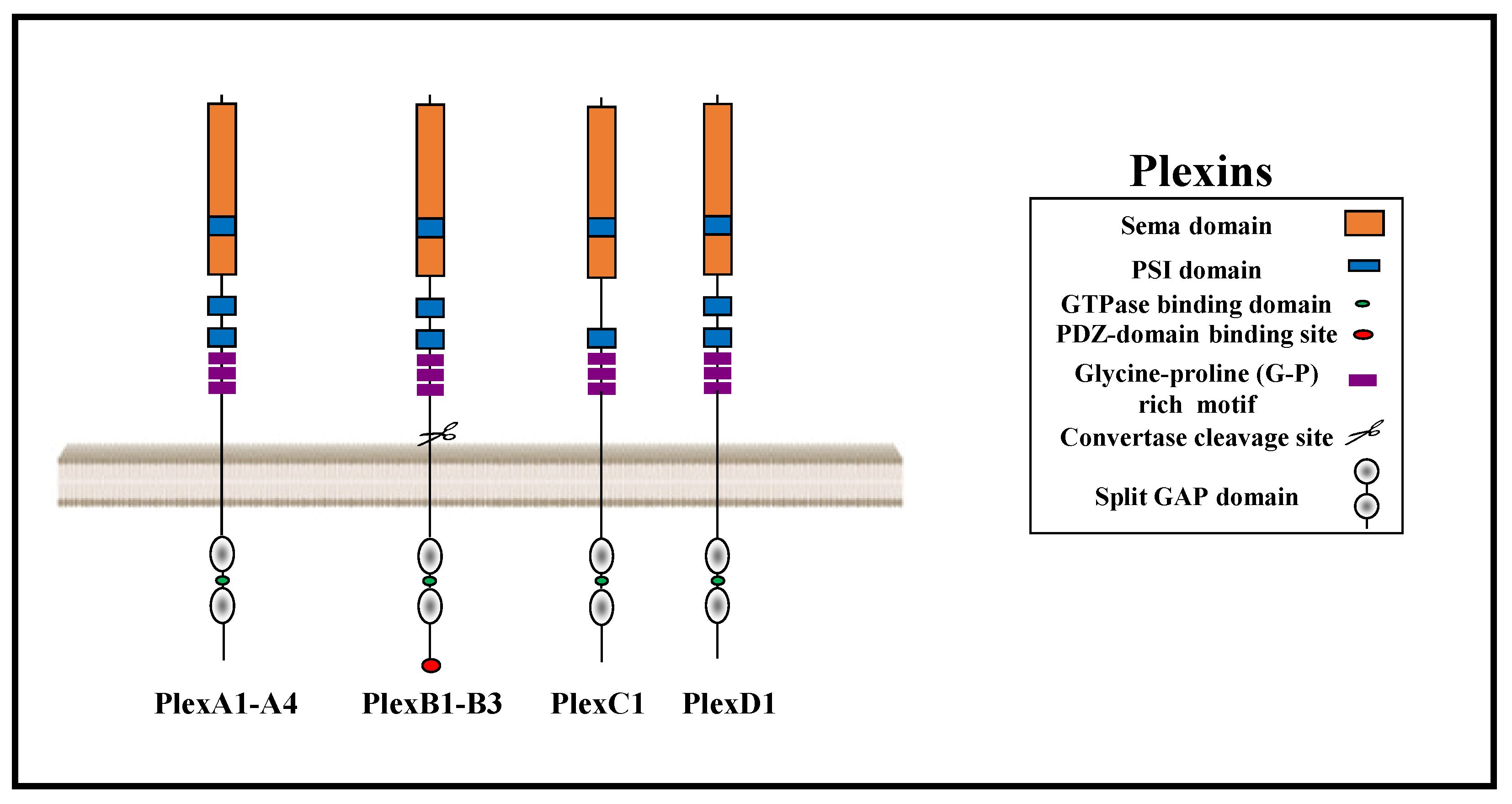
Plexins are a family of nine single-pass transmembrane receptors with a conserved GTPase activating protein (GAP) domain. The plexin family is divided into four subfamilies: Type-A, type-B, type-C, and type-D plexins. Plexins function as receptors for axon guidance factors of the semaphorin family. The semaphorin gene family contains 22 genes that are divided into eight subclasses of which subclasses three to seven represent vertebrate semaphorins. The plexins and their semaphorin ligands have important roles as regulators of angiogenesis, cancer proliferation, and metastasis. Class 3 semaphorins, with the exception of sema3E, are the only semaphorins that do not bind directly to plexins. In order to transduce their signals, they bind instead to complexes consisting of receptors of the neuropilin family and various plexins. Some plexins also form complexes with tyrosine-kinase receptors such as the epidermal growth factor receptor ErbB2, the mesenchymal epithelial transition factor receptor (MET), and the Vascular endothelial growth factor receptor 2 (VEGFR2) and, as a result, can modulate cell proliferation and tumor progression.
1. The Plexin Receptor Family
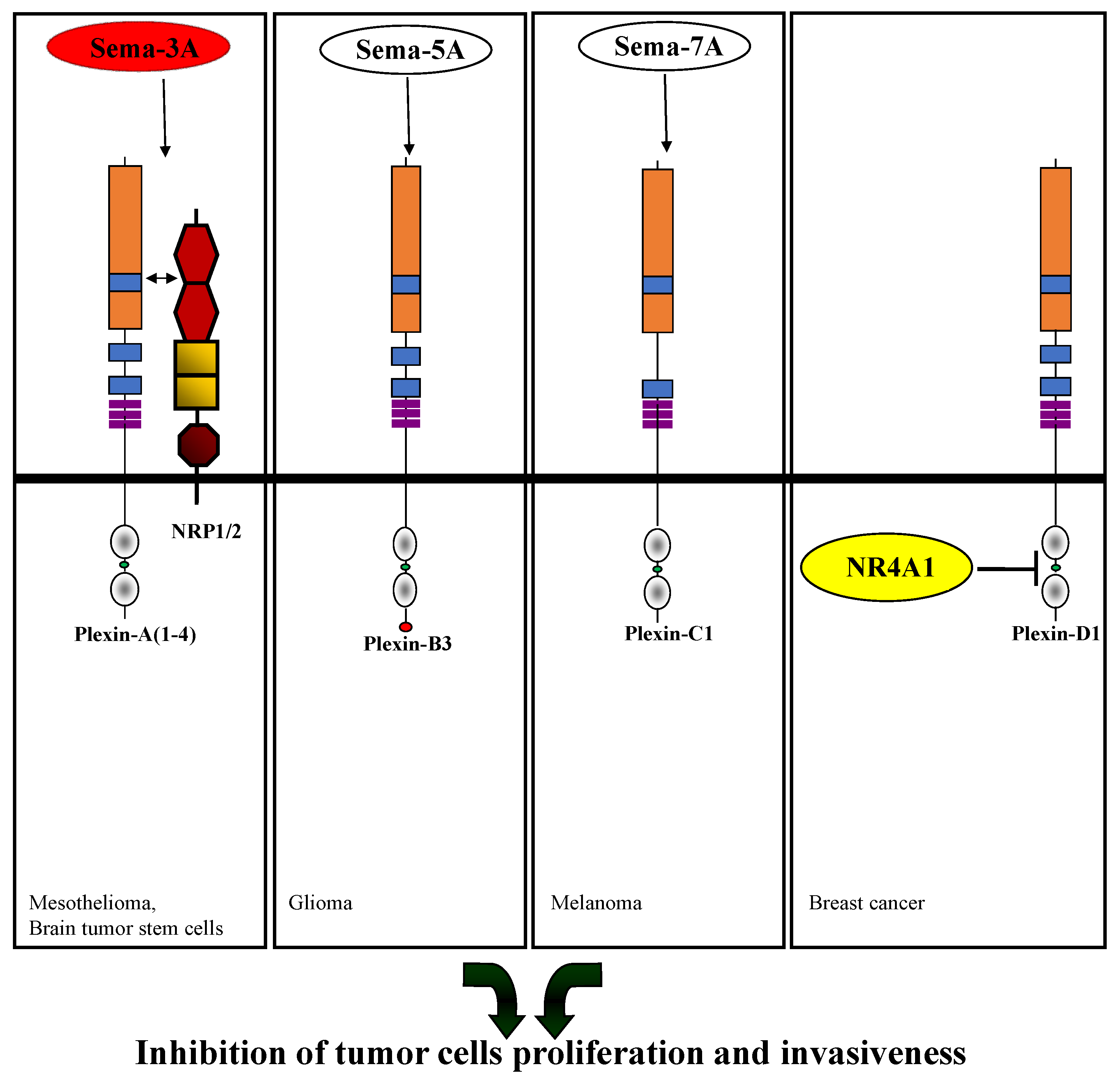
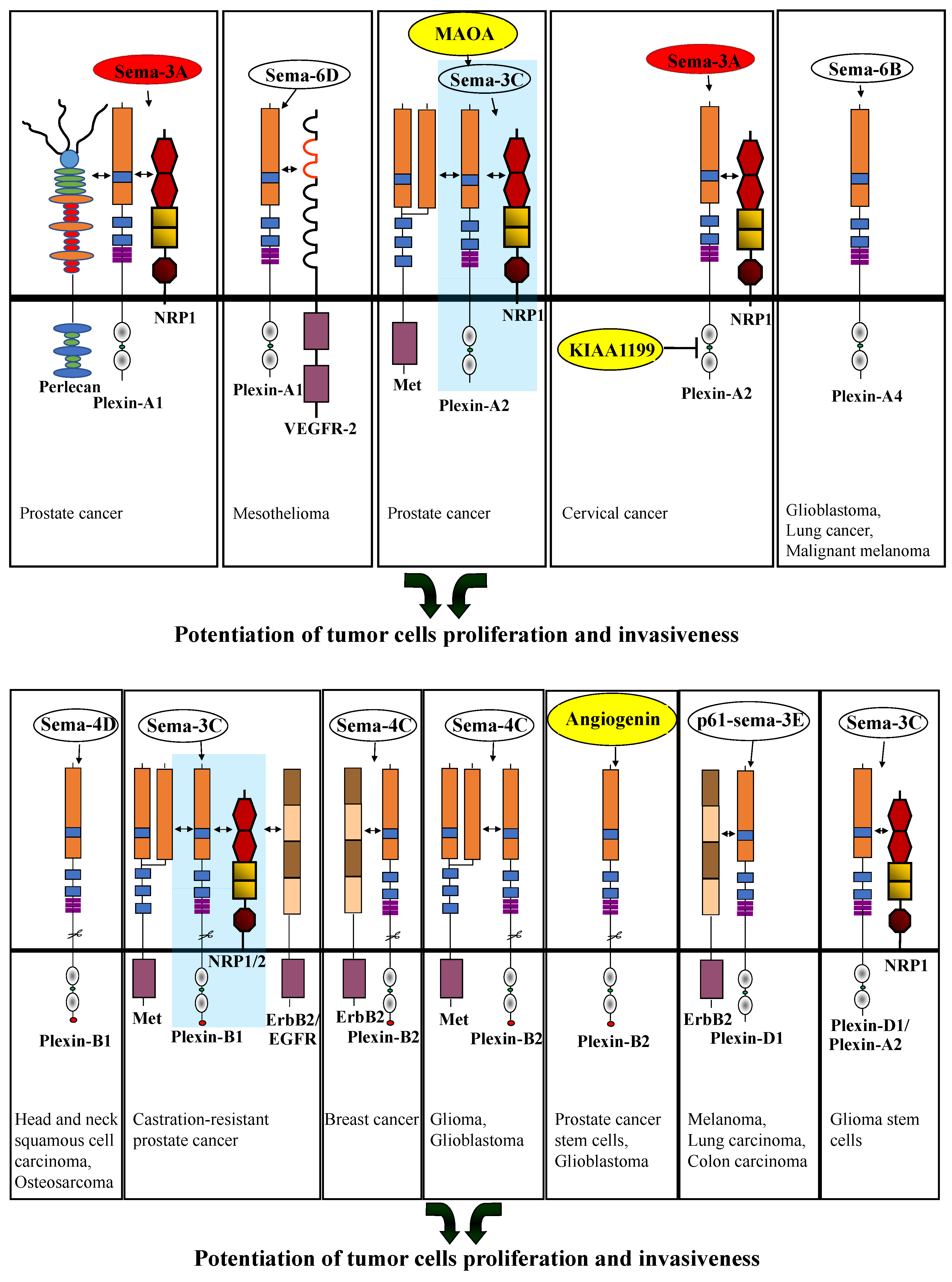
2. The Role of the Different Plexins in Tumor Progression
2.1. Type-A Plexins
|
Plexin |
Modulators of Plexin-Mediated Signal Transduction |
Role |
Cancer Type |
Refs |
|---|---|---|---|---|
|
Plexin-A1 |
Sema3A |
Inhibition of proliferation |
Malignant mesothelial cells |
[28] |
|
Sema3A, NRP-1, Perlecan |
Promotion of metastatic dissemination |
Prostate cancer cells |
[29] |
|
|
Sema6D, VEGFR-2 |
Promotion of survival and tumor growth |
Malignant mesothelioma cells |
[30] |
|
|
Sema3A |
Promotion of proliferation and glycolytic activity |
Lung cancer cells |
[31] |
|
|
Sema3A, NRP-1 |
Inhibition of proliferation |
Brain tumor stem cells |
[32] |
|
|
Plexin-A2 |
Enhancement of migration and invasion |
Prostate cancer cells |
[33] |
|
|
Sema3C, NRP-1, MAOA, MET |
Promotion of perineural invasion |
Prostate cancer cells |
[34] |
|
|
Sema3A, KIAA1199 |
Inhibition of apoptosis |
Cervical cancer cells |
[35] |
|
|
Enabling cell proliferation and the development of tumors |
Glioblastoma derived cells |
[36] |
||
|
Plexin-A3 |
Inhibition of cell invasion |
Epithelial ovarian cancer cells |
[37] |
|
|
Plexin-A4 |
Sema6B |
Promotion of pro- proliferative signals |
Glioblastoma-derived cells, lung-cancer-derived cells, malignant-melanoma-derived cells |
[26] |
|
miR-564 |
Promotion of cell proliferation and migration |
Non-small cell lung carcinoma cells |
[38] |
|
|
Plexin-B1 |
Sema4D |
Promotion of EMT and tumor cell metastasis |
Head and neck squamous cell carcinoma |
[39] |
|
Sema4D |
Promotion of cell invasion, proliferation, and migration |
Osteosarcoma cells |
[40] |
|
|
Sema3C, NRP-1/2, EGFR, ErbB2, MET |
Promotion of cancer growth |
Castration-resistant prostate cancer cells |
[41] |
|
|
TMPRSS2-ERG |
Promotion of cell migration and invasion |
Prostate cancer cells |
[42] |
|
|
Promotion of cell migration and invasion |
Ovarian cancer derived cells |
[43] |
||
|
Inhibition of breast cancer cell motility |
Breast cancer cells |
[44] |
||
|
Suppression of tumorigenesis |
Primary melanoma cells |
[45] |
||
|
Inhibition of cell proliferation |
Basal cell carcinoma cells |
[46] |
||
|
Plexin-B2 |
Inhibition of cell proliferation |
Basal cell carcinoma cells |
[46] |
|
|
Sema4C, ErbB2 |
Promotion of proliferation and development of tumor metastasis |
Breast cancer derived cells |
[47] |
|
|
Sema4C, MET |
Promotion of glioma and glioblastoma cell invasion |
Glioma and Glioblastoma cells |
[48] |
|
|
Angiogenin |
Inhibition of tumor cell proliferation and inhibition of tumor development |
Glioblastoma cells, breast cancer cells, and myelogenous leukemia cells |
[49] |
|
|
Angiogenin |
Enhancement of CSC stemness and resistance to chemotherapy |
Prostate cancer stem cells |
[50] |
|
|
Angiogenin |
Promotion of proliferation, invasion, and tumor growth |
Glioblastoma cells |
[51] |
|
|
Circular RNA, Circ_0013958 |
Promotion of proliferation, migration, invasion, and tumor growth |
Ovarian cancer cells |
[52] |
|
|
EGFR |
Promotion of proliferation, invasiveness and tumor-forming ability by constitutively active plexin-B2 |
Stem cells from cancers from unknown primary tumors |
[53] |
|
|
Plexin-B3 |
Sema5A |
Inhibition of migration and invasion |
Glioma cells |
[54] |
|
Inhibition of migration, invasion, and tumor metastasis |
Pancreatic cancer cells |
[55] |
||
|
Promotion of cancer cell growth, cell migration, cell invasion, and tumor progression |
Triple-negative breast cancer cells |
[56] |
||
|
Plexin-C1 |
Sema7A |
Inhibition of tumor progression |
Melanoma cells |
[57] |
|
Inhibition of tumor progression |
Glioma cells |
[58] |
||
|
Sema7A, β1 integrin receptors |
Promotion of metastasis |
Melanoma and breast cancer cells |
[59] |
|
|
Promotion of migration and proliferation |
Gastric cancer cells |
[60] |
||
|
Plexin-D1 |
P61-sema3E, ErbB2 |
Promotion of tumor cell invasiveness and tumor cell metastasis |
Melanoma cells, lung carcinoma cells, colon carcinoma cells |
|
|
NR4A1 |
Promotion of apoptosis |
Breast cancer cells |
[63] |
|
|
Sema3C, plexin-A2, NRP-1 |
Promotion of cell survival |
Glioma stem cells |
[64] |
2.2. Type-B Plexins
2.3. Plexin-C1
2.4. Plexin-D1
3. Conclusions
Plexins are a family of single-pass transmembrane receptors which have the ability to inhibit or potentiate tumor cell proliferation and invasiveness. Plexins function primarily as semaphorin receptors. Various plexins are expressed in tumor cells as well as in a variety of other cell types, which are recruited to the tumor microenvironment, such as endothelial cells. The final effects on tumor progression are, of course, an integration of all of these effects. Here only the direct effects of plexin-mediated signal transduction on the behavior of tumor cells were covered. Interestingly, the effects that given semaphorins have on the behavior of tumor cells are strongly modulated by the association of their plexin and neuropilin receptors with additional cell surface molecules. For example, sema3A,which normally inhibits tumor cell invasiveness when signaling through its plexin-A1 receptor, induces tumor cell invasiveness following the association of its plexin-A1 receptor with cell surface-associated perlecan. It is likely that in addition to the known interactions that were covered here, there are additional interactions that can also modulate plexin mediated responses of tumor cells to semaphorins. The identification of such modulating interactions is likely to become the focus of future studies.
References
- Hota, P.K.; Buck, M. Plexin structures are coming: Opportunities for multilevel investigations of semaphorin guidance receptors, their cell signaling mechanisms, and functions. Cell Mol. Life Sci. 2012, 69, 3765–3805.
- Luo, Y.; Raible, D.; Raper, J.A. Collapsin: A protein in brain that induces the collapse and paralysis of neuronal growth cones. Cell 1993, 75, 217–227.
- Takahashi, T.; Strittmatter, S.M. PlexinA1 Autoinhibition by the Plexin Sema Domain. Neuron 2001, 29, 429–439.
- Janssen, B.J.; Robinson, R.A.; Perez-Branguli, F.; Bell, C.H.; Mitchell, K.J.; Siebold, C.; Jones, E.Y. Structural basis of semaphorin-plexin signalling. Nature 2010, 467, 1118–1122.
- Nogi, T.; Yasui, N.; Mihara, E.; Matsunaga, Y.; Noda, M.; Yamashita, N.; Toyofuku, T.; Uchiyama, S.; Goshima, Y.; Kumanogoh, A.; et al. Structural basis for semaphorin signalling through the plexin receptor. Nature 2010, 467, 1123–1127.
- Artigiani, S.; Barberis, D.; Fazzari, P.; Longati, P.; Angelini, P.; van de Loo, J.W.; Comoglio, P.M.; Tamagnone, L. Functional regulation of semaphorin receptors by proprotein convertases. J. Biol. Chem. 2003, 278, 10094–10101.
- Siebold, C.; Jones, E.Y. Structural insights into semaphorins and their receptors. Semin. Cell Dev. Biol. 2012, 24, 139–145.
- Oinuma, I.; Ishikawa, Y.; Katoh, H.; Negishi, M. The Semaphorin 4D receptor Plexin-B1 is a GTPase activating protein for R-Ras. Science 2004, 305, 862–865.
- Oinuma, I.; Katoh, H.; Negishi, M. Molecular dissection of the semaphorin 4D receptor plexin-B1-stimulated R-Ras GTPase-activating protein activity and neurite remodeling in hippocampal neurons. J. Neurosci. 2004, 24, 11473–11480.
- Saito, Y.; Oinuma, I.; Fujimoto, S.; Negishi, M. Plexin-B1 is a GTPase activating protein for M-Ras, remodelling dendrite morphology. EMBO Rep. 2009, 10, 614–621.
- Wang, Y.; He, H.; Srivastava, N.; Vikarunnessa, S.; Chen, Y.B.; Jiang, J.; Cowan, C.W.; Zhang, X. Plexins Are GTPase-Activating Proteins for Rap and Are Activated by Induced Dimerization. Sci. Signal. 2012, 5, ra6.
- Uesugi, K.; Oinuma, I.; Katoh, H.; Negishi, M. Different requirement for Rnd GTPases of R-Ras GAP activity of plexin-C1 and plexin-D1. J. Biol. Chem. 2009, 284, 6743–6751.
- Tamagnone, L.; Artigiani, S.; Chen, H.; He, Z.; Ming, G.I.; Song, H.; Chedotal, A.; Winberg, M.L.; Goodman, C.S.; Poo, M.; et al. Plexins are a large family of receptors for transmembrane, secreted, and GPI-anchored semaphorins in vertebrates. Cell 1999, 99, 71–80.
- Tamagnone, L.; Comoglio, P.M. Signalling by semaphorin receptors: Cell guidance and beyond. Trends Cell Biol. 2000, 10, 377–383.
- Rohm, B.; Ottemeyer, A.; Lohrum, M.; Puschel, A.W. Plexin/neuropilin complexes mediate repulsion by the axonal guidance signal semaphorin 3A. Mech. Dev. 2000, 93, 95–104.
- Hayashi, M.; Nakashima, T.; Taniguchi, M.; Kodama, T.; Kumanogoh, A.; Takayanagi, H. Osteoprotection by semaphorin 3A. Nature 2012, 485, 69–74.
- Puschel, A.W. GTPases in semaphorin signaling. Adv. Exp. Med. Biol. 2007, 600, 12–23.
- Zanata, S.M.; Hovatta, I.; Rohm, B.; Puschel, A.W. Antagonistic effects of Rnd1 and RhoD GTPases regulate receptor activity in Semaphorin 3A-induced cytoskeletal collapse. J. Neurosci. 2002, 22, 471–477.
- Rajan, S.; Terman, J.R.; Reisler, E. MICAL-mediated oxidation of actin and its effects on cytoskeletal and cellular dynamics. Front. Cell Dev. Biol. 2023, 11, 1124202.
- Perrot, V.; Vazquez-Prado, J.; Gutkind, J.S. Plexin B regulates Rho through the guanine nucleotide exchange factors Leukemia-associated RhoGEF (LARG) and PDZ-RhoGEF. J. Biol. Chem. 2002, 278, 26111–26119.
- Oinuma, I.; Katoh, H.; Harada, A.; Negishi, M. Direct interaction of Rnd1 with Plexin-B1 regulates PDZ-RhoGEF-mediated Rho activation by Plexin-B1 and induces cell contraction in COS-7 cells. J. Biol. Chem. 2003, 278, 25671–25677.
- Swiercz, J.M.; Worzfeld, T.; Offermanns, S. Semaphorin 4D signaling requires the recruitment of phospholipase C gamma into the plexin-B1 receptor complex. Mol. Cell. Biol. 2009, 29, 6321–6334.
- Driessens, M.H.; Olivo, C.; Nagata, K.; Inagaki, M.; Collard, J.G. B plexins activate Rho through PDZ-RhoGEF. FEBS Lett. 2002, 529, 168–172.
- Worzfeld, T.; Swiercz, J.M.; Senturk, A.; Genz, B.; Korostylev, A.; Deng, S.; Xia, J.; Hoshino, M.; Epstein, J.A.; Chan, A.M.; et al. Genetic dissection of plexin signaling in vivo. Proc. Natl. Acad. Sci. USA 2014, 111, 2194–2199.
- Shalaby, M.A.; Hampson, L.; Oliver, A.; Hampson, I. Identification of PlexinD1 and AHDC1 as a putative interactors for Tip-1 protein. Genes Genom. 2011, 33, 399–405.
- Kigel, B.; Rabinowicz, N.; Varshavsky, A.; Kessler, O.; Neufeld, G. Plexin-A4 promotes tumor progression and tumor angiogenesis by enhancement of VEGF and bFGF signaling. Blood 2011, 118, 4285–4296.
- Takahashi, T.; Fournier, A.; Nakamura, F.; Wang, L.H.; Murakami, Y.; Kalb, R.G.; Fujisawa, H.; Strittmatter, S.M. Plexin-neuropilin-1 complexes form functional semaphorin-3A receptors. Cell 1999, 99, 59–69.
- Catalano, A.; Caprari, P.; Rodilossi, S.; Betta, P.; Castellucci, M.; Casazza, A.; Tamagnone, L.; Procopio, A. Cross-talk between vascular endothelial growth factor and semaphorin-3A pathway in the regulation of normal and malignant mesothelial cell proliferation. FASEB J. 2004, 18, 358–360.
- Tellman, T.V.; Cruz, L.A.; Grindel, B.J.; Farach-Carson, M.C. Cleavage of the Perlecan-Semaphorin 3A-Plexin A1-Neuropilin-1 (PSPN) Complex by Matrix Metalloproteinase 7/Matrilysin Triggers Prostate Cancer Cell Dyscohesion and Migration. Int. J. Mol. Sci. 2021, 22, 3218.
- Catalano, A.; Lazzarini, R.; Di, N.S.; Orciari, S.; Procopio, A. The Plexin-A1 Receptor Activates Vascular Endothelial Growth Factor-Receptor 2 and Nuclear Factor-kappaB to Mediate Survival and Anchorage-Independent Growth of Malignant Mesothelioma Cells. Cancer Res. 2009, 69, 1485–1493.
- Yamada, D.; Watanabe, S.; Kawahara, K.; Maeda, T. Plexin A1 signaling confers malignant phenotypes in lung cancer cells. Biochem. Biophys. Res. Commun. 2016, 480, 75–80.
- Higgins, D.M.O.; Caliva, M.; Schroeder, M.; Carlson, B.; Upadhyayula, P.S.; Milligan, B.D.; Cheshier, S.H.; Weissman, I.L.; Sarkaria, J.N.; Meyer, F.B.; et al. Semaphorin 3A mediated brain tumor stem cell proliferation and invasion in EGFRviii mutant gliomas. BMC Cancer 2020, 20, 1213.
- Tian, T.V.; Tomavo, N.; Huot, L.; Flourens, A.; Bonnelye, E.; Flajollet, S.; Hot, D.; Leroy, X.; de Launoit, Y.; Duterque-Coquillaud, M. Identification of novel TMPRSS2:ERG mechanisms in prostate cancer metastasis: Involvement of MMP9 and PLXNA2. Oncogene 2013, 33, 2204–2214.
- Yin, L.; Li, J.; Wang, J.; Pu, T.; Wei, J.; Li, Q.; Wu, B.J. MAOA promotes prostate cancer cell perineural invasion through SEMA3C/PlexinA2/NRP1-cMET signaling. Oncogene 2021, 40, 1362–1374.
- Shostak, K.; Zhang, X.; Hubert, P.; Goktuna, S.I.; Jiang, Z.; Klevernic, I.; Hildebrand, J.; Roncarati, P.; Hennuy, B.; Ladang, A.; et al. NF-kappaB-induced KIAA1199 promotes survival through EGFR signalling. Nat. Commun. 2014, 5, 5232.
- Toledano, S.; Sabag, A.D.; Ilan, N.; Liburkin-Dan, T.; Kessler, O.; Neufeld, G. Plexin-A2 enables the proliferation and the development of tumors from glioblastoma derived cells. Cell Death. Dis. 2023, 14, 41.
- Syed, V.; Zhang, X.; Lau, K.M.; Cheng, R.; Mukherjee, K.; Ho, S.M. Profiling estrogen-regulated gene expression changes in normal and malignant human ovarian surface epithelial cells. Oncogene 2005, 24, 8128–8143.
- Ding, H.; Li, L.; Gu, B.; Ni, Y.; Chen, S. MicroRNA-564 inhibits the progression of non-small cell lung cancer via targeting plexin A4. Exp. Ther. Med. 2021, 21, 358.
- Zhang, C.; Qiao, H.; Guo, W.; Liu, Y.; Yang, L.; Liu, Y.; Jin, B.; Fu, M.; Wang, G.; Li, W. CD100-Plexin-B1 induces Epithelial-Mesenchymal Transition of head and neck squamous cell carcinoma and promotes metastasis. Cancer Lett. 2019, 455, 1–13.
- Li, C.; Wan, L.; Wang, P.; Guan, X.; Li, C.; Wang, X. Sema4D/Plexin-B1 promotes the progression of osteosarcoma cells by activating Pyk2-PI3K-AKT pathway. J. Musculoskelet. Neuronal Interact. 2021, 21, 577–583.
- Peacock, J.W.; Takeuchi, A.; Hayashi, N.; Liu, L.; Tam, K.J.; Al, N.N.; Khazamipour, N.; Tombe, T.; Dejima, T.; Lee, K.C.; et al. SEMA3C drives cancer growth by transactivating multiple receptor tyrosine kinases via Plexin B1. EMBO Mol. Med. 2018, 10, 219–238.
- Liu, B.; Gu, X.; Huang, T.; Luan, Y.; Ding, X. Identification of TMPRSS2-ERG mechanisms in prostate cancer invasiveness: Involvement of MMP-9 and plexin B1. Oncol. Rep. 2017, 37, 201–208.
- Ye, S.; Hao, X.; Zhou, T.; Wu, M.; Wei, J.; Wang, Y.; Zhou, L.; Jiang, X.; Ji, L.; Chen, Y.; et al. Plexin-B1 silencing inhibits ovarian cancer cell migration and invasion. BMC Cancer 2010, 10, 611.
- Malik, M.F.A.; Riaz, S.K.; Waqar, S.H.; Haq, F.; Ye, L.; Jiang, W.G. Role of Plexin B1 in a Breast Cancer Cohort of Pakistani Patients and its Contribution Towards Cancer Metastasis as Indicated by an In Vitro Model. Anticancer Res. 2017, 37, 4483–4488.
- Argast, G.M.; Croy, C.H.; Couts, K.L.; Zhang, Z.; Litman, E.; Chan, D.C.; Ahn, N.G. Plexin B1 is repressed by oncogenic B-Raf signaling and functions as a tumor suppressor in melanoma cells. Oncogene 2009, 28, 2697–2709.
- Jiang, C.; Javed, A.; Kaiser, L.; Nava, M.M.; Xu, R.; Brandt, D.T.; Zhao, D.; Mayer, B.; Fernández-Baldovinos, J.; Zhou, L.; et al. Mechanochemical control of epidermal stem cell divisions by B-plexins. Nat. Commun. 2021, 12, 1308.
- Gurrapu, S.; Pupo, E.; Franzolin, G.; Lanzetti, L.; Tamagnone, L. Sema4C/PlexinB2 signaling controls breast cancer cell growth, hormonal dependence and tumorigenic potential. Cell Death Differ. 2018, 25, 1259–1275.
- Le, A.P.; Huang, Y.; Pingle, S.C.; Kesari, S.; Wang, H.; Yong, R.L.; Zou, H.; Friedel, R.H. Plexin-B2 promotes invasive growth of malignant glioma. Oncotarget 2015, 6, 7293–7304.
- Yu, W.; Goncalves, K.A.; Li, S.; Kishikawa, H.; Sun, G.; Yang, H.; Vanli, N.; Wu, Y.; Jiang, Y.; Hu, M.G.; et al. Plexin-B2 Mediates Physiologic and Pathologic Functions of Angiogenin. Cell 2017, 171, 849–864.
- Li, S.; Goncalves, K.A.; Lyu, B.; Yuan, L.; Hu, G.F. Chemosensitization of prostate cancer stem cells in mice by angiogenin and plexin-B2 inhibitors. Commun. Biol. 2020, 3, 26–0750.
- Yang, H.; Yuan, L.; Ibaragi, S.; Li, S.; Shapiro, R.; Vanli, N.; Goncalves, K.A.; Yu, W.; Kishikawa, H.; Jiang, Y.; et al. Angiogenin and plexin-B2 axis promotes glioblastoma progression by enhancing invasion, vascular association, proliferation and survival. Br. J. Cancer 2022, 127, 422–435.
- Liang, Y.; Meng, K.; Qiu, R. Circular RNA Circ_0013958 Functions as a Tumor Promoter in Ovarian Cancer by Regulating miR-637/PLXNB2 Axis. Front. Genet. 2021, 12, 644451.
- Brundu, S.; Napolitano, V.; Franzolin, G.; Lo, C.E.; Mastrantonio, R.; Sardo, G.; Cascardi, E.; Verginelli, F.; Sarnataro, S.; Gambardella, G.; et al. Mutated axon guidance gene PLXNB2 sustains growth and invasiveness of stem cells isolated from cancers of unknown primary. EMBO Mol. Med. 2023, 15, e16104.
- Li, X.; Law, J.W.; Lee, A.Y. Semaphorin 5A and plexin-B3 regulate human glioma cell motility and morphology through Rac1 and the actin cytoskeleton. Oncogene 2011, 31, 595–610.
- Saxena, S.; Prajapati, D.R.; Goel, P.; Tomar, B.; Hayashi, Y.; Atri, P.; Rachagani, S.; Grandgenett, P.M.; Hollingsworth, M.A.; Batra, S.K.; et al. Plexin-B3 Regulates Cellular Motility, Invasiveness, and Metastasis in Pancreatic Cancer. Cancers 2021, 13, 818.
- Kuhlmann, L.; Govindarajan, M.; Mejia-Guerrero, S.; Ignatchenko, V.; Liu, L.Y.; Grünwald, B.T.; Cruickshank, J.; Berman, H.; Khokha, R.; Kislinger, T. Glycoproteomics Identifies Plexin-B3 as a Targetable Cell Surface Protein Required for the Growth and Invasion of Triple-Negative Breast Cancer Cells. J. Proteome Res. 2022, 21, 2224–2236.
- Scott, G.A.; McClelland, L.A.; Fricke, A.F.; Fender, A. Plexin C1, A Receptor for Semaphorin 7A, Inactivates Cofilin and Is a Potential Tumor Suppressor for Melanoma Progression. J. Investig. Dermatol. 2009, 129, 954–963.
- Zhao, X.; Wang, P.; Liu, J.; Zheng, J.; Liu, Y.; Chen, J.; Xue, Y. Gas5 Exerts Tumor-suppressive Functions in Human Glioma Cells by Targeting miR-222. Mol. Ther. 2015, 23, 1899–1911.
- Ma, B.; Herzog, E.L.; Lee, C.G.; Peng, X.; Lee, C.M.; Chen, X.; Rockwell, S.; Koo, J.S.; Kluger, H.; Herbst, R.S.; et al. Role of Chitinase 3-like-1 and Semaphorin 7A in Pulmonary Melanoma Metastasis. Cancer Res. 2014, 75, 487–496.
- Chen, J.; Liu, H.; Chen, J.; Sun, B.; Wu, J.; Du, C. PLXNC1 Enhances Carcinogenesis Through Transcriptional Activation of IL6ST in Gastric Cancer. Front. Oncol. 2020, 10, 33.
- Casazza, A.; Finisguerra, V.; Capparuccia, L.; Camperi, A.; Swiercz, J.M.; Rizzolio, S.; Rolny, C.; Christensen, C.; Bertotti, A.; Sarotto, I.; et al. Sema3E-Plexin D1 signaling drives human cancer cell invasiveness and metastatic spreading in mice. J. Clin. Investig. 2010, 120, 2684–2698.
- Casazza, A.; Kigel, B.; Maione, F.; Capparuccia, L.; Kessler, O.; Giraudo, E.; Mazzone, M.; Neufeld, G.; Tamagnone, L. Tumour growth inhibition and anti-metastatic activity of a mutated furin-resistant Semaphorin 3E isoform. EMBO Mol. Med. 2012, 4, 234–250.
- Luchino, J.; Hocine, M.; Amoureux, M.C.; Gibert, B.; Bernet, A.; Royet, A.; Treilleux, I.; Lecine, P.; Borg, J.P.; Mehlen, P.; et al. Semaphorin 3E Suppresses Tumor Cell Death Triggered by the Plexin D1 Dependence Receptor in Metastatic Breast Cancers. Cancer Cell 2013, 24, 673–685.
- Man, J.; Shoemake, J.; Zhou, W.; Fang, X.; Wu, Q.; Rizzo, A.; Prayson, R.; Bao, S.; Rich, J.N.; Yu, J.S. Sema3C promotes the survival and tumorigenicity of glioma stem cells through Rac1 activation. Cell Rep. 2014, 9, 1812–1826.
- Liu, H.; Juo, Z.S.; Shim, A.H.; Focia, P.J.; Chen, X.; Garcia, K.C.; He, X. Structural Basis of Semaphorin-Plexin Recognition and Viral Mimicry from Sema7A and A39R Complexes with PlexinC1. Cell 2010, 142, 749–761.
- Odabas, G.; Cetin, M.; Turhal, S.; Baloglu, H.; Sayan, A.E.; Yagci, T. Plexin C1 Marks Liver Cancer Cells with Epithelial Phenotype and Is Overexpressed in Hepatocellular Carcinoma. Can. J. Gastroenterol. Hepatol. 2018, 2018, 4040787.
- NazimTurhal, S.; Dogan, M.; Esendagli, G.; Artac, M.; Korkmaz, L.; Coskun, H.S.; Goker, E.; PerranYumuk, F.; Bilgetekin, I.; Kose, F.; et al. The Relationship Between Plexin C1 Overexpression and Survival in Hepatocellular Carcinoma: A Turkish Oncology Group (TOG) Study. J. Gastrointest. Cancer 2021, 53, 356–362.
- Ni, Z.; Huang, C.; Zhao, H.; Zhou, J.; Hu, M.; Chen, Q.; Ge, B.; Huang, Q. PLXNC1: A Novel Potential Immune-Related Target for Stomach Adenocarcinoma. Front. Cell Dev. Biol. 2021, 9, 662707.
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13.
- Roodink, I.; Raats, J.; Van Der, Z.B.; Verrijp, K.; Kusters, B.; Van Bokhoven, H.; Linkels, M.; de Waal, R.M.; Leenders, W.P. Plexin d1 expression is induced on tumor vasculature and tumor cells: A novel target for diagnosis and therapy? Cancer Res. 2005, 65, 8317–8323.
- Vivekanandhan, S.; Mukhopadhyay, D. Divergent roles of Plexin D1 in Cancer. Biochim. Biophys. Acta Rev. Cancer 2019, 1872, 103–110.
- Gu, C.; Yoshida, Y.; Livet, J.; Reimert, D.V.; Mann, F.; Merte, J.; Henderson, C.E.; Jessell, T.M.; Kolodkin, A.L.; Ginty, D.D. Semaphorin 3E and plexin-D1 control vascular pattern independently of neuropilins. Science 2005, 307, 265–268.
- Yang, W.J.; Hu, J.; Uemura, A.; Tetzlaff, F.; Augustin, H.G.; Fischer, A. Semaphorin-3C signals through Neuropilin-1 and PlexinD1 receptors to inhibit pathological angiogenesis. EMBO Mol. Med. 2015, 20, 1267–1284.
- Smolkin, T.; Nir-Zvi, I.; Duvshani, N.; Mumblat, Y.; Kessler, O.; Neufeld, G. Complexes of plexin-A4 and plexin-D1 convey semaphorin-3C signals to induce cytoskeletal collapse in the absence of neuropilins. J. Cell Sci. 2018, 131, jcs208298.
- Mumblat, Y.; Kessler, O.; Ilan, N.; Neufeld, G. Full length semaphorin-3C functions as an inhibitor of tumor lymphangiogenesis and tumor metastasis. Cancer Res. 2015, 75, 2177–2186.
- Bassi, D.E.; Fu, J.; de Lopez, C.R.; Klein-Szanto, A.J. Proprotein convertases: “master switches” in the regulation of tumor growth and progression. Mol. Carcinog. 2005, 44, 151–161.
- Shalaby, M.A.; Hampson, L.; Oliver, A.; Hampson, I. Plexin d1: New potential biomarker for cervical cancer. J. Immunoass. Immunochem. 2012, 33, 223–233.

