Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Xiaoman Zhou | -- | 2646 | 2023-06-20 03:08:02 | | | |
| 2 | Conner Chen | -10 word(s) | 2636 | 2023-06-25 02:07:48 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Zhou, X.; Chi, K.; Zhang, C.; Liu, Q.; Yang, G. Sialylation Accumulates in Cancer Tissues and Promotes Development. Encyclopedia. Available online: https://encyclopedia.pub/entry/45821 (accessed on 26 June 2026).
Zhou X, Chi K, Zhang C, Liu Q, Yang G. Sialylation Accumulates in Cancer Tissues and Promotes Development. Encyclopedia. Available at: https://encyclopedia.pub/entry/45821. Accessed June 26, 2026.
Zhou, Xiaoman, Kaijun Chi, Chairui Zhang, Quan Liu, Ganglong Yang. "Sialylation Accumulates in Cancer Tissues and Promotes Development" Encyclopedia, https://encyclopedia.pub/entry/45821 (accessed June 26, 2026).
Zhou, X., Chi, K., Zhang, C., Liu, Q., & Yang, G. (2023, June 20). Sialylation Accumulates in Cancer Tissues and Promotes Development. In Encyclopedia. https://encyclopedia.pub/entry/45821
Zhou, Xiaoman, et al. "Sialylation Accumulates in Cancer Tissues and Promotes Development." Encyclopedia. Web. 20 June, 2023.
Copy Citation
The tumor microenvironment (TME), where the tumor cells incite the surrounding normal cells to create an immune suppressive environment, reduces the effectiveness of immune responses during cancer development. Sialylation, a type of glycosylation that occurs on cell surface proteins, lipids, and glycoRNAs, is known to accumulate in tumors and acts as a “cloak” to help tumor cells evade immunological surveillance. The role of sialylation in tumor proliferation and metastasis has become increasingly evident. With the advent of single-cell and spatial sequencing technologies, more research is being conducted to understand the effects of sialylation on immunity regulation.
sialylation
Siglecs
immune checkpoint
1. Typical Sialylated Glycans in Tumors
Cancer cells are cloaked by a plethora of glycosylation, with many of them exhibiting a high level of sialylation. Glycosylation has the potential to mask essential antigenic and receptor-binding sites, as well as interact with certain checkpoints, enabling cells to avoid being recognized by enemy-identifying signals [1][2]. In addition to physical shielding, certain glycan structures possess specific physiological functions; they can act as ligands to mediate signal recognition and immune suppression [3]. These glycans comprise sialyl-Tn (STn), sialyl-T (ST), disialyl-T, sialyl-Lewis antigens, polysialic acid, and gangliosides (Figure 1). The broad distribution and advantageous effects on cancer cells have rendered sialylated glycans a hallmark of cancer [4].
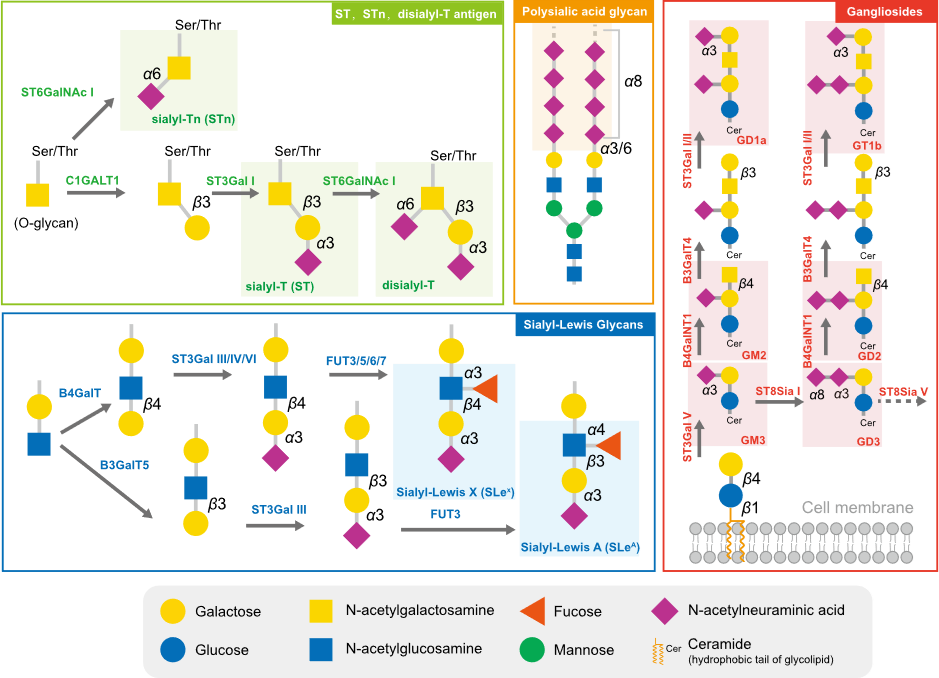
Figure 1. The typical sialylated glycans in tumors. The sialylated glycan structures typically highly expressed in tumors are marked with the translucent rectangular box.
The short sialylated O-glycan STn and ST antigens are aberrantly expressed in several cancers, with high levels being mainly observed in carcinomas and associated with aggressive tumors, such as those with chemotherapy resistance and poor prognosis [5][6]. The increased level of STn in tumor cells is primarily attributed to the upregulation of ppGalNAcT and ST6GalNAc I, combined with the low efficiency of COSMC, which assists C1GALT1 in extending glycans [6]. High serum concentrations of STn have been observed in patients with breast cancer [5], prostate cancer [6], bladder cancer [7], cervical cancer [8], and ovarian cancers and correlated to tumor grade and metastasis [9]. Due to the high prevalence of STn antigen expression in tumors, serum STn levels can be clinically utilized to assist with tumor diagnosis. ST antigens were also elevated in cancer, and the silencing of ST3GAL1 significantly reduced the level of these antigens, further reducing the tumor size in the prostate cancer xenograft mouse model [10].
Sialyl-Lewis (SLe) antigens, SLeA and SLeX, are other typical structures of sialyl glycans and act as ligands for selectins, a family of lectins involved in lymphocyte trafficking. Cancer cells that are disguised by SLeA and SLeX are mistaken for leucocytes during their epithelial–mesenchymal transition and travel throughout the circulation system [11]. E-selectins, the primary receptors of SLeA and SLeX, are adhesion molecules required for leukocyte recruitment during the early stages of inflammation. The accumulation of SLeA and SLeX leads to cell adhesion and subsequent trans endothelial migration of traveling tumor cells. In ovarian tumors, mucins MUC16 and MUC1 are major carriers of SLeA and SLex and are used as biomarkers [12].
Polysialic acid is a polymer of sialic acid, with α2,8 and α2,9 linkages and a length ranging from 8 to 400 units. It is a crucial glycosylation type for several essential proteins, providing them with a negative charge. In mammalian cells, three sialyltransferases (ST8Sia II, ST8Sia III, and ST8Sia IV) are responsible for the extension of polysialic acid glycans [13]. Polysialic acid is often detected in brain tissues and is also found in immune cells. Several molecules are found to undergo polysialylation, such as neural cell adhesion molecules (NCAMs) [14][15], chemokine receptor 7 (CCR7) [16][17], CD36 [18], and E-selectin ligand 1 (ESL-1) [19]. Interestingly, the sialyltransferase ST8Sia II and ST8Sia III can polysialylate independently. These polysialylated glycoproteins participate in physiological processes, including cell adhesion between cells and cells with a matrix, cell migration, synapse formation, and the functional plasticity of the developing nervous system. In tumor cells, the level of polysialic acid chains correlates with an aggressive phenotype and the resistance of cancer treatment [20].
Glycolipids, molecules composed of one or more carbohydrate residues linked to a hydrophobic lipid moiety via a β-glycosidic linkage, are mainly found in lipid rafts on the outer leaflet of the plasma membrane bilayer. Some gangliosides (GD2, GD3, GM2, GM3, fucosyl-GM1) are distinguishable from other gangliosides via their significantly lower or absent expression in normal cells, yet they are highly expressed in tumor cells [21]. GD3 and GD2 are highly expressed in certain tumors and can be utilized as targets for immunotherapy. However, they can also inhibit the function of immune cells, such as macrophages, via binding to Siglec-7 [22].
2. Sialyltransferases Are Critical Enzymes for Hypersialylation
Approximately twenty sialyltransferases are involved in the positive regulation of human cell sialylation, and these sialyltransferases are classified into four types based on the differences in their substrate structure and the linkage of the formed sialylation: ST3Gal I-VI (add Neu5Ac to galactose in an α2,3 linkage); ST6Gal I-II (add Neu5Ac to galactose in an α2,6 linkage); ST6GalNAc I-VI (add Neu5Ac to GalNAc in an α2,6 linkage); and ST8Sia I-VI (add Neu5Ac to Neu5Ac in an α2,8 or α2,9 linkage) [23]. The increased modification of sialic acid in various malignant tumors is caused by the high expression of sialyltransferases. Hypersialylation in the tumor microenvironment alters its physiological characteristics, blocking some immunological recognition and communication [24]. More importantly, sialylation also promotes tumor cell survival and drug resistance, as well as suppressing surrounding immune cells, which helps tumor cells to survive [25].
The sialyltransferase ST6Gal I is a well-studied enzyme, catalyzing the addition of Neu5Ac to galactose residues of Gal β1-4 GlcNAc in an α2,6 bond mainly on N-glycans. Elevated levels of ST6Gal I have been linked to a number of different cancer types and can be a driver of malignant progression, as well as resistance to therapy [26][27][28][29][30][31][32]. This is further supported by the pancreas-specific genetic deletion of ST6GAL1 in a mouse model, which delays cancer formation [33]. Additionally, ST6Gal I has been observed to add sialylation in cell surface receptors, such as PDGFRB [27], EGFR [34][35], and PECAM [36], which increases protein levels and phosphorylation to stimulate pathways such as PI3K/AKT and RAS [26], thus contributing to tumor growth. ST6Gal I has also been implicated in the immune evasion in hepatocarcinoma cells, where it increases levels of MMP9 and suppresses T-cell proliferation [37]. Moreover, ST6Gal I is released into the extracellular milieu in either exosome or free forms, thereby remodeling cell surface and secreted glycans, which has been linked to aggressive tumor cell proliferation in breast cancer [38]. ST6Gal II is another enzyme that can add α2,6-linked sialic acid to N-glycans. Nonetheless, this enzyme is predominately expressed in the embryonic and perinatal stages of brain tissue [39]. In a recent study of breast cancer, ST6Gal II accumulated in tumor tissue and was associated with tumor malignancy. The inhibition of ST6Gal II caused the downregulation of cell adhesion and invasion-associated proteins, resulting in reduced tumor migration [40]. Similarly, silencing ST6GAL2 in a follicular thyroid carcinoma reduced tumor growth by inactivating the Hippo pathway in an in vivo model [41].
The ST3Gal family, consisting of six members (ST3Gal I-VI), facilitates the transfer of sialic acid to the terminal galactose residues of glycochains via an α2,3-linkage in both glycoproteins and glycolipids. ST3Gal I, which predominantly functions in core-1 O-glycans, catalyzes the transfer of Neu5Ac to a galactose residue in an α2,3 bond to produce sialyl-T antigen. The upregulation of ST3Gal I has been reported in many malignant tissues, such as ovarian cancer [42], glioblastoma tumors [43], and melanomas [44], and it has been associated with tumorigenesis, poor clinical outcomes and an inflammatory phenotype. Additionally, CD55, an essential immune checkpoint molecule, has been reported to be O-glycosylated by ST3Gal I to help cancer cells escape immune attack [45].
ST3Gal II, in contrast, prefers gangliosides as its substrate to form GD1a and GT1b [46][47]. The elevated expression of ST3Gal II has been associated with advanced stages of cancer and poor clinical outcomes. In addition, ST3Gal II is the only enzyme responsible for synthesizing the glycosphingolipid SSEA4, a well-known biomarker of several cancers [48][49]. Furthermore, ST3GAL2 knockdown led to a dramatic growth reduction in colorectal cancer in xenografted mice models [50].
ST3Gal III, ST3Gal IV, and ST3Gal VI are implicated in the formation of SLeA and SLeX glycans on the cell surface, which act as binding ligands for selectins and are essential for metastasis [51][52]. The high expression of ST3Gal III has a strong positive correlation with poor prognosis in gastric cancer [53], and ST3Gal IV is the main enzyme for generating ligands of Siglec-7 and -9 [54][55]. ST3Gal VI generates selectin ligands and accumulates in liver and urinary bladder cancers [56][57]. Moreover, ST3GAL5 encodes GM3 synthase, the rate-limiting enzyme for the production of downstream gangliosides, and is, therefore, crucial to gangliosides synthesis [46]. In renal cell carcinoma research, ST3GAL5 was consistently overexpressed in tumor tissue and correlated with the infiltration of exhausted CD8+ T cells, indicating that ST3Gal V contributes to immune suppression [58].
The ST6GalNAc family, consisting of six members, ST6GalNAc I-VI, catalyzes the α2,6 glycosidic linkage of Neu5Ac to the GalNAc residues on O-glycans or glycolipids. ST6GalNAc I, which adds Neu5Ac to O-linked GalNAc residues to form sialyl-Tn (STn), is particularly significant [59]. Evidence suggests that the overexpression of STn is associated with poor clinical prognosis in a wide range of cancer types [9][60], making it a well-known tumor-associated carbohydrate antigen. One functional study found that ST6GalNAc I can promote tumor growth and metastasis and is related to cancer cell stemness [61]. Furthermore, cytokines such as IL-13 promote the phosphorylation of STAT6, which, in turn, activates the transcription of ST6GALNAC1, thereby facilitating the formation of STn [62]. The STn inhibits T-cell responses by binding to Siglec-15, leading to immune evasion in the tumor microenvironment [63][64].
ST6GalNAc II is an enzyme responsible for synthesizing ST and STn antigens. The role of ST6GalNAc II in tumors varies with the stage and status of the tumor. In breast cancer metastasis, ST6GalNAc II catalyzes the formation of ST and STn that blocks tumor binding to galectin, negatively affecting tumor metastasis [65][66]. However, in the tumor microenvironment, ST6GalNAc II is positively correlated with higher tumor stage and worse prognosis [67].
Another ST6GalNAc family member for O-glycan is ST6GalNAc IV, a key ST6GalNAc enzyme that is involved in the formation of disialyl-T antigen and GD1α from sialyl-lactotetraosyl-ceramide GM1b (gangliosides). In a primary lung cancer model, the upregulation of ST6GALNAC4 was demonstrated to confer glycosylation changes in tumor cells, contributing to their metastatic activity. This is likely due to the preservation of the T-antigen presentation and adherence to galectin 3 [68]. The catalytic product of ST6GalNAc IV, the disialyl-T antigen, was shown to be a ligand for Siglec-7. The high expression of ST6GalNAc IV increased disialyl-T antigens in CD162 and CD45 and inhibited NK cell activity via the binding of Siglec-7 in chronic lymphocytic leukemia B cells [69]. Moreover, in liver cancer, the elevated ST6GALNAC4 promoted tumor proliferation, migration and invasion ability, and affected the expression of immune checkpoints on tumor cells [70].
The sialyltransferases ST6GalNAc III, ST6GalNAc V, and ST6GalNAc VI are mainly involved in glycolipid synthesis. ST6GalNAc III and ST6GalNAc V use GM1b as a substrate to synthesize GD1α [71][72], whereas ST6GalNAc VI catalyzes the synthesis of α-series gangliosides, including GD1α, GT1aα, and GQ1bα; globo-series glycosphingolipids (GSL); and disialyl LeA. It has been reported that ST6GalNAc III increased M2 macrophages via the accumulation of prostaglandin and arachidonic acid in gastric cancer [73]. ST6GALNAC5 was expressed at low levels in tumors, and its overexpression significantly inhibited tumor growth and invasiveness [74]. On the other hand, the downregulation of ST6GALNAC6 resulted in a change from disialyl LeA to sialyl LeA and an elevation in E-selectin binding activity during metastasis, which supports inflammation-driven carcinogenesis by reducing its binding to the immunoregulatory Siglec-7 [75].
Members of the ST8Sia family catalyze the transfer of sialic acid to another sialic acid, forming α2,8 linkages. Notably, the 2,8-disialic glycan structure, ligands for Siglec-7 and Siglec-9, can potentially regulate immune responses. Notably, ST8Sia I, also known as GD3 synthase, is positively correlated with the astrocytoma grade and accumulates in glioblastomas [76]. Similarly, ST8Sia II and ST8Sia IV are polysialyltransferases that produce polysialylated cell adhesion molecules, which are highly expressed during cancer development. In tumors, the expression of ST8SIA2 has been shown to correlate with the tumor stage [77]. Moreover, the overexpression of ST8SIA2 increased the invasiveness and metastatic abilities of small-cell lung cancer cells in vitro [77][78]. Additionally, ST8SIA4 is overexpressed in breast cancer tissues and contributes to chemoresistance in acute myeloid leukemia [79][80]. Furthermore, ST8Sia III, which causes the sialylation of a variety of glycolipids (GM3, GD3, and α2,3-sialylparagloboside), was identified as a therapeutic target for glioblastomas [81]. Other ST8Sia family members, such as ST8Sia V and ST8Sia VI, are also related to malignant potential. ST8Sia V, the enzyme adding Neu5Ac to gangliosides, was expressed at a low level and negatively correlated with patient survival in bladder cancer and colon cancer [82][83]. ST8Sia VI generates disialic acid structures preferentially on O-linked glycoproteins, and these products are proven to bind with Siglec-7 and Siglec-9. Studies have shown that ST8Sia VI contributes to tumor growth in a mouse model by inhibiting immune responses via the alteration of the macrophage polarization towards M2 and increasing the immune modulator arginase in the tumor microenvironment [84].
3. The Function of Sialidases in Tumor Sialylation Regulation
The sialidases and sialyltransferases in cells collectively act to maintain sialylation homeostasis. In tumor cells, an abnormally increased level of sialylation is generally attributed to elevated sialyltransferase activities; however, the role of sialidases in regulating the sialylation levels remains to be addressed.
Mammalian sialidases, NEU1-4, are enzymes with distinct cellular localizations. NEU1, mainly present in lysosomes, is associated with the degradation of sialylated glycans and the recycling of sialic acid. Evidence suggests that the upregulation of NEU1 in cancers may increase the utilization of sialic acid, thus contributing to the maintenance of cell sialylation. NEU1 is highly expressed in various cancers, such as liver cancer [85], pancreatic cancer [86], ovarian cancer [87], and melanoma [88]. However, NEU1 is reported to be expressed at low levels in certain stages of tumors and has been found to remove cytosolic sialic acid modifications and inhibit tumor progression [89][90]. Therefore, the effect of NEU1 on tumors must be comprehensively and dialectically analyzed. NEU2 is located in the cytosol and predominantly inhibits tumor growth. The decrease in NEU2 leads to increased sialylation levels and reduces the stemness-like properties of cancer stem cells [91]. Additionally, NEU2 causes a reduction in α2,6-linked sialylation on the Fas protein, leading to apoptosis in pancreatic cancer [92]. In ovarian cancer cells, the overexpression of NEU2 leads to a significant reduction in α2,3- and α2,6-linked sialylation and induces cellular autophagy by upregulating the expression of ATG5, an essential protein involved in autophagosome formation [93]. NEU3, a membrane sialidase, is essential for the hydrolysis of sialic acid in ganglioside. In colon cancer, the upregulation of NEU3 accumulates lactosylceramide and leads to protection against programmed cell death [94]. NEU4 is located in the ER membrane, mitochondria and lysosomes, and is downregulated in many tumors. NEU4 has been reported to negatively regulate the motility of tumors via the desialylation of CD44 in hepatocellular carcinoma [95], as well as reduce sialyl Lewis antigens to prevent cell adhesion to E-selectin in colon cancer [96]. While the role of sialidases in tumors may vary based on tumor type and status, tumor cells consistently regulate sialidase expression and control sialylation in a way that promotes tumor progression.
Sialyltransferases and sialidases are strictly and dynamically regulated to increase and maintain high sialylation levels, which helps to induce the immunosuppressive status of the tumor microenvironment via interactions with immune cells, thus facilitating tumor survival and growth.
References
- Zheng, L.; Yang, Q.; Li, F.; Zhu, M.; Yang, H.; Tan, T.; Wu, B.; Liu, M.; Xu, C.; Yin, J.; et al. The Glycosylation of Immune Checkpoints and Their Applications in Oncology. Pharmaceuticals 2022, 15, 1451.
- Casalino, L.; Gaieb, Z.; Goldsmith, J.A.; Hjorth, C.K.; Dommer, A.C.; Harbison, A.M.; Fogarty, C.A.; Barros, E.P.; Taylor, B.C.; McLellan, J.S.; et al. Beyond Shielding: The Roles of Glycans in the SARS-CoV-2 Spike Protein. ACS Cent. Sci. 2020, 6, 1722–1734.
- Gonzalez-Gil, A.; Schnaar, R.L. Siglec Ligands. Cells 2021, 10, 1260.
- Vajaria, B.N.; Patel, P.S. Glycosylation: A hallmark of cancer? Glycoconj. J. 2017, 34, 147–156.
- Xu, F.; Gao, Y.; Diao, X.; Li, J.; Jiang, H.; Zhao, H. Diagnostic value of sialyl-Tn immunocytochemistry in breast cancer presenting with pathological nipple discharge. Cancer Med. 2021, 10, 1783–1790.
- Yamamoto, D.; Hongo, H.; Kosaka, T.; Aoki, N.; Oya, M.; Sato, T. The sialyl-Tn antigen synthase genes regulates migration-proliferation dichotomy in prostate cancer cells under hypoxia. Glycoconj. J. 2023, 40, 199–212.
- Carvalho, S.; Abreu, C.M.; Ferreira, D.; Lima, L.; Ferreira, J.A.; Santos, L.L.; Ribeiro, R.; Grenha, V.; Martinez-Fernandez, M.; Duenas, M.; et al. Phenotypic Analysis of Urothelial Exfoliated Cells in Bladder Cancer via Microfluidic Immunoassays: Sialyl-Tn as a Novel Biomarker in Liquid Biopsies. Front. Oncol. 2020, 10, 1774.
- Benbrook, D.M.; Deng, W.; Gold, M.A.; Rai, R.; Conrad, R.; van der Wel, H.; Husain, S.; Moore, K.; Spirtos, N.; Jackson, A.L.; et al. Association of Sialyl Tn antigen with cervical cancer lymph node status: An NRG oncology/GOG study. Gynecol. Oncol. 2023, 171, 67–75.
- Munkley, J. The Role of Sialyl-Tn in Cancer. Int. J. Mol. Sci. 2016, 17, 275.
- Bai, R.; Luan, X.; Zhang, Y.; Robbe-Masselot, C.; Brockhausen, I.; Gao, Y. The expression and functional analysis of the sialyl-T antigen in prostate cancer. Glycoconj. J. 2020, 37, 423–433.
- Manfioletti, G.; Fedele, M. Epithelial-Mesenchymal Transition (EMT) 2021. Int. J. Mol. Sci. 2022, 23, 5848.
- Ricardo, S.; Marcos-Silva, L.; Valente, C.; Coelho, R.; Gomes, R.; David, L. Mucins MUC16 and MUC1 are major carriers of SLe(a) and SLe(x) in borderline and malignant serous ovarian tumors. Virchows Arch. 2016, 468, 715–722.
- Foley, D.A.; Swartzentruber, K.G.; Colley, K.J. Identification of sequences in the polysialyltransferases ST8Sia II and ST8Sia IV that are required for the protein-specific polysialylation of the neural cell adhesion molecule, NCAM. J. Biol. Chem. 2009, 284, 15505–15516.
- Seki, T.; Arai, Y. Expression of highly polysialylated NCAM in the neocortex and piriform cortex of the developing and the adult rat. Anat. Embryol. 1991, 184, 395–401.
- Li, J.; Yang, R.; Yang, H.; Chen, S.; Wang, L.; Li, M.; Yang, S.; Feng, Z.; Bi, J. NCAM regulates the proliferation, apoptosis, autophagy, EMT, and migration of human melanoma cells via the Src/Akt/mTOR/cofilin signaling pathway. J. Cell. Biochem. 2020, 121, 1192–1204.
- Hauser, M.A.; Kindinger, I.; Laufer, J.M.; Spate, A.K.; Bucher, D.; Vanes, S.L.; Krueger, W.A.; Wittmann, V.; Legler, D.F. Distinct CCR7 glycosylation pattern shapes receptor signaling and endocytosis to modulate chemotactic responses. J. Leukoc. Biol. 2016, 99, 993–1007.
- Verhallen, L.; Lackman, J.J.; Wendt, R.; Gustavsson, M.; Yang, Z.; Narimatsu, Y.; Sorensen, D.M.; Lafferty, K.M.; Gouwy, M.; Marques, P.E.; et al. “Glyco-sulfo barcodes” regulate chemokine receptor function. Cell. Mol. Life Sci. 2023, 80, 55.
- Yabe, U.; Sato, C.; Matsuda, T.; Kitajima, K. Polysialic acid in human milk. CD36 is a new member of mammalian polysialic acid-containing glycoprotein. J. Biol. Chem. 2003, 278, 13875–13880.
- Werneburg, S.; Buettner, F.F.; Erben, L.; Mathews, M.; Neumann, H.; Muhlenhoff, M.; Hildebrandt, H. Polysialylation and lipopolysaccharide-induced shedding of E-selectin ligand-1 and neuropilin-2 by microglia and THP-1 macrophages. Glia 2016, 64, 1314–1330.
- Jarahian, M.; Marofi, F.; Maashi, M.S.; Ghaebi, M.; Khezri, A.; Berger, M.R. Re-Expression of Poly/Oligo-Sialylated Adhesion Molecules on the Surface of Tumor Cells Disrupts Their Interaction with Immune-Effector Cells and Contributes to Pathophysiological Immune Escape. Cancers 2021, 13, 5203.
- Daniotti, J.L.; Lardone, R.D.; Vilcaes, A.A. Dysregulated Expression of Glycolipids in Tumor Cells: From Negative Modulator of Anti-tumor Immunity to Promising Targets for Developing Therapeutic Agents. Front. Oncol. 2015, 5, 300.
- Kasprowicz, A.; Sophie, G.D.; Lagadec, C.; Delannoy, P. Role of GD3 Synthase ST8Sia I in Cancers. Cancers 2022, 14, 1299.
- Harduin-Lepers, A.; Vallejo-Ruiz, V.; Krzewinski-Recchi, M.A.; Samyn-Petit, B.; Julien, S.; Delannoy, P. The human sialyltransferase family. Biochimie 2001, 83, 727–737.
- Rodrigues, E.; Macauley, M.S. Hypersialylation in Cancer: Modulation of Inflammation and Therapeutic Opportunities. Cancers 2018, 10, 207.
- Dobie, C.; Skropeta, D. Insights into the role of sialylation in cancer progression and metastasis. Br. J. Cancer 2021, 124, 76–90.
- Gc, S.; Bellis, S.L.; Hjelmeland, A.B. ST6Gal1: Oncogenic signaling pathways and targets. Front. Mol. Biosci. 2022, 9, 962908.
- Gc, S.; Tuy, K.; Rickenbacker, L.; Jones, R.; Chakraborty, A.; Miller, C.R.; Beierle, E.A.; Hanumanthu, V.S.; Tran, A.N.; Mobley, J.A.; et al. alpha2,6 Sialylation mediated by ST6GAL1 promotes glioblastoma growth. JCI Insight 2022, 7, e158799.
- Smithson, M.; Irwin, R.; Williams, G.; Alexander, K.L.; Smythies, L.E.; Nearing, M.; McLeod, M.C.; Al Diffalha, S.; Bellis, S.L.; Hardiman, K.M. Sialyltransferase ST6GAL-1 mediates resistance to chemoradiation in rectal cancer. J. Biol. Chem. 2022, 298, 101594.
- Duarte, H.O.; Rodrigues, J.G.; Gomes, C.; Hensbergen, P.J.; Ederveen, A.L.H.; de Ru, A.H.; Mereiter, S.; Polonia, A.; Fernandes, E.; Ferreira, J.A.; et al. ST6Gal1 targets the ectodomain of ErbB2 in a site-specific manner and regulates gastric cancer cell sensitivity to trastuzumab. Oncogene 2021, 40, 3719–3733.
- Wichert, B.; Milde-Langosch, K.; Galatenko, V.; Schmalfeldt, B.; Oliveira-Ferrer, L. Prognostic role of the sialyltransferase ST6GAL1 in ovarian cancer. Glycobiology 2018, 28, 898–903.
- Wang, L.; Chen, X.; Wang, L.; Wang, S.; Li, W.; Liu, Y.; Zhang, J. Knockdown of ST6Gal-I expression in human hepatocellular carcinoma cells inhibits their exosome-mediated proliferation- and migration-promoting effects. IUBMB Life 2021, 73, 1378–1391.
- Zhang, M.; Qi, T.; Yang, L.; Kolarich, D.; Heisterkamp, N. Multi-Faceted Effects of ST6Gal1 Expression on Precursor B-Lineage Acute Lymphoblastic Leukemia. Front. Oncol. 2022, 12, 828041.
- Kurz, E.; Chen, S.; Vucic, E.; Baptiste, G.; Loomis, C.; Agrawal, P.; Hajdu, C.; Bar-Sagi, D.; Mahal, L.K. Integrated Systems Analysis of the Murine and Human Pancreatic Cancer Glycomes Reveals a Tumor-Promoting Role for ST6GAL1. Mol. Cell. Proteomics 2021, 20, 100160.
- Britain, C.M.; Holdbrooks, A.T.; Anderson, J.C.; Willey, C.D.; Bellis, S.L. Sialylation of EGFR by the ST6Gal-I sialyltransferase promotes EGFR activation and resistance to gefitinib-mediated cell death. J. Ovarian Res. 2018, 11, 12.
- Liu, Q.; Ma, H.; Sun, X.; Liu, B.; Xiao, Y.; Pan, S.; Zhou, H.; Dong, W.; Jia, L. The regulatory ZFAS1/miR-150/ST6GAL1 crosstalk modulates sialylation of EGFR via PI3K/Akt pathway in T-cell acute lymphoblastic leukemia. J. Exp. Clin. Cancer Res. 2019, 38, 199.
- Kitazume, S.; Imamaki, R.; Ogawa, K.; Komi, Y.; Futakawa, S.; Kojima, S.; Hashimoto, Y.; Marth, J.D.; Paulson, J.C.; Taniguchi, N. Alpha2,6-sialic acid on platelet endothelial cell adhesion molecule (PECAM) regulates its homophilic interactions and downstream antiapoptotic signaling. J. Biol. Chem. 2010, 285, 6515–6521.
- Wang, L.; Li, S.; Yu, X.; Han, Y.; Wu, Y.; Wang, S.; Chen, X.; Zhang, J.; Wang, S. alpha2,6-Sialylation promotes immune escape in hepatocarcinoma cells by regulating T cell functions and CD147/MMP signaling. J. Physiol. Biochem. 2019, 75, 199–207.
- Hait, N.C.; Maiti, A.; Wu, R.; Andersen, V.L.; Hsu, C.C.; Wu, Y.; Chapla, D.G.; Takabe, K.; Rusiniak, M.E.; Bshara, W.; et al. Extracellular sialyltransferase st6gal1 in breast tumor cell growth and invasiveness. Cancer Gene Ther. 2022, 29, 1662–1675.
- Takashima, S.; Tsuji, S.; Tsujimoto, M. Comparison of the enzymatic properties of mouse beta-galactoside alpha2,6-sialyltransferases, ST6Gal I and II. J. Biochem. 2003, 134, 287–296.
- Cheng, J.; Wang, R.; Zhong, G.; Chen, X.; Cheng, Y.; Li, W.; Yang, Y. ST6GAL2 Downregulation Inhibits Cell Adhesion and Invasion and is Associated with Improved Patient Survival in Breast Cancer. Onco Targets Ther. 2020, 13, 903–914.
- Xu, G.; Chen, J.; Wang, G.; Xiao, J.; Zhang, N.; Chen, Y.; Yu, H.; Wang, G.; Zhao, Y. Resveratrol Inhibits the Tumorigenesis of Follicular Thyroid Cancer via ST6GAL2-Regulated Activation of the Hippo Signaling Pathway. Mol. Ther. Oncolytics 2020, 16, 124–133.
- Wu, X.; Zhao, J.; Ruan, Y.; Sun, L.; Xu, C.; Jiang, H. Sialyltransferase ST3GAL1 promotes cell migration, invasion, and TGF-beta1-induced EMT and confers paclitaxel resistance in ovarian cancer. Cell Death Dis. 2018, 9, 1102.
- Chong, Y.K.; Sandanaraj, E.; Koh, L.W.; Thangaveloo, M.; Tan, M.S.; Koh, G.R.; Toh, T.B.; Lim, G.G.; Holbrook, J.D.; Kon, O.L.; et al. ST3GAL1-Associated Transcriptomic Program in Glioblastoma Tumor Growth, Invasion, and Prognosis. J. Natl. Cancer Inst. 2016, 108, djv326.
- Pietrobono, S.; Anichini, G.; Sala, C.; Manetti, F.; Almada, L.L.; Pepe, S.; Carr, R.M.; Paradise, B.D.; Sarkaria, J.N.; Davila, J.I.; et al. ST3GAL1 is a target of the SOX2-GLI1 transcriptional complex and promotes melanoma metastasis through AXL. Nat. Commun. 2020, 11, 5865.
- Lin, W.D.; Fan, T.C.; Hung, J.T.; Yeo, H.L.; Wang, S.H.; Kuo, C.W.; Khoo, K.H.; Pai, L.M.; Yu, J.; Yu, A.L. Sialylation of CD55 by ST3GAL1 Facilitates Immune Evasion in Cancer. Cancer Immunol. Res. 2021, 9, 113–122.
- Sipione, S.; Monyror, J.; Galleguillos, D.; Steinberg, N.; Kadam, V. Gangliosides in the Brain: Physiology, Pathophysiology and Therapeutic Applications. Front. Neurosci. 2020, 14, 572965.
- Zhang, N.; Lin, S.; Cui, W.; Newman, P.J. Overlapping and unique substrate specificities of ST3GAL1 and 2 during hematopoietic and megakaryocytic differentiation. Blood Adv. 2022, 6, 3945–3955.
- Saito, S.; Aoki, H.; Ito, A.; Ueno, S.; Wada, T.; Mitsuzuka, K.; Satoh, M.; Arai, Y.; Miyagi, T. Human alpha2,3-sialyltransferase (ST3Gal II) is a stage-specific embryonic antigen-4 synthase. J. Biol. Chem. 2003, 278, 26474–26479.
- Kannagi, R.; Cochran, N.A.; Ishigami, F.; Hakomori, S.; Andrews, P.W.; Knowles, B.B.; Solter, D. Stage-specific embryonic antigens (SSEA-3 and -4) are epitopes of a unique globo-series ganglioside isolated from human teratocarcinoma cells. EMBO J. 1983, 2, 2355–2361.
- Deschuyter, M.; Leger, D.Y.; Verboom, A.; Chaunavel, A.; Maftah, A.; Petit, J.M. ST3GAL2 knock-down decreases tumoral character of colorectal cancer cells in vitro and in vivo. Am. J. Cancer Res. 2022, 12, 280–302.
- Kono, M.; Ohyama, Y.; Lee, Y.C.; Hamamoto, T.; Kojima, N.; Tsuji, S. Mouse beta-galactoside alpha 2,3-sialyltransferases: Comparison of in vitro substrate specificities and tissue specific expression. Glycobiology 1997, 7, 469–479.
- Okajima, T.; Fukumoto, S.; Miyazaki, H.; Ishida, H.; Kiso, M.; Furukawa, K.; Urano, T.; Furukawa, K. Molecular cloning of a novel alpha2,3-sialyltransferase (ST3Gal VI) that sialylates type II lactosamine structures on glycoproteins and glycolipids. J. Biol. Chem. 1999, 274, 11479–11486.
- Quirino, M.W.L.; Albuquerque, A.P.B.; De Souza, M.F.D.; Da Silva Filho, A.F.; Martins, M.R.; Da Rocha Pitta, M.G.; Pereira, M.C.; De Melo Rego, M.J.B. alpha2,3 sialic acid processing enzymes expression in gastric cancer tissues reveals that ST3Gal3 but not Neu3 are associated with Lauren’s classification, angiolymphatic invasion and histological grade. Eur. J. Histochem. 2022, 66, 3330.
- Rodriguez, E.; Boelaars, K.; Brown, K.; Eveline Li, R.J.; Kruijssen, L.; Bruijns, S.C.M.; van Ee, T.; Schetters, S.T.T.; Crommentuijn, M.H.W.; van der Horst, J.C.; et al. Sialic acids in pancreatic cancer cells drive tumour-associated macrophage differentiation via the Siglec receptors Siglec-7 and Siglec-9. Nat. Commun. 2021, 12, 1270.
- Narimatsu, Y.; Joshi, H.J.; Nason, R.; Van Coillie, J.; Karlsson, R.; Sun, L.; Ye, Z.; Chen, Y.H.; Schjoldager, K.T.; Steentoft, C.; et al. An Atlas of Human Glycosylation Pathways Enables Display of the Human Glycome by Gene Engineered Cells. Mol. Cell 2019, 75, 394–407 e395.
- Sun, M.; Zhao, X.; Liang, L.; Pan, X.; Lv, H.; Zhao, Y. Sialyltransferase ST3GAL6 mediates the effect of microRNA-26a on cell growth, migration, and invasion in hepatocellular carcinoma through the protein kinase B/mammalian target of rapamycin pathway. Cancer Sci. 2017, 108, 267–276.
- Dalangood, S.; Zhu, Z.; Ma, Z.; Li, J.; Zeng, Q.; Yan, Y.; Shen, B.; Yan, J.; Huang, R. Identification of glycogene-type and validation of ST3GAL6 as a biomarker predicts clinical outcome and cancer cell invasion in urinary bladder cancer. Theranostics 2020, 10, 10078–10091.
- Liu, J.; Li, M.; Wu, J.; Qi, Q.; Li, Y.; Wang, S.; Liang, S.; Zhang, Y.; Zhu, Z.; Huang, R.; et al. Identification of ST3GAL5 as a prognostic biomarker correlating with CD8(+) T cell exhaustion in clear cell renal cell carcinoma. Front. Immunol. 2022, 13, 979605.
- Marcos, N.T.; Pinho, S.; Grandela, C.; Cruz, A.; Samyn-Petit, B.; Harduin-Lepers, A.; Almeida, R.; Silva, F.; Morais, V.; Costa, J.; et al. Role of the human ST6GalNAc-I and ST6GalNAc-II in the synthesis of the cancer-associated sialyl-Tn antigen. Cancer Res. 2004, 64, 7050–7057.
- Rajesh, C.; Radhakrishnan, P. The (Sialyl) Tn antigen: Contributions to immunosuppression in gastrointestinal cancers. Front. Oncol. 2022, 12, 1093496.
- Wang, W.Y.; Cao, Y.X.; Zhou, X.; Wei, B.; Zhan, L.; Sun, S.Y. Stimulative role of ST6GALNAC1 in proliferation, migration and invasion of ovarian cancer stem cells via the Akt signaling pathway. Cancer Cell Int. 2019, 19, 86.
- Kvorjak, M.; Ahmed, Y.; Miller, M.L.; Sriram, R.; Coronnello, C.; Hashash, J.G.; Hartman, D.J.; Telmer, C.A.; Miskov-Zivanov, N.; Finn, O.J.; et al. Cross-talk between Colon Cells and Macrophages Increases ST6GALNAC1 and MUC1-sTn Expression in Ulcerative Colitis and Colitis-Associated Colon Cancer. Cancer Immunol. Res. 2020, 8, 167–178.
- Murugesan, G.; Correia, V.G.; Palma, A.S.; Chai, W.; Li, C.; Feizi, T.; Martin, E.; Laux, B.; Franz, A.; Fuchs, K.; et al. Siglec-15 recognition of sialoglycans on tumor cell lines can occur independently of sialyl Tn antigen expression. Glycobiology 2021, 31, 44–54.
- Wang, J.; Sun, J.; Liu, L.N.; Flies, D.B.; Nie, X.; Toki, M.; Zhang, J.; Song, C.; Zarr, M.; Zhou, X.; et al. Siglec-15 as an immune suppressor and potential target for normalization cancer immunotherapy. Nat. Med. 2019, 25, 656–666.
- Murugaesu, N.; Iravani, M.; van Weverwijk, A.; Ivetic, A.; Johnson, D.A.; Antonopoulos, A.; Fearns, A.; Jamal-Hanjani, M.; Sims, D.; Fenwick, K.; et al. An in vivo functional screen identifies ST6GalNAc2 sialyltransferase as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 304–317.
- Ferrer, C.M.; Reginato, M.J. Sticking to sugars at the metastatic site: Sialyltransferase ST6GalNAc2 acts as a breast cancer metastasis suppressor. Cancer Discov. 2014, 4, 275–277.
- Miao, X.; Zhao, Y. ST6GalNAcII mediates tumor invasion through PI3K/Akt/NF-kappaB signaling pathway in follicular thyroid carcinoma. Oncol. Rep. 2016, 35, 2131–2140.
- Reticker-Flynn, N.E.; Bhatia, S.N. Aberrant glycosylation promotes lung cancer metastasis through adhesion to galectins in the metastatic niche. Cancer Discov. 2015, 5, 168–181.
- Adams, O.J.; Stanczak, M.A.; von Gunten, S.; Laubli, H. Targeting sialic acid-Siglec interactions to reverse immune suppression in cancer. Glycobiology 2018, 28, 640–647.
- Dai, T.; Li, J.; Liang, R.B.; Yu, H.; Lu, X.; Wang, G. Identification and Experimental Validation of the Prognostic Significance and Immunological Correlation of Glycosylation-Related Signature and ST6GALNAC4 in Hepatocellular Carcinoma. J. Hepatocell. Carcinoma 2023, 10, 531–551.
- Ikehara, Y.; Shimizu, N.; Kono, M.; Nishihara, S.; Nakanishi, H.; Kitamura, T.; Narimatsu, H.; Tsuji, S.; Tatematsu, M. A novel glycosyltransferase with a polyglutamine repeat; a new candidate for GD1alpha synthase (ST6GalNAc V)(1). FEBS Lett. 1999, 463, 92–96.
- Okajima, T.; Fukumoto, S.; Ito, H.; Kiso, M.; Hirabayashi, Y.; Urano, T.; Furukawa, K. Molecular cloning of brain-specific GD1alpha synthase (ST6GalNAc V) containing CAG/Glutamine repeats. J. Biol. Chem. 1999, 274, 30557–30562.
- Dai, J.; Li, Q.; Quan, J.; Webb, G.; Liu, J.; Gao, K. Construction of a lipid metabolism-related and immune-associated prognostic score for gastric cancer. BMC Med. Genom. 2023, 16, 93.
- Kroes, R.A.; He, H.; Emmett, M.R.; Nilsson, C.L.; Leach, F.E., 3rd; Amster, I.J.; Marshall, A.G.; Moskal, J.R. Overexpression of ST6GalNAcV, a ganglioside-specific alpha2,6-sialyltransferase, inhibits glioma growth in vivo. Proc. Natl. Acad. Sci. USA 2010, 107, 12646–12651.
- Miyazaki, K.; Ohmori, K.; Izawa, M.; Koike, T.; Kumamoto, K.; Furukawa, K.; Ando, T.; Kiso, M.; Yamaji, T.; Hashimoto, Y.; et al. Loss of disialyl Lewis(a), the ligand for lymphocyte inhibitory receptor sialic acid-binding immunoglobulin-like lectin-7 (Siglec-7) associated with increased sialyl Lewis(a) expression on human colon cancers. Cancer Res. 2004, 64, 4498–4505.
- Yeh, S.C.; Wang, P.Y.; Lou, Y.W.; Khoo, K.H.; Hsiao, M.; Hsu, T.L.; Wong, C.H. Glycolipid GD3 and GD3 synthase are key drivers for glioblastoma stem cells and tumorigenicity. Proc. Natl. Acad. Sci. USA 2016, 113, 5592–5597.
- Hao, J.; Zeltz, C.; Pintilie, M.; Li, Q.; Sakashita, S.; Wang, T.; Cabanero, M.; Martins-Filho, S.N.; Wang, D.Y.; Pasko, E.; et al. Characterization of Distinct Populations of Carcinoma-Associated Fibroblasts from Non-Small Cell Lung Carcinoma Reveals a Role for ST8SIA2 in Cancer Cell Invasion. Neoplasia 2019, 21, 482–493.
- Gong, L.; Zhou, X.; Yang, J.; Jiang, Y.; Yang, H. Effects of the regulation of polysialyltransferase ST8SiaII on the invasiveness and metastasis of small cell lung cancer cells. Oncol. Rep. 2017, 37, 131–138.
- Ma, X.; Dong, W.; Su, Z.; Zhao, L.; Miao, Y.; Li, N.; Zhou, H.; Jia, L. Functional roles of sialylation in breast cancer progression through miR-26a/26b targeting ST8SIA4. Cell Death Dis. 2016, 7, e2561.
- Zhang, Z.; Zhao, Y.; Jiang, L.; Miao, X.; Zhou, H.; Jia, L. Glycomic alterations are associated with multidrug resistance in human leukemia. Int. J. Biochem. Cell Biol. 2012, 44, 1244–1253.
- Baeza-Kallee, N.; Berges, R.; Souberan, A.; Colin, C.; Denicolai, E.; Appay, R.; Tchoghandjian, A.; Figarella-Branger, D. Glycolipids Recognized by A2B5 Antibody Promote Proliferation, Migration, and Clonogenicity in Glioblastoma Cells. Cancers 2019, 11, 1267.
- Penrose, H.M.; Cable, C.; Heller, S.; Ungerleider, N.; Nakhoul, H.; Baddoo, M.; Hartono, A.B.; Lee, S.B.; Burow, M.E.; Flemington, E.F.; et al. Loss of Forkhead Box O3 Facilitates Inflammatory Colon Cancer: Transcriptome Profiling of the Immune Landscape and Novel Targets. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 391–408.
- Huang, R.; Zheng, Z.; Xian, S.; Zhang, J.; Jia, J.; Song, D.; Yan, P.; Yin, H.; Hu, P.; Zhu, X.; et al. Identification of prognostic and bone metastatic alternative splicing signatures in bladder cancer. Bioengineered 2021, 12, 5289–5304.
- Friedman, D.J.; Crotts, S.B.; Shapiro, M.J.; Rajcula, M.; McCue, S.; Liu, X.; Khazaie, K.; Dong, H.; Shapiro, V.S. ST8Sia6 Promotes Tumor Growth in Mice by Inhibiting Immune Responses. Cancer Immunol. Res. 2021, 9, 952–966.
- Wu, Z.; He, L.; Yang, L.; Fang, X.; Peng, L. Potential Role of NEU1 in Hepatocellular Carcinoma: A Study Based on Comprehensive Bioinformatical Analysis. Front. Mol. Biosci. 2021, 8, 651525.
- Qorri, B.; Harless, W.; Szewczuk, M.R. Novel Molecular Mechanism of Aspirin and Celecoxib Targeting Mammalian Neuraminidase-1 Impedes Epidermal Growth Factor Receptor Signaling Axis and Induces Apoptosis in Pancreatic Cancer Cells. Drug Des. Dev. Ther. 2020, 14, 4149–4167.
- Ren, L.R.; Zhang, L.P.; Huang, S.Y.; Zhu, Y.F.; Li, W.J.; Fang, S.Y.; Shen, L.; Gao, Y.L. Effects of sialidase NEU1 siRNA on proliferation, apoptosis, and invasion in human ovarian cancer. Mol. Cell. Biochem. 2016, 411, 213–219.
- Peng, Q.; Gao, L.; Cheng, H.B.; Wang, J.S.; Wang, J. Sialidase NEU1 May Serve as a Potential Biomarker of Proliferation, Migration and Prognosis in Melanoma. World J. Oncol. 2022, 13, 222–234.
- Zhou, X.; Zhai, Y.; Liu, C.; Yang, G.; Guo, J.; Li, G.; Sun, C.; Qi, X.; Li, X.; Guan, F. Sialidase NEU1 suppresses progression of human bladder cancer cells by inhibiting fibronectin-integrin alpha5beta1 interaction and Akt signaling pathway. Cell Commun. Signal. 2020, 18, 44.
- Garcia-Dominguez, D.J.; Hajji, N.; Lopez-Alemany, R.; Sanchez-Molina, S.; Figuerola-Bou, E.; Moron Civanto, F.J.; Rello-Varona, S.; Andres-Leon, E.; Benito, A.; Keun, H.C.; et al. Selective histone methyltransferase G9a inhibition reduces metastatic development of Ewing sarcoma through the epigenetic regulation of NEU1. Oncogene 2022, 41, 2638–2650.
- Nath, S.; Mondal, S.; Butti, R.; Prasanna Gunasekaran, V.; Chatterjee, U.; Halder, A.; Kundu, G.C.; Mandal, C. Desialylation of Sonic-Hedgehog by Neu2 Inhibits Its Association with Patched1 Reducing Stemness-Like Properties in Pancreatic Cancer Sphere-forming Cells. Cells 2020, 9, 1512.
- Nath, S.; Mandal, C.; Chatterjee, U.; Mandal, C. Association of cytosolic sialidase Neu2 with plasma membrane enhances Fas-mediated apoptosis by impairing PI3K-Akt/mTOR-mediated pathway in pancreatic cancer cells. Cell Death Dis. 2018, 9, 210.
- Satyavarapu, E.M.; Nath, S.; Mandal, C. Desialylation of Atg5 by sialidase (Neu2) enhances autophagosome formation to induce anchorage-dependent cell death in ovarian cancer cells. Cell Death Discov. 2021, 7, 26.
- Iioka, H.; Saito, K.; Kondo, E. Crumbs3 regulates the expression of glycosphingolipids on the plasma membrane to promote colon cancer cell migration. Biochem. Biophys. Res. Commun. 2019, 519, 287–293.
- Zhang, X.; Dou, P.; Akhtar, M.L.; Liu, F.; Hu, X.; Yang, L.; Yang, D.; Zhang, X.; Li, Y.; Qiao, S.; et al. NEU4 inhibits motility of HCC cells by cleaving sialic acids on CD44. Oncogene 2021, 40, 5427–5440.
- Shiozaki, K.; Yamaguchi, K.; Takahashi, K.; Moriya, S.; Miyagi, T. Regulation of sialyl Lewis antigen expression in colon cancer cells by sialidase NEU4. J. Biol. Chem. 2011, 286, 21052–21061.
More
Information
Subjects:
Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.1K
Revisions:
2 times
(View History)
Update Date:
25 Jun 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No