1. TGM Immobilization
The immobilization of transglutaminase (TGM) on solid supports is a widely used method to increase TGM stability and improve its spectrum of use and reuse. Site-specific modification is needed for such applications, especially when unstable targets are in question. Harsh conditions are usually applied during chemical immobilization. If compared to enzymatic catalysis, selective and fast performance in aqueous media is performed under mild process conditions, which has many advantages. Immobilizing the enzyme onto a given matrix is benefiting the enzyme with a stabile structure, which provides an advantage in resisting temperature and pH alterations by retaining catalytic activities
[1][2][3][4]. Immobilized enzymes are more stable and have improved properties and features in terms of their kinetic aspects when compared to the free form of the enzyme. The enzymes’ improved features are due to the conformational changes that happen in the enzyme structure as a result of the chosen and most suitable immobilization method. In this manner, enhanced activity, stability, and selectivity can be observed
[5][6][7][8]. For larger-scale applications, immobilized enzymes are the considered subject of choice. Not many studies were reported on immobilizing TGM. However, immobilized TGM was investigated on a few supports, such as thermo-responsive carboxylated poly (N-isopolylacrylamide), agarose beads, polypropylene microporous membranes, and various nanomaterials, such as magnetic nanoparticles (MNPs) or carbon nanotubes (CNTs), as well as in the form of crosslinked enzymes aggregates (CLEAs). For example, TGM was immobilized on multi-walled CNTs for tissue scaffold designing. The highest immobilization efficiency of 58% was achieved and a 4.8 fold increase in catalytic efficiency was observed
[9]. Gianetto et al. reported on a new amperometric immunosensor, which was based on the covalent immobilization of TGM onto functionalized gold nanoparticles, and used for the determination of anti-tissue TGM antibodies in human serum
[10]. Another piezoelectric immunosensor was developed for the detection of anti-tissue TGM antibodies as specific biomarkers for early diagnosis of celiac disease by Manfredi et al.
[11]. Leitgeb et al. studied and investigated the immobilization of TGM onto surface-modified MNPs with carboxymethyl dextran for cleaner production technologies. In this case, the TGM was hyperactivated and exhibited 99% immobilization efficiency with 110% residual activity. It showed excellent thermal stability at 50 °C and at 70 °C
[12]. Another TGM investigation by the same group reported on the synthesis of TGM immobilized in the form of CLEAs and magnetic CLEAs (mCLEAs). The TGM was precipitated in 2-propanol and later crosslinked with glutaraldehyde (GA), which resulted in 63% and 73% of residual activity for TGM CLEAs and mCLEAs, respectively. The CLEAs and mCLEAs showed great immobilization efficiency as well (95% and 90% for CLEAs and mCLEAs, respectively)
[13]. Zhou et al. reported on TGM immobilized covalently on thermo-responsive carboxylated poly(N-isopropylacrylamide), where the immobilized TGM exhibited reversible solubility in an aqueous solution with a low critical solution temperature of 39 °C. Such immobilized TGM can be used to modify proteins in food processing and biomedical engineering
[14]. The Wen-qiong report showed immobilization of TGM on an ultrafiltration polyethersulfone membrane surface, where it retained 50% of residual activity after 20 days. Additionally, the TGM-immobilized membrane had a higher relative membrane flux of 0.15 MPa in a membrane reactor
[15].
2. Food Related Industries
As proteins are important food components, which play an important role in the phyisicochemical properties of food, the usage of TGM to crosslink food proteins to change their functional characteristics has been in progress for more than 30 years
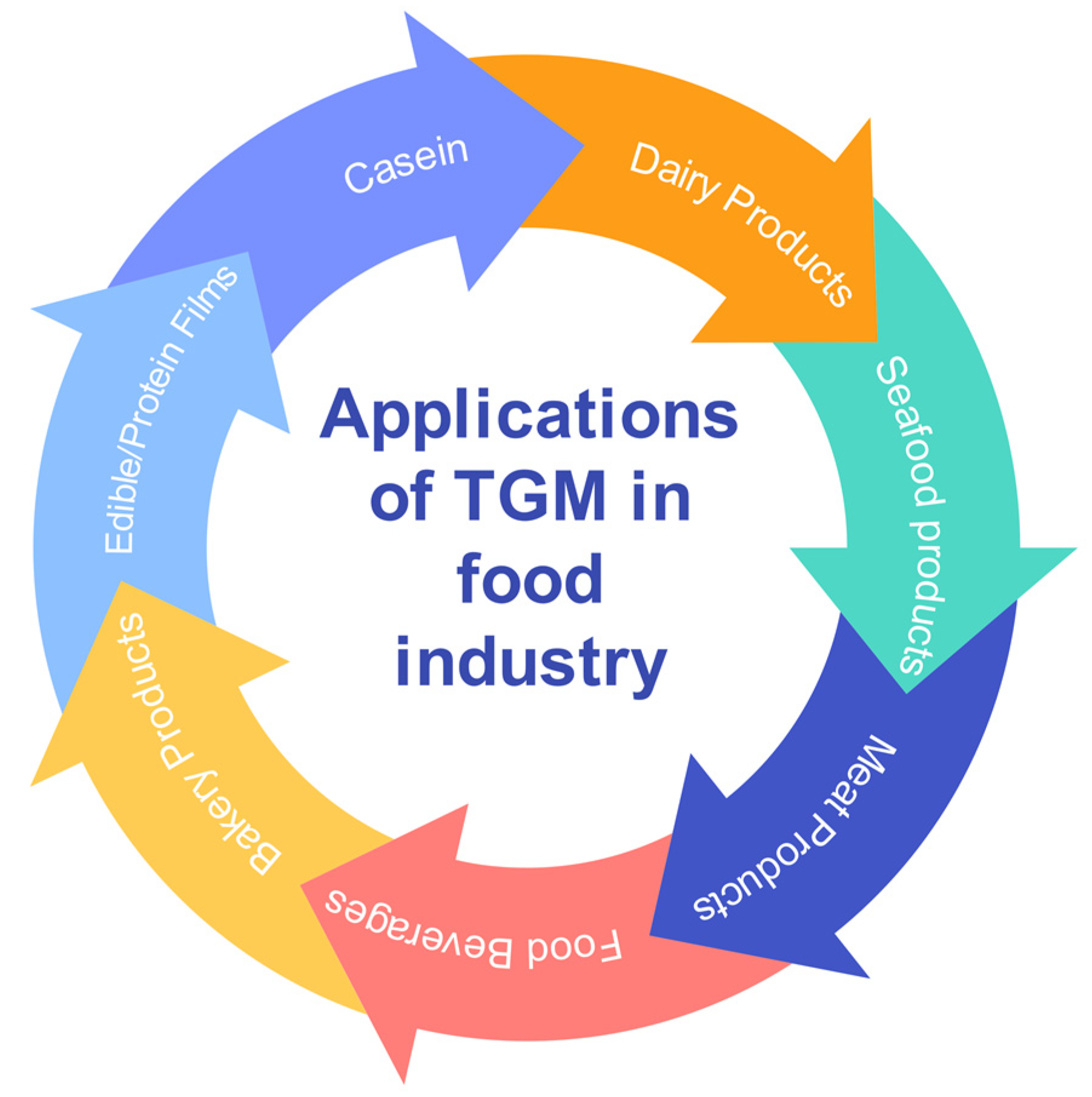
[16][17][18]. Enzymatic preparations of TGM have an important role in the food industry due to their practical utilization. Many reports describe the use of TGM in various food-related industries for the crosslinking of proteins, as in meat, cheese, yogurt, or bread (
Figure 1).
Figure 1. Opportunities for applications of TGM in the food industry.
It can also be used to produce composite edible films. TGM catalyzes the formation of crosslinks within a molecule as well as between molecules of other proteins. This feature impacts the changes in protein functionalities, such as solubility, foaming, emulsifying capacity, and gelation. As TGM has broad substrate specificity,
Table 1 shows the reactivity of microbial TGM, which was investigated on different types of proteins that were derived from various foods
[19][20][21][22].
Table 1. Reactivity of microbial TGM in relation to different food proteins.
2.1. Dairy Industry
TGM uses proteins such as casein or whey proteins as substrates to improve the foaming, emulsifying, and gelling properties of different foods. Casein is a major milk protein, which is a great substrate for TGM in dairy products due to its low degree of tertiary structure, flexibility, and the absence of disulfide bonds, which allows the exposure of reactive groups to TGM. On the other hand, the globular whey proteins, which do contain disulfide bonds, are poor substrates for TGM in the crosslinking process and therefore require modifications. Such modifications can be performed with reducing agents or increasing the pH value. They can also be achieved by heat denaturation or application of high hydrostatic pressure. However, such alterations in treatment can affect the interactions between the enzyme TGM in the TGM inhibitors that are present in milk serum and can also induce denaturation and result in cleavage of disulfide bonds, which later leads to unfolding of the proteins
[23]. For example, the enzymatic crosslinking of casein was more resistant to digestion in comparison to the non-crosslinked casein. This suggests that the development of new types of products can offer carious food with improved structural characteristics, such as the polymerization of milk proteins with TGM results in protein film formation, which improves the functional properties of dairy products
[24][25].
In the dairy industry, TGM has been introduced in many products. In yoghurts, it is used to prevent syneresis and for texture firming or softening since TGM-modified casein allows the manufacture of dairy products with a more consistent structure. The result of this reduced syneresis is a firmer and more homogeneous product. Various methods and protocols were also carried out investigating the use of TGM to increase cheese yield while enhancing the quality of low-fat cheese. TGM is also used in cheese manufacturing, where three methods are performed, including TGM:
- -
-
the addition of TGM to milk, followed by heating for pasteurization and deactivation of enzymes, concluded with the addition of rennet to the milk;
- -
-
the addition of rennet to the milk, followed by the addition of TGM;
- -
-
the addition of TGM and rennet at the same time.
However, the reported investigations confirmed that the addition of TGM before the rennet prevented the coagulation of milk, while the simultaneous addition of both resulted in reduced resistance and hardness of the cheese
[26]. By improving the cheese yield, textural properties, and its water-holding capacity, the use of TGM in cheese production is crucial. Other investigations reported on the improved heat stability and consistency of processed cheese after implementing TGM into the production
[27][28][29][30][31].
Microbial TGM was used to treat the rheological and microstructural properties of yoghurt, where it was applied to milk before the fermentation. The TGM-mediated treatment decreased the ropiness of yoghurts and contributed to the acceptability of their texture, a study by Marhons suggests
[32]. Salunke et al. investigated the use of micellar casein concentrate and milk proteins that were treated with TGM in different imitation cheese products. As TGM has the potential to modify the surface properties of milk protein concentrate and micellar casein concentrate, it may also improve functionality in imitation cheese, such as mozzarella
[33]. Another study by the same group investigated the melt and stretch properties of dairy-based imitation mozzarella cheese, where the effect was studied of TGM-treated concentrates. The results demonstrated that TGM treatment modifies the investigated stretch and melt functionalities of milk protein concentrate and micellar casein concentrate
[34]. Another study by Monsalve-Atencio et al. investigated the effect of TGM and its interaction with another enzyme, phospholipase, on the composition, yield, texture, and microstructure of semi-soft fresh cheese. The interaction of TGM with phospholipase showed the highest content of moisture in cheese value, which suggests an economically improved application of TGM in cheesemaking
[35]. The addition of microbial TGM in quark cheese was studied, where the physicochemical, textural, sensory, and microbial properties of cheese were studied as well
[36].
2.2. Baking Industry
The use of TGM in the baking industry is improving the quality of flour, and, consequently, the texture and volume of bread as proteins from grains are good substrates for crosslinkers by TGM. For example, rice flour is known to contain valuable nutrients, such as proteins, vitamins B and E, as well as fiber. However, it can only be used in and is limited to non-fermented bakery products. Investigations showed that the addition of TGM to rice flour improved the rheological properties of dough, and, by that, increased the content of triglycerides
[37][38][39][40]. Similar studies were reported concerning cassava and wheat flour
[41][42]. A TGM-induced protein aggregation method to improve the baking properties was investigated by Beck et al.
[43], where the effect was studied of microbial TGM on the properties of rye dough. It was reported that the addition of TGM modified the rheological properties of rye flour dough, which resulted in a progressive increase in shear modulus. The increased TG concentration also showed an increase in crumb springiness and hardness, which demonstrated the improved breadmaking with the use of TGM. Another study also reports on improved rheological properties of gluten-free batter with the implementation of TGM. The crumb properties revealed that increased TGM concentration increased crumb chewiness and firmness
[44]. The use of varying amounts of TGM also improved the baking quality of high-level sun pest wheat, where it was observed that TGM plays an important role in the baking quality. Increasing TGM activity caused increased bread characteristics of wheat, such as bread yield, height, crumb softness, pore structure, as well as decreased weight loss and wideness of the bread samples. The study concluded that the addition of TGM can restore the properties of bread and improve its overall protein structure
[45]. Lang et al. evaluated the influence of TGM on the technological properties of gluten-free cakes of brown, black, and red rice. The effect of baking on the phenolic compound content was investigated as well
[39].
2.3. Meat Industry
Numerous reports and studies are available investigating the use of TGM in meat products as one of the most widespread applications of TGM is in the restructuring of meat. Despite improving the structure and texture of the meat product, the use of TGM also provides cohesion without thermal processing or any additives, such as phosphates
[46][47][48][49][50]. Studies show that the crosslinking activity of TGM in meat depends strongly on the temperature, pH, protein surface charge, and ionic strength. It was shown to improve other characteristics as well, such as water-binding, gelation, and emulsion stability. With the use of TGM in meat production, the secondary structure of a myosin heavy chain is changed by reduced α-helix content and increasing β-sheet content, which results in the formation of high-molecular-weight polymers. With such structural modifications, strong gels were formed with compact structural properties, which allow cohesiveness and improve the hardness of the meat. In addition, some studies also show that different degrees of gelation can be observed when induced by the addition of TGM
[51][52][53]. This is also valid for fish skin gelatin. Namely, TGM-modified cold-water fish skin gelatin could be a potential mammalian gelatin replacer
[54]. The addition of TGM also improves the quality of collagen, blood proteins, and provides higher nutritional value by supplementing with respective amino acids, such as exogenous lysine. TGM has allowed the production of new meat products, which use lower-quality raw materials instead of high-value meat products. The impact of TGM on the protein of such raw materials (skimmed milk powder or soy powder) does not alter the appearance, smell, texture, or nutritional value from products made with high-quality meat
[55]. A study by Ribeiro et al. produced bovine meat with different levels of TGM combined with papain. The effects were investigated on pH, water activity, instrumental color, proximate composition, texture, and yield. It was concluded that the addition of TGM increased the yield of meat loafs
[52]. In an interesting study by Wen et al., the enzyme TGM was used to develop 3D-printable meat analogs that imitate the physiochemical properties of beef. The TGM improved the rheological properties of raw meat, and provided a method for modifying the texture of meat analogs using TGM catalysis
[56].
As the majority of the population are omnivores, TGM is being used widely in plant-based (PB) food industries, which are designing PB products that mimic the look, taste, and feel of animal-sourced foods. Zhou et al. developed PB protein gels for meat analogues that are created using slats, polysaccharides, and crosslinking enzymes, such as TGM, to modulate their gelation and assembly properties. In their study, the TGM increased the gel strength by forming covalent crosslinks between the potato protein molecules with more meat-like structures
[57]. Additionally, traditional sausage production technologies can be used for PB analogues, such as a PB salami-type sausage analogue, which was manufactured with TGM-mediated soy protein isolate gels as binders, investigated in a study by Herz et al.
[58].
In addition, the use of TGM is gaining much attention in the field of food packaging products; e.g., hemp proteins were used as raw material to obtain biodegradable films since they were demonstrated to act as both acyl donor and acceptor substrates of microbial TGM crosslinking
[59][60]. Because such bioplastics show higher gas permeability and greater hydrophobicity, they may be useful as packaging systems for protecting food products from physical contamination and, thus, for extending their shelf-life. In the crosslinked gelatin-based films, food preservatives such as lysozyme or nisin may be incorporated to extend the shelf life of perishable foods. It was demonstrated that microbial-TGM-crosslinked gelatin-based films incorporated with lysozyme can control the release of this food preservative effectively.
3. Biotechnology and Cosmetics
Microbial TGM is an interesting tool for protein modification, which catalyzes protein crosslinking through isopeptide bond formation, which occurs between γ-carboxamide groups of glutamine residues, which include the acyl donor, and primary amines, such as ε-amino groups of lysine residues, which include the acyl acceptor. Therefore, research on TGM use can be applied to biomedical, biomaterial, cosmetic, and feedstock technologies. TGM can be used for modification of gelatin hydrogels and collagen for enhancing binding in different tissues. Microbial TGM was used to prepare collagen-grafted chitosan, which could serve not only to reduce the loss of moisture but also to absorb the moisture. With such properties, it showed the potentiality to repair skin in the cosmetic, biomedical, and pharmaceutical fields
[61]. TGM-crosslinked whey proteins were also used to prepare a D-limonene emulsion, which can solve the problems of easy oxidation and poor water solubility of D-limonene. Limonene is an important ingredient in the formulation of different cosmetic and personal care products, such as aftershave lotions, bath products, cleansing products, eye shadows, hair products, lipsticks, shampoos, etc.
[62]. Moreover, microbial TGM, as a protein-crosslinking enzyme in the processing of hair, improved the rigidity of hair fibers by 15.64% compared to a control when it was applied to damaged hair
[63].
It has also been applied for bioconjugation, in order to create antibody–drug conjugates for various therapeutic applications. Additionally, new uses for TGM in the field of novel biomaterials are suggested and can be generated via site-specific substrate binding by proteins modified by TGM
[64][65]. Regarding different feedstocks, TGM has shown to improve the physical properties of fish feed, while, in cosmetics, the TGM-catalyzed reactions between amino groups of starch and γ-carboxamide groups of collagen peptides were investigated and reported to increase the effectiveness of some synthesized materials, such as drugs
[61][66][67].
Fusion proteins with dual functions are important for immunochemical assays. Among such assays are the enzyme-linked immunosorbent assay (ELISA) and Western blot assays. In that manner, genetic fusion provides poor yields when large-sized hybrid molecules are assembled. Therefore, TGM catalysis is considered as a method for the preparation of protein–protein conjugates. It was demonstrated that the coupling of two functional proteins, namely peroxidase and protein G, is possible through lysine and glutamine active sites. As a result, only a small amount of the desired conjugate was yielded. Later, when TGM-mediated conjugation was performed, only the desired conjugates were obtained
[68][69]. This finding provided TGM the recognition that respective tags can be applied to recombinant production to terminal and internal sites. Therefore, acyl donor and acceptor incorporation enable covalent linkage in the monomeric subunits, which enhances the thermal stability of the dimer
[70]. Native antibody site-specific modification enhances the properties of antibody-based bioconjugates. However, such antibodies have a single functionality. A work by Walker et al. addressed this limitation by designing heterofunctional substrates for microbial TGM that can contain both azide and methyltetrazine “click handles”, which present a powerful method in the toolbox for native antibody modification
[71][72][73][74][75]. Antibody–drug conjugates for cancer treatment have placed site-specific TGM-catalyzed conjugation with cytotoxic properties at the very pinnacle of research. TGM’s remarkable properties make it a versatile tool for post-translational modification of various proteins (
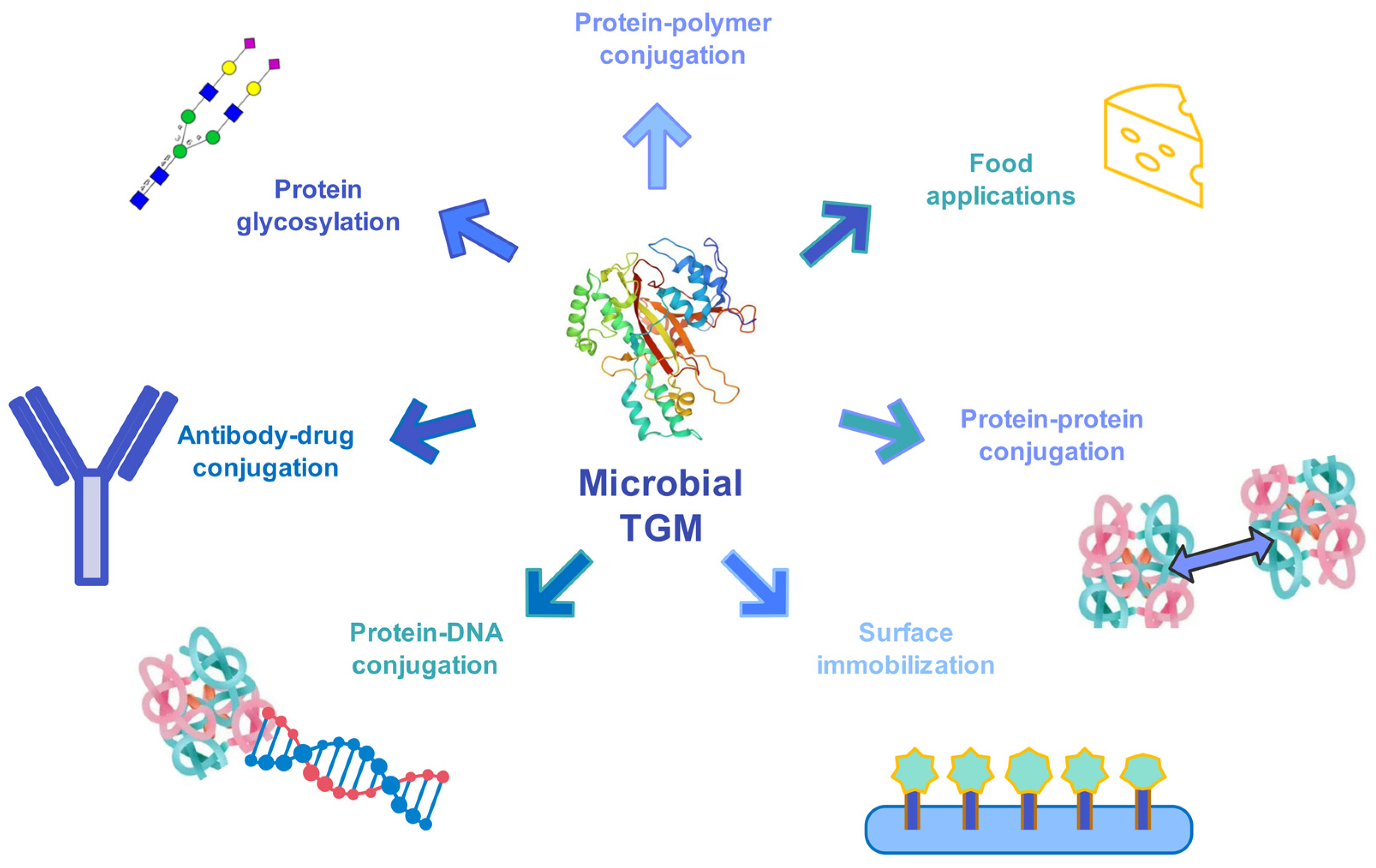
Figure 2). A few examples are PEGylation of small-protein drugs to elevate their half-life, or immobilization of biocatalysts that are prone to aggregation in order to increase their stability or covalent attachment of nucleic acids to proteins for combining the properties of both biomolecules
[76][77][78][79]. The reactivity of microbial TGM was investigated at intrinsic lysine and glutamine sites of different antibodies
[72][75][80][81][82][83]. The amino component in N-terminal pentaglycyl was found to mediate protein modification by TGM, which was performed by crosslinking of the enhanced green fluorescent protein. Many other protein–protein conjugates were performed using TGM-catalyzed conjugation. As reported by Bhokisham et al., attachment on solid supports occurred, which caused optimization of the reaction stoichiometry
[84].
Figure 2. Biotechnological applications of microbial TGM—an overview.
Covalent coupling of nucleic acid macromolecules, such as DNA and RNA, to proteins is a powerful method in molecular biology since both components exhibit excellent functions
[85][86][87]. Therefore, protein–oligonucleotide conjugates became valuable tools in analytic and biomedical applications, for example, in drug delivery
[88] and molecular diagnostics
[89].
Protein–oligonucleotide ligation often relies on chemical methods that include functional groups of the desired target protein. As a result, partial functional loss may occur due to the steric hindrance or protein treatment with organic solvents. Therefore, site-specific coupling is performed to avoid residue modification, which is essential for protein function. In that manner, innovative strategies were developed to include microbial TGM in the modification process. Some research reports show aminated DNA, which was attached chemically to the acyl donor substrate and further coupled to alkaline phosphatase
[90].
A modern technique to improve the pharmacokinetic properties of pharmacophore is PEGylation, where polyethylene glycol is attached to small-protein-based drugs. PEGylated pharmaceutics usually reduce immunogenicity compared to the non-PEGylated counterparts due to the enlarged hydrodynamic radius and higher conformational stability
[91]. With increased bioavailability, the intravenous administration of such protein-based drugs can become more patient-friendly. The controlled conjugation strategies are very important since the modifications at binding interfaces or active sites of residues can affect the in vivo functions. Leading to undesired reactivity during conjugation, it can lead to a heterogeneous mixture of products with changed pharmacokinetic properties. Nevertheless, PEGylated proteins that are synthesized by chemical derivatization are already available on the market
[91]. However, compared to chemical modification of such proteins, the site selectivity of microbial TGM is accessing PEGylated derivatives of various drugs without altering their properties
[92].
Another way to manipulate specificity is covalent immobilization of TGM on solid supports. In a study by Grigoletto et al., TGM was coupled to agarose beads through an N-terminus to investigate its activity and substrate specificity. The immobilized TGM exhibited changed enzymatic activity and kinetic parameters, which were the result of chemical modification. Due to PEGylation, the immobilized TGM appeared more site-selective
[76]. A biodegradable alternative to PEG, hydroxyethyl starch, is a polymeric molecule, which was used for microbial TGM-mediated protein conjugation and served as an acyl donor/acceptor to ligate monodansyl cadaverine
[93]. Moreover, microbial TGM was also proved to be able to catalyze an acyltransfer reaction between the aminated oligosaccharides and acyl donor molecules
[94], while glycosylation of catalase and trypsin was obtained by the transamidation of carboxamide functions.
Antibody–drug conjugates are promising tools for tumor treatments, where a chemotherapeutic is bound covalently to immunoglobulin, with the intent to enlarge the therapeutic range with the combination of powerful organic toxins and targeted specificity antibodies. Cytotoxic drugs are used widely to treat malignancies and solid tumors, and have, under specific clinical conditions, altered the natural course of certain diseases. Due to their intrinsic mode of active site, they are effective but can also cause significant on-target events that could result in the discontinuation of medication, which would increase the risk of recurrence of the tumor. For maintaining the efficiency of chemotherapeutics, efforts were undertaken to investigate novel approaches. Among such approaches was the conjugation of cytotoxic agents to antibodies. Many studies describe improved pharmacokinetics, enhanced efficacy, and reduced toxicity of antibody–drug conjugates when they are ligated site-selectively
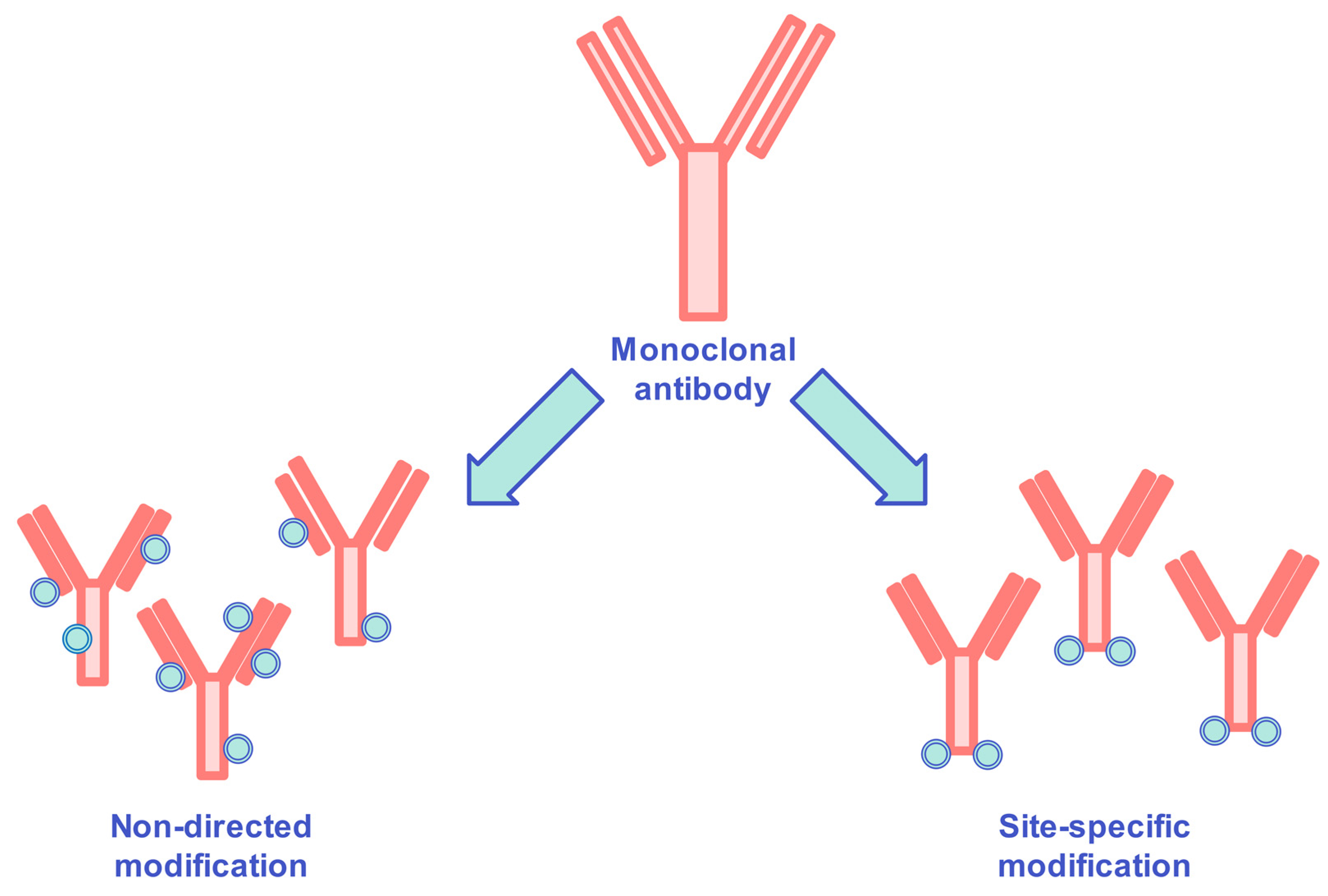
[95]. Non-directional methods generate heterogeneous products with statistically distributed coupling sites and fluctuating hydrophobic profiles, while site-specific conjugation provides reproducible hydrophobic properties with antibody modification restrictions (
Figure 3).
Figure 3. Schematic variations of non-directed modification and site-specific modification of antibody–drug conjugates.
Most of the antibody–drug conjugates in clinical use nowadays are assembled by random modification of cysteines and lysines. Many studies where an enzymatic approach was developed to modify antibodies site-selectively were reported
[96][97][98][99][100][101]. Such an approach includes additional cysteine or selenocysteine residues
[102][103]. Microbial TGM shows incapability of labelling glutamines in native human antibodies efficiently, although many Gln sites are exposed and available
[104]. Therefore, a specific recognition tag is being incorporated and is reported to be a powerful strategy in overcoming such limitations. Research was performed at Pfizer, where a TGM recognition sequence was placed at various surface-exposed regions of a specific growth factor receptor antibody, which facilitated efficient labeling by microbial TGM
[70].
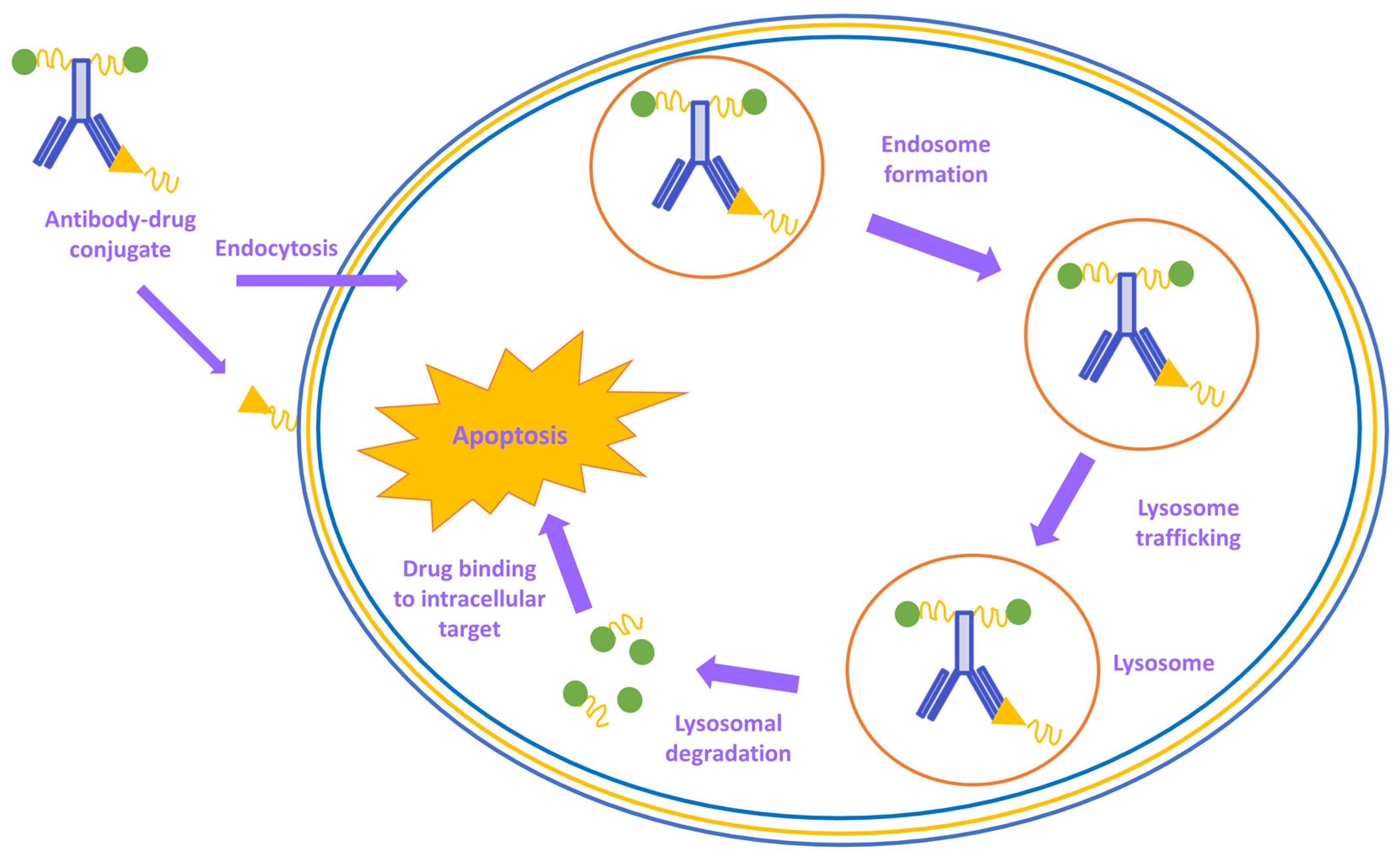
In the synthesis process of an antibody–drug conjugate, all aspects must be taken into consideration. The target, antigen, antibody, linker, and the cytotoxic load must be evaluated for the targeted cancer indication. The antibodies with cytotoxic load must obtain excellent targeting capabilities to distinguish between healthy and tumor cells. In the process, antibodies that are engineered to follow a specific tumor antigen attach themselves to the surface of tumor cells. When processing within lysosomes or endosomes takes place, the antibody–drug conjugate releases its lethal load and destroys the targeted tumor cells (
Figure 4). Due to its highly targeted tumor antigen expression, recognition, and requirements for effective internalization and processing, the antibody–drug conjugates are believed to provide a broader therapeutic opportunity in the ever-growing field of enzyme-mediated modification strategies
[73].
Figure 8. Schematic biological activity mechanism of antibody–drug conjugates.
 +1 credit
+1 credit