Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Djamel Drider | -- | 1803 | 2023-07-24 11:31:00 | | | |
| 2 | Jessie Wu | -3 word(s) | 1800 | 2023-07-25 05:31:21 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Ladjouzi, R.; Dussert, E.; Teiar, R.; Belguesmia, Y.; Drider, D. Enterocin DD14. Encyclopedia. Available online: https://encyclopedia.pub/entry/47176 (accessed on 22 July 2026).
Ladjouzi R, Dussert E, Teiar R, Belguesmia Y, Drider D. Enterocin DD14. Encyclopedia. Available at: https://encyclopedia.pub/entry/47176. Accessed July 22, 2026.
Ladjouzi, Rabia, Elodie Dussert, Radja Teiar, Yanath Belguesmia, Djamel Drider. "Enterocin DD14" Encyclopedia, https://encyclopedia.pub/entry/47176 (accessed July 22, 2026).
Ladjouzi, R., Dussert, E., Teiar, R., Belguesmia, Y., & Drider, D. (2023, July 24). Enterocin DD14. In Encyclopedia. https://encyclopedia.pub/entry/47176
Ladjouzi, Rabia, et al. "Enterocin DD14." Encyclopedia. Web. 24 July, 2023.
Copy Citation
Enterocin DD14 (EntDD14) is a two-peptide leaderless bacteriocin (LLB) produced by Enterococcus faecalis 14, a human strain isolated from meconium. Studies performed on EntDD14 enabled it to show its activity against Gram-positive bacteria such as Listeria monocytogenes, Clostridium perfringens, Enterococcus faecalis, and Staphylococcus aureus.
Enterocin DD14 (EntDD14)
leaderless bacteriocin (LLB)
specific transport
antibacterial and antiviral properties
DdA
1. The Genetic and Synthesis Backgrounds of Enterocin DD14
The EntDD14 cluster encompasses 10 ORFs, namely ddABCDEFGHIJ [1]. Most of the genes present in the EntDD14 cluster are adjacent or overlap and in silico analysis suggested the presence of at least two operons. Thus, two putative transcriptional promoters and two putative terminators were detected. The first operon comprises ddA and ddB genes encoding the two-peptide LLB (DdA and DdB). Genes ddA and ddB are co-transcribed and their transcription is under the control of the putative P1 promoter, which is located upstream of the ddA gene (the P1 promoter presented a −35 (TTGAtA) and −10 (aATAAT) regions). The second contains the remaining genes ddCDEFGHIJ which encode two unknown proteins (DdC, D), membrane proteins YdbS and YdbT containing PHb2 domain DdE, F, and the ABC transporter (DdG, H, I and J) [1]. The transcription of this second operon seems to be dependent on the putative P2 promoter located upstream of the ddC gene (the P2 promoter presented a −35 (TTGttA) and a −10 (TATAtT) regions). The two putative ρ-independent transcriptional terminators designated T1 and T2 were identified between the 3’-end of ddB and the P2 promoter for T1 and the T2 at 163 nt downstream of the 3-end of the ddJ gene.
To gain insights into the transport of EntDD14 and immunity mechanisms of E. faecalis 14, different derivative strains were constructed and their phenotype has been examined. The data gathered enabled researchers to suggest the model depicted in Figure 1. Transport of EntDD14 outside the cell is essentially ensured by a channel formed by DdE and DdF proteins [2]. This channel is used as a primary non-ATP-dependent transport to externalize EntDD14, until a certain threshold of extracellular bacteriocin concentration (Figure 1A), the bacteriocinogenic strain switches to the ABC transporter composed of DdGHIJ, which is an ATP-dependent transport. This ABC system requires ATP to transport the bacteriocin in the opposite of the concentration gradient. These two transport systems, namely DdEF and DdGHIJ, and ABC transporters (DdGHIJ) should interact synergistically to control the flow of bacteriocin, through a mechanism that needs to be studied. When DdE and/or DdF proteins are not produced (Figure 1B), the bacteriocinogenic strain was unable to export EntDD14 outside of the cytoplasm, despite the expression of genes coding for the ABC transporters and those coding for active EntDD14 (ddAB). The intracellular accumulation of EntDD14 was to be toxic for the bacteriocinogenic strain as a loss of 1 log viability was registered in a mutant deficient in ddEF [3]. When the ABC transporter system (DdGHIJ) was altered by deleting one or more components (Figure 1B), the bacteriocinogenic strain was still able to produce EntDD14, but at a lower rate than the wild-type [1][2]. These data indicate the role of ABC transporter in the transport of EntDD14, but not as the essential transporter. Nonetheless, researchers have also established its involvement in the protection of the bacteriocinogenic strain against extracellular EntDD14 [2].
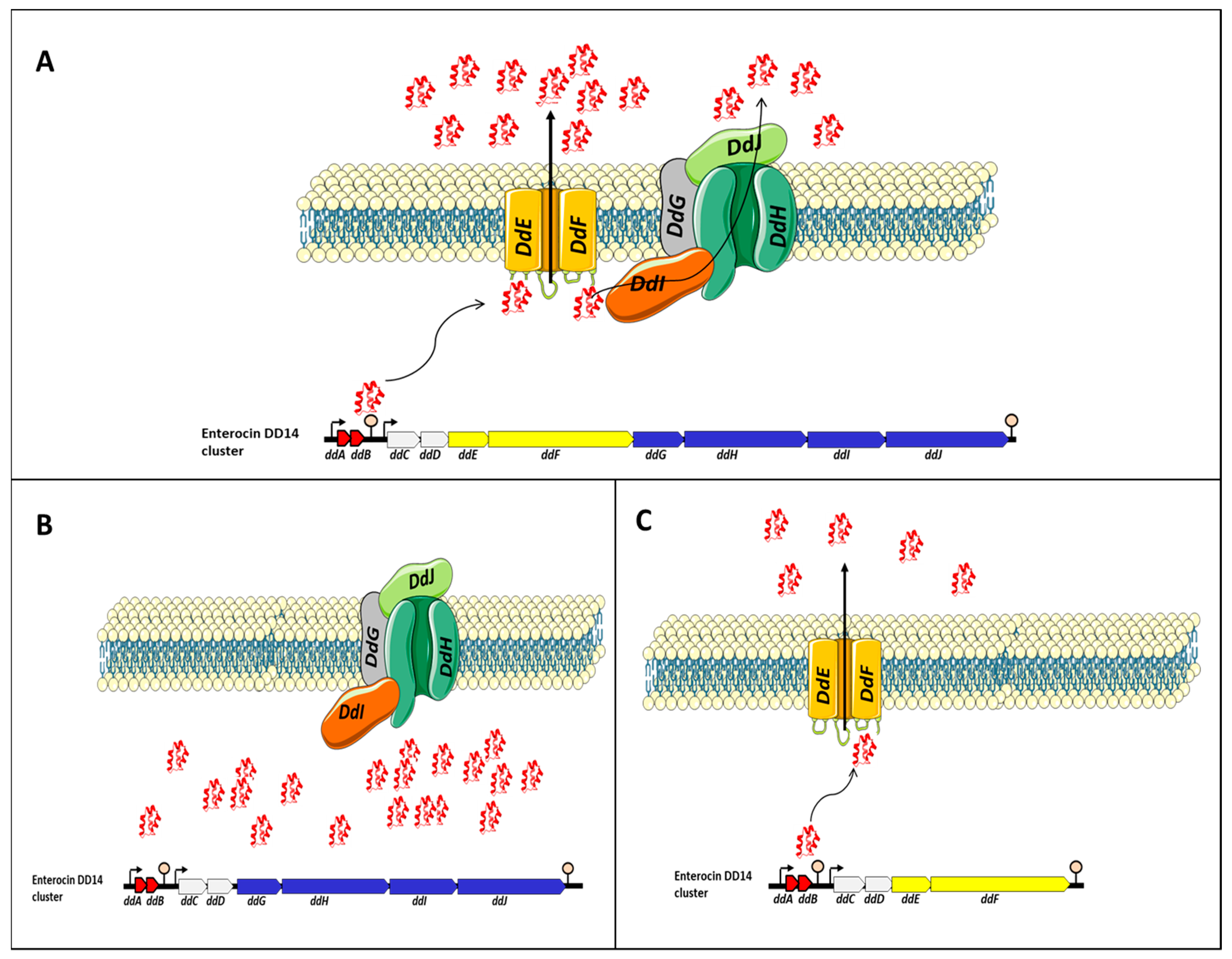
Figure 1. Proposed model for the transport of EntDD14. The legend is presented in the main text. (A) EntDD14 transport in the wild-type strain. (B) In the absence of DdE and/or DdF proteins, the strain was unable to export EntDD14 outside of the cytoplasm and an intracellular accumulation of EntDD14 was observed. (C) In the absence of one or more components the ABC transporter system (DdGHIJ), the bacteriocinogenic strain was still able to produce EntDD14, but at a lower rate than the wild-type.
2. Purification of Enterocin DD14
Despite their renewed interest in many applications, the purification of bacteriocins remains important in many cases, peculiarly in view of obtaining large amounts of purified and relatively clean active fraction which is generally difficult and costly [4][5]. The purification of bacteriocins, including leaderless ones, is usually carried out with a multistep procedure including ammonium sulfate precipitation, followed by one or more chromatography-based methods [4][5]. This strategy was largely utilized to purify bacteriocins devoid of leader peptide [6][7], such as enterocins L50A and L50B [8], A5-11A and A5-11B [9], and 62-6 [10]. Nonetheless, this strategy is time-consuming, expensive, and leads to low yields of purified peptides [5][11]. The ammonium sulfate precipitation enables a reduction in the volume of the culture supernatant and concomitantly increases the specific activity of the bacteriocin [12][13]. This step is usually not suitable for the purification of low molecular weight peptides due to the low precipitation yields and high saturation percentages [14][15][16], and this step requires a dialysis desalting phase, which can trigger significant loss of bacteriocins, because of their hydrophobic nature [16]. To overcome this drawback, easier methods of purification of bacteriocin were developed consisting of two-step-based methods. Such a simplified procedure was successfully applied for the purification of EntDD14 [17]. Accordingly, EntDD14 was purified using ion exchange chromatography and reverse phase HPLC, successively. To sum up, the producing strain, E. faecalis 14, was grown at 37 °C for 24 h with shaking (160 rpm), in an M17 medium supplemented with 5% glucose and buffered with 60 mM sodium phosphate (pH 6.3). After this period of incubation, the culture was centrifuged and the cell-free supernatant was incubated for 24 h at room temperature with CM Sephadex® C-25 resin, which was resuspended and equilibrated in distilled water. The resin was then washed with distilled water and 0.5 M NaCl. Thus, EntDD14 bound to the resin was eluted with 1.5 M NaCl. Removal of the salt from the solution containing EntDD14 was performed by running on PD MidiTrap G-10 columns. This procedure is routinely performed and usually leads to better purification yields. An additional drying step using a speedvac-type rotary concentrator could be applied with the objective to store this peptide at 4 °C until its use [17][18]. To characterize the molecular mass of EntDD14A and EntDD14B, an additional separative step by the HPLC method is required, due to their close masses which are, respectively, 5200.74 Da and 5206.41 Da [17]. These molecular masses are comparable to those reported for L50A and L50B [8], MR10A and MR10B [19], 7A and 7B [20] and A5-11A and A5-11B [9]. It should be, however, mentioned that the purification of bacteriocins could be achieved using other strategies such as membrane separation, which has emerged and applied for the purification of diverse bacteriocins [21][22][23], including some enterocins [24][25].
3. Antibacterial Spectrum of Enterocin DD14 and Potentiation of Antibiotic Activity
Leaderless bacteriocins, mainly enterocins, are active against a wide range of Gram-positive pathogenic bacteria including Listeria monocytogenes, Clostridium perfringens, Enterococcus faecalis and Staphylococcus aureus [8][17][19][26][27]. Related to its spectrum of activity, EntDD14 was proven to be active against MRSA strains in both in vitro [28] and in vivo [29]. The combination of antimicrobials is foreseen to be a valuable tool allowing for the alleviation of bacterial resistance [30]. Related to this, researchers first reported the capability of nisin to potentiate the activity of colistin against Gram-negative bacilli [31]. Next, researchers established the abilities of EntDD28 and EntDD93, two bacteriocins closely related to EntDD14, to augment the activity of erythromycin or kanamycin against the MRSA-S1 strain [27], and that of EntDD14 to potentiate the activity of methicillin against different MRSA strains [28]. Remarkably, MRSA-S1 was resistant to erythromycin and kanamycin; however, if any of these antibiotics was used in combination with EntDD14, EntDD28, or EntDD93, the MRSA-S1 target strain returned to the sensitive phenotype. In addition, an anti-biofilm activity of EntDD28 and EntDD93 was demonstrated on the human MRSA strains [27]. In another study, researchers established that EntDD14 loaded on alginate nanoparticles has a more potent activity across MRSA strains [28], or Clostridium perfringens [18]. It is of note that treatment of C. perfringens with such nanobiotics (bacteriocins loaded on alginate nanoparticles) has annihilated the expression of genes encoding some virulence factors [18]. Recently, a study carried out in the laboratory has established an anti-adhesive action of EntDD14 against several MRSA strains, noticeably across S. aureus USA300, which is known for its virulence traits and ability to colonize different surfaces [32]. Researchers also unveiled the potential of EntDD14 to decrease the secretion of pro-inflammatory interleukins, IL-6 and IL-8, on human intestinal Caco-2 cells previously inflamed with bacterial lipopolysaccharide [32]. Generally active only against closely phylogenetically related microorganism, some LAB-bacteriocins, however, are exceptionally active against Gram-negative bacilli [33][34]. EntDD14 is devoid of such antibacterial activity. Nevertheless, when EntDD14 was used in combination with colistin and tested against a panel of Gram-negative bacilli such as Escherichia coli strains, researchers could observe a better spectrum of colistin, arguing for a potentialization of this antibiotic by EntDD14 [35].
4. The Antiviral Spectrum of Enterocin DD14
Bacteriocins have already been described for their antiviral activity [32][36], and these include EntCRL35 or GEn17 on Herpes Simplex Virus-1 (HSV-1) [37][38]. Moreover, a study based on a global simulation approach established that bacteriocins warrant more attention for their potential against SARS-CoV-2 viruses [39]. Experimental data reported that some bacteriocins such as plantaricin or nisin were able to interact with enveloped viruses and spike protein, respectively [40][41]. It has been observed that EntDD14 has an antiviral activity in vitro against enveloped viruses (personal communication).
5. Cytotoxicity of Enterocin DD14
Bacteriocins such as nisin are used in the food industry as a food preservative and have been extensively studied for their cytotoxic effects on different cell lines. For example, nisin has been shown to have an IC50 of less than 10 µg·mL−1 in intestinal Caco-2 cells after 24 h of contact, and an IC50 of around 150 µg·mL−1 in Vero SF cells (monkey epithelial cells from kidney) [42]. In comparison, enterocin AS-48 (produced by Enterococcus faecalis UGRA 10) showed no cytotoxicity up to 200 µg·mL−1 on melanoma cell line A2058 [43]. In this context, EntDD14 was studied for its cytotoxic effects on several cell lines. First, the cytotoxicity of the purified enterocin DD14 was assessed using the IPEC-1 porcine intestinal epithelial cell line. After 24 h of contact, a dose-dependent decrease in IPEC-1 cell viability was observed when exposed to EntDD14. At concentrations of 50 μg·mL−1 and 100 μg·mL−1, reductions of 9.6% and 20% in cell viability were noted, respectively [44]. The cytotoxic effect of EntDD14 on two cell lines of human origin, intestinal Caco-2 cells, and HT29 mucus-producing cells was studied [18]. Thus, no apparent cytotoxicity was observed when EntDD14 was in contact with HT29 cells for 4 h and 24 h incubation and the two concentrations were tested (60 and 120 µg·mL−1). Regarding Caco-2 cells, the viability remained approximately 95% when exposed to a concentration of 120 µg·mL−1 of EntDD14 for 24 h [18]. Finally, another study (data under publication elsewhere) has shown the cytotoxic effects of EntDD14 on two cell lines, VERO E6 cells (derived from the kidney of an African green monkey) and Huh 7 cells (human hepatocellular carcinoma). For both cell lines, the viability decreased to 75% after 24 h incubation with EntDD14 at 120 µg·mL−1 (Submitted). To conclude, EntDD14 was found to be less cytotoxic than nisin.
References
- Ladjouzi, R.; Lucau-Danila, A.; Benachour, A.; Drider, D. A Leaderless Two-Peptide Bacteriocin, Enterocin DD14, Is Involved in Its Own Self-Immunity: Evidence and Insights. Front. Bioeng. Biotechnol. 2020, 8, 644.
- Pérez-Ramos, A.; Ladjouzi, R.; Mihasan, M.; Teiar, R.; Benachour, A.; Drider, D. Advances in Characterizing the Transport Systems of and Resistance to EntDD14, A Leaderless Two-Peptide Bacteriocin with Potent Inhibitory Activity. Int. J. Mol. Sci. 2023, 24, 1517.
- Pérez-Ramos, A.; Ladjouzi, R.; Benachour, A.; Drider, D. Evidence for the Involvement of Pleckstrin Homology Domain-Containing Proteins in the Transport of Enterocin DD14 (EntDD14); a Leaderless Two-Peptide Bacteriocin. Int. J. Mol. Sci. 2021, 22, 12877.
- De Vuyst, L.; Leroy, F. Bacteriocins from Lactic Acid Bacteria: Production, Purification, and Food Applications. J. Mol. Microbiol. Biotechnol. 2007, 13, 194–199.
- Jamaluddin, N.; Stuckey, D.C.; Ariff, A.B.; Faizal Wong, F.W. Novel Approaches to Purifying Bacteriocin: A Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 2453–2465.
- Ovchinnikov, K.V.; Chi, H.; Mehmeti, I.; Holo, H.; Nes, I.F.; Diep, D.B. Novel Group of Leaderless Multipeptide Bacteriocins from Gram-Positive Bacteria. Appl. Environ. Microbiol. 2016, 82, 5216–5224.
- Carlin Fagundes, P.; Nascimento de Sousa Santos, I.; Silva Francisco, M.; Mattos Albano, R.; de Freire Bastos, M.D.C. Genetic and Biochemical Characterization of Hyicin 3682, the First Bacteriocin Reported for Staphylococcus hyicus. Microbiol. Res. 2017, 198, 36–46.
- Cintas, L.M.; Casaus, P.; Herranz, C.; Hâvarstein, L.S.; Holo, H.; Hernández, P.E.; Nes, I.F. Biochemical and Genetic Evidence That Enterococcus faecium L50 Produces Enterocins L50A and L50B, the Sec-Dependent Enterocin P, and a Novel Bacteriocin Secreted without an N-Terminal Extension Termed Enterocin Q. J. Bacteriol. 2000, 182, 6806–6814.
- Batdorj, B.; Dalgalarrondo, M.; Choiset, Y.; Pedroche, J.; Métro, F.; Prévost, H.; Chobert, J.-M.; Haertlé, T. Purification and Characterization of Two Bacteriocins Produced by Lactic Acid Bacteria Isolated from Mongolian airag. J. Appl. Microbiol. 2006, 101, 837–848.
- Dezwaan, D.C.; Mequio, M.J.; Littell, J.S.; Allen, J.P.; Rossbach, S.; Pybus, V. Purification and Characterization of Enterocin 62-6, a Two-Peptide Bacteriocin Produced by a Vaginal Strain of Enterococcus faecium: Potential Significance in Bacterial Vaginosis. Microb. Ecol. Health Dis. 2007, 19, 241–250.
- Ahmad, V.; Jamal, Q.M.S.; Siddiqui, M.U.; Shukla, A.K.; Alzohairy, M.A.; Al Karaawi, M.A.; Kamal, M.A. Methods of Screening-Purification and Antimicrobial Potentialities of Bacteriocin in Health Care. Curr. Drug Metab. 2017, 18, 814–830.
- Pingitore, E.V.; Salvucci, E.; Sesma, F.; Nader-Macias, M.E. Different Strategies for Purification of Antimicrobial Peptides from Lactic Acid Bacteria (LAB). Commun. Curr. Res. Educ. Top. Trends Appl. Microbiol. 2007, 1, 557–568.
- Cui, Y.; Luo, L.; Wang, X.; Lu, Y.; Yi, Y.; Shan, Y.; Liu, B.; Zhou, Y.; Lü, X. Mining, Heterologous Expression, Purification, Antibactericidal Mechanism, and Application of Bacteriocins: A Review. Compr. Rev. Food Sci. Food Saf. 2021, 20, 863–899.
- Guyonnet, D.; Fremaux, C.; Cenatiempo, Y.; Berjeaud, J.M. Method for Rapid Purification of Class IIa Bacteriocins and Comparison of Their Activities. Appl. Environ. Microbiol. 2000, 66, 1744–1748.
- Borzenkov, V.; Surovtsev, V.; Dyatlov, I. Obtaining Bacteriocins by Chromatographic Methods. Adv. Biosci. Biotechnol. 2014, 2014, 55054.
- Yap, P.G.; Lai, Z.W.; Tan, J.S. Bacteriocins from Lactic Acid Bacteria: Purification Strategies and Applications in Food and Medical Industries: A Review. Beni-Suef Univ. J. Basic Appl. Sci. 2022, 11, 51.
- Caly, D.L.; Chevalier, M.; Flahaut, C.; Cudennec, B.; Al Atya, A.K.; Chataigné, G.; D’Inca, R.; Auclair, E.; Drider, D. The Safe Enterocin DD14 Is a Leaderless Two-Peptide Bacteriocin with Anti-Clostridium Perfringens Activity. Int. J. Antimicrob. Agents 2017, 49, 282–289.
- Zgheib, H.; Belguesmia, Y.; Boukherroub, R.; Drider, D. Alginate Nanoparticles Enhance Anti-Clostridium Perfringens Activity of the Leaderless Two-Peptide Enterocin DD14 and Affect Expression of Some Virulence Factors. Probiotics Antimicrob. Proteins 2021, 13, 1213–1227.
- Martín-Platero, A.M.; Valdivia, E.; Ruíz-Rodríguez, M.; Soler, J.J.; Martín-Vivaldi, M.; Maqueda, M.; Martínez-Bueno, M. Characterization of Antimicrobial Substances Produced by Enterococcus faecalis MRR 10-3, Isolated from the Uropygial Gland of the Hoopoe (Upupa epops). Appl. Environ. Microbiol. 2006, 72, 4245–4249.
- Liu, X.; Vederas, J.C.; Whittal, R.M.; Zheng, J.; Stiles, M.E.; Carlson, D.; Franz, C.M.A.P.; McMullen, L.M.; van Belkum, M.J. Identification of an N-Terminal Formylated, Two-Peptide Bacteriocin from Enterococcus faecalis 710C. J. Agric. Food Chem. 2011, 59, 5602–5608.
- Ugras, S.; Sezen, K.; Kati, H.; Demirbag, Z. Purification and Characterization of the Bacteriocin Thuricin Bn1 Produced by Bacillus thuringiensis subsp. kurstaki Bn1 Isolated from a Hazelnut Pest. J. Microbiol. Biotechnol. 2013, 23, 167–176.
- Berić, T.; Stanković, S.; Draganić, V.; Kojić, M.; Lozo, J.; Fira, D. Novel Antilisterial Bacteriocin Licheniocin 50.2 from Bacillus licheniformis VPS50.2 Isolated from Soil Sample. J. Appl. Microbiol. 2014, 116, 502–510.
- Forestier, A.; Belguesmia, Y.; Krier, F.; Drider, D.; Dhulster, P.; Firdaous, L. Recovery of Nisin from Culture Supernatants of Lactococcus lactis by Ultrafiltration: Flux Properties and Separation Efficiency. Food Bioprod. Process. 2022, 136, 196–210.
- Izquierdo, E.; Wagner, C.; Marchioni, E.; Aoude-Werner, D.; Ennahar, S. Enterocin 96, a Novel Class II Bacteriocin Produced by Enterococcus faecalis WHE 96, Isolated from Munster Cheese. Appl. Environ. Microbiol. 2009, 75, 4273–4276.
- Khan, H.; Flint, S.H.; Yu, P.-L. Development of a Chemically Defined Medium for the Production of Enterolysin A from Enterococcus faecalis B9510. J. Appl. Microbiol. 2013, 114, 1092–1102.
- Acedo, J.Z.; van Belkum, M.J.; Lohans, C.T.; Towle, K.M.; Miskolzie, M.; Vederas, J.C. Nuclear Magnetic Resonance Solution Structures of Lacticin Q and Aureocin A53 Reveal a Structural Motif Conserved among Leaderless Bacteriocins with Broad-Spectrum Activity. Biochemistry 2016, 55, 733–742.
- Al Atya, A.K.; Belguesmia, Y.; Chataigne, G.; Ravallec, R.; Vachée, A.; Szunerits, S.; Boukherroub, R.; Drider, D. Anti-MRSA Activities of Enterocins DD28 and DD93 and Evidences on Their Role in the Inhibition of Biofilm Formation. Front. Microbiol. 2016, 7, 817.
- Belguesmia, Y.; Spano, G.; Drider, D. Potentiating Effects of Leaderless Enterocin DD14 in Combination with Methicillin on Clinical Methicillin-Resistant Staphylococcus aureus S1 Strain. Microbiol. Res. 2021, 252, 126864.
- Bendjeddou, K.; Hamma-Faradji, S.; Meddour, A.A.; Belguesmia, Y.; Cudennec, B.; Bendali, F.; Daube, G.; Taminiau, B.; Drider, D. Gut Microbiota, Body Weight and Histopathological Examinations in Experimental Infection by Methicillin-Resistant Staphylococcus aureus: Antibiotic versus Bacteriocin. Benef. Microbes 2021, 12, 295–305.
- Zgheib, H.; Drider, D.; Belguesmia, Y. Broadening and Enhancing Bacteriocins Activities by Association with Bioactive Substances. Int. J. Environ. Res. Public Health 2020, 17, 7835.
- Naghmouchi, K.; Baah, J.; Hober, D.; Jouy, E.; Rubrecht, C.; Sané, F.; Drider, D. Synergistic Effect between Colistin and Bacteriocins in Controlling Gram-Negative Pathogens and Their Potential to Reduce Antibiotic Toxicity in Mammalian Epithelial Cells. Antimicrob. Agents Chemother. 2013, 57, 2719–2725.
- Teiar, R.; Pérez-Ramos, A.; Zgheib, H.; Cudennec, B.; Belguesmia, Y.; Drider, D. Anti-Adhesion and Anti-Inflammatory Potential of the Leaderless Class IIb Bacteriocin Enterocin DD14. Probiotics Antimicrob. Proteins 2022, 14, 613–619.
- Messaoudi, S.; Kergourlay, G.; Dalgalarrondo, M.; Choiset, Y.; Ferchichi, M.; Prévost, H.; Pilet, M.F.; Chobert, J.M.; Manai, M.; Dousset, X. Purification and Characterization of a New Bacteriocin Active against Campylobacter Produced by Lactobacillus salivarius SMXD51. Food Microbiol. 2012, 32, 129–134.
- Line, J.E.; Svetoch, E.A.; Eruslanov, B.V.; Perelygin, V.V.; Mitsevich, E.V.; Mitsevich, I.P.; Levchuk, V.P.; Svetoch, O.E.; Seal, B.S.; Siragusa, G.R.; et al. Isolation and Purification of Enterocin E-760 with Broad Antimicrobial Activity against Gram-Positive and Gram-Negative Bacteria. Antimicrob. Agents Chemother. 2008, 52, 1094–1100.
- Al Atya, A.K.; Abriouel, H.; Kempf, I.; Jouy, E.; Auclair, E.; Vachée, A.; Drider, D. Effects of Colistin and Bacteriocins Combinations on the In Vitro Growth of Escherichia coli Strains from Swine Origin. Probiotics Antimicrob. Proteins 2016, 8, 183–190.
- Ramakrishnan, M.A. Determination of 50% Endpoint Titer Using a Simple Formula. World J. Virol. 2016, 5, 85.
- Wachsman, M.B.; Castilla, V.; de Ruiz Holgado, A.P.; de Torres, R.A.; Sesma, F.; Coto, C.E. Enterocin CRL35 Inhibits Late Stages of HSV-1 and HSV-2 Replication in Vitro. Antivir. Res. 2003, 58, 17–24.
- Cavicchioli, V.Q.; Carvalho, O.V.; Paiva, J.C.; Todorov, S.D.; Silva Júnior, A.; Nero, L.A. Inhibition of Herpes Simplex Virus 1 (HSV-1) and Poliovirus (PV-1) by Bacteriocins from Lactococcus lactis subsp. lactis and Enterococcus Durans Strains Isolated from Goat Milk. Int. J. Antimicrob. Agents 2018, 51, 33–37.
- Erol, I.; Kotil, S.E.; Ortakci, F.; Durdagi, S. Exploring the Binding Capacity of Lactic Acid Bacteria Derived Bacteriocins against RBD of SARS-CoV-2 Omicron Variant by Molecular Simulations. J. Biomol. Struct. Dyn. 2023, 1–11.
- Omer, A.A.M.; Hinkula, J.; Tran, P.-T.-H.; Melik, W.; Zattarin, E.; Aili, D.; Selegård, R.; Bengtsson, T.; Khalaf, H. Plantaricin NC8 Aβ Rapidly and Efficiently Inhibits Flaviviruses and SARS-CoV-2 by Disrupting Their Envelopes. PLoS ONE 2022, 17, e0278419.
- Bhattacharya, R.; Gupta, A.M.; Mitra, S.; Mandal, S.; Biswas, S.R. A Natural Food Preservative Peptide Nisin Can Interact with the SARS-CoV-2 Spike Protein Receptor Human ACE2. Virology 2021, 552, 107–111.
- Kourmentza, K.; Gromada, X.; Michael, N.; Degraeve, C.; Vanier, G.; Ravallec, R.; Coutte, F.; Karatzas, K.A.; Jauregi, P. Antimicrobial Activity of Lipopeptide Biosurfactants against Foodborne Pathogen and Food Spoilage Microorganisms and Their Cytotoxicity. Front. Microbiol. 2021, 11, 561060.
- Abengózar, M.Á.; Cebrián, R.; Saugar, J.M.; Gárate, T.; Valdivia, E.; Martínez-Bueno, M.; Maqueda, M.; Rivas, L. Enterocin AS-48 as Evidence for the Use of Bacteriocins as New Leishmanicidal Agents. Antimicrob. Agents Chemother. 2017, 61, e02288-16.
- Lynch, D.; O’Connor, P.M.; Cotter, P.D.; Hill, C.; Field, D.; Begley, M. Identification and Characterisation of Capidermicin, a Novel Bacteriocin Produced by Staphylococcus capitis. PLoS ONE 2019, 14, e0223541.
More
Information
Subjects:
Microbiology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
751
Revisions:
2 times
(View History)
Update Date:
25 Jul 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No