Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Robert Wieder | -- | 4438 | 2023-06-19 14:53:06 | | | |
| 2 | Sirius Huang | Meta information modification | 4438 | 2023-06-20 03:25:10 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Wieder, R. Breast Cancer Dormancy in the Bone Marrow. Encyclopedia. Available online: https://encyclopedia.pub/entry/45797 (accessed on 21 June 2026).
Wieder R. Breast Cancer Dormancy in the Bone Marrow. Encyclopedia. Available at: https://encyclopedia.pub/entry/45797. Accessed June 21, 2026.
Wieder, Robert. "Breast Cancer Dormancy in the Bone Marrow" Encyclopedia, https://encyclopedia.pub/entry/45797 (accessed June 21, 2026).
Wieder, R. (2023, June 19). Breast Cancer Dormancy in the Bone Marrow. In Encyclopedia. https://encyclopedia.pub/entry/45797
Wieder, Robert. "Breast Cancer Dormancy in the Bone Marrow." Encyclopedia. Web. 19 June, 2023.
Copy Citation
Breast cancer cells travel via the bloodstream to the bone before the cancer is detectable in the breast. These disseminated cells are resistant to adjuvant chemotherapy and hormone therapy administered for the very purpose of eliminating them. They recur steadily for more than 20 years, resulting in incurable diseases. The bone marrow location, or niche, which normally provides a nest for blood-forming cells to enable them to generate blood for the entire lifetime of an individual, also protects these disseminated tumor cells and places them into a state of quiescence called dormancy.
dormancy
micrometastases
hematopoietic niche
bone marrow microenvironment
1. Introduction
More than 43,000 women in the US die from breast cancer (BC) every year, primarily from metastatic disease [1]. However, BC cells metastasize to the bone marrow (BM) before primary tumors can be detected and are found in the BM of 27–40% of newly diagnosed patients with localized primary disease [2][3]. Once they arrive at the BM, most micrometastases are killed by the hostile microenvironment; nevertheless, some of them enter a state of dormancy [4]. Dormant cells have cancer stem cell characteristics [5] and are resistant to adjuvant chemotherapy administered for the distinct purpose of eliminating them [6][7]. The metastatic niche contributes significantly to this resistance [8][9].
2. Breast Cancer Metastasis and Dormancy in the Bone Marrow
2.1. Hematogenous Transit of Cancer Cells to the BM HSC Niches
The homing of cancer cells to the BM is reported to approximate that of HSCs [10]. It is mediated through the C-X-C motif chemokine receptor 4 (CXCR4), the receptor for stromal cell-derived factor-1/C-X-C motif chemokine 12 (SDF-1/CXC ligand (CXCL)12) [11][12][13] and through annexin II [14], which is required for hematopoietic stem cell transplants [15]. Annexin II serves as an anchor for CXCL12 to localize HSCs [16] and cancer cells [17] to the niche. Homing also involves cadherin-11 [18][19], osteopontin [20], connective tissue growth factor (CTGF) [20], and Runt-related transcription factor 2 (RUNX2) [21]. Cadherin-11 induces the expression of the gamma-carboxyglutamic acid (Gla) domain-containing protein 6 (GAS6) receptors AXL tyrosine kinase (AXL), skywalker (Sky) and Mer proto-oncogene tyrosine kinase (Mer), which induce dormancy in hematopoietic stem cells [22].
Disseminated tumor cells (DTCs) in the BM interact with a wide array of cell types, proteins, proteoglycans, growth factors and cytokines endemic to the hematopoietic microenvironment, which, together with its biophysical and bioenergetic characteristics, regulate dormancy and participate in reawakening [23]. The BM hematopoietic microenvironment is made up of a complex network of cells consisting of mesenchymal cells of different lineages and degrees of stemness, osteogenic cells, chondrocytes, adipocytes, neuroglial cells, hematopoietic lineage cells, including megakaryocytes and macrophages, cells of the sympathetic nervous system and a network of endothelial cells that include cells lining the sinusoids, arterioles and transition zones [24]. These data indicate the existence of two primary niches that maintain HSC dormancy, the preosteoblast endosteal-lining niche [25] and a more centrally located parasinusoidal endothelial niche, with some endothelial cells also residing near the endosteum [26]. The two positions have been reconciled by arguments that the endosteal niche is also vascularized [27], that the niches are in close proximity and that, because of the newly recognized heterogeneity in the HSC population gleaned from single-cell sequencing, different niches may provide support or imprint distinct HSC states for differently primed HSCs [24][28][29][30][31].
The endosteal niches consist of osteoblasts embedded in the bone matrix and preosteoblasts that are in contact with HSCs and Nestin-GFP+hi mesenchymal stem cells (MSCs), which regulate HSC maintenance. Macrophages also inhabit the endosteum and help maintain the HSC niche. The sympathetic nervous system maintains HSCs through nerve fibers in the endosteum. Structural elements of the endosteum also support the dormancy of HSCs, including fibronectin and heparan sulfate proteoglycans (HSPGs) that serve as reservoirs for fibroblast growth factor (FGF)-2, which is important in HSC dormancy (Figure 1). I have discussed these members of the HSC niche below.
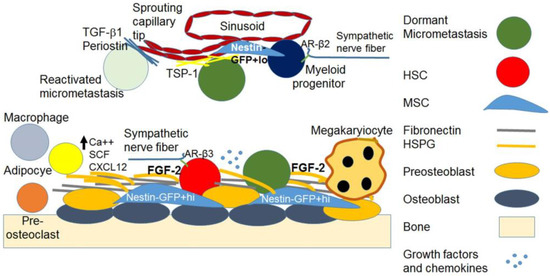
Figure 1. The bone marrow hematopoietic and dormant metastatic breast cancer niches. The image depicts simplified representations of the endosteal and parasinusoidal niche containing the HSC, the dormant micrometastasis, and the supporting cellular, structural and soluble elements. Growth factors and chemokines listed in the text are depicted as a cluster of light blue dots.
The true, rare, dormant and undifferentiated HSCs reside on preosteoblasts in the endosteal stromal and preosteoblast stem cell niches, which support their quiescence and self-renewal [32][33]. Once they acquire a lineage marker, they move to the sinusoidal endothelium where they can be tapped to differentiate into myeloid lineages [32][33].
Only immature osteoblasts support the dormancy of HSCs [34], whereas ossified, alkaline phosphatase (ALP)-expressing differentiated osteoblasts no longer support HSCs [35][36][37][38]. Signaling through parathyroid hormone (PTH) and interleukin (IL)-6, and adhesion to vascular cell adhesion molecule (VCAM)-1 and activated leukocyte cell adhesion molecule (CD166) in osteoblast precursors is necessary for appropriate HSC maintenance and lymphoid differentiation [35][36][37]. An array of adhesion molecules, growth factors and chemokines interact to maintain HSCs in a quiescent state in their niche and to mobilize them as needed for recruitment to the vascular niche [33]. These include Angiopoietin-1, Tie-2 and N-cadherin, which are associated with quiescence and cell-cycle control and adhesion molecules, including very late antigen (VLA)-4, lymphocyte function-associated (LFA)-1, osteopontin and integrins [33]. These molecules, along with CXCL12 (SDF-1) on stromal cells, its receptor CXCR-4 on HSCs, matrix metalloproteinase (MMP)-9, MMP-2 and stem cell factor (Kit Ligand) induced by granulocyte-colony stimulating factor (G-CSF), SDF-1, FGF-4, vascular endothelial growth factor (VEGF) and placental growth factor (PLGF), are required for HSC recruitment and mobilization [33]. Calcium in the endosteal niche is also important for the support of HSCs [39]. HSC maintenance and quiescence, hematopoietic reconstitution and protection from aging-associated DNA damage also depend on interactions with periostin via integrin αv and inhibition of focal adhesion kinase (FAK)/phosphoinositide 3-kinase (PI3K)/Ak strain transforming (AKT) signaling, leading to an increase in p27Kip1 [40].
The sympathetic nervous system β2 adrenergic signaling releases HSCs from the osteoblastic niche by inducing the transcription of the calcium-regulating hormone vitamin D receptor (VDR) and its downstream gene receptor activator of nuclear factor kappa-Β ligand (RANKL), an effect stabilized by 1,25 dihydroxy-vitamin D3 [41]. While the mean extracellular calcium ion concentration [Ca++]e in the BM is 1.0 ± 0.54 mM, which is not significantly different from that in the blood serum, the [Ca++] in the location of the HSCs is 1.5 ± 0.57 mM and significantly increases with aging to support myeloid clonal expansion [42].
Aging and senescence of HSC niches result in changes in the character, makeup, location and differentiation dynamics of HSCs by regulating niche remodeling [28]. Aging induces a functional decrease in adrenergic receptor (AR)-β3 activation and increases AR-β2 (ADRB2) activation [28][33][43]. This induces HSC migration from osteoblasts to the sinusoids, skewing hematopoiesis towards myeloid differentiation, megakaryocyte and platelet production, and decreases endosteal cells, resulting in lymphoid deficiency [28][33][43]. Aging stroma results in a decrease in endosteal and in an increase in sinusoidal Nestin-GFP+hi cells, events that also play a role in the myeloid skewing of hematopoiesis. The movement of Nestin-GFP+hi cells, which give rise to all mesenchymal lineages, including osteoblasts and adipocytes, to the sinusoidal space induces the migration of Jagged-stained cells and associated HSCs to the sinusoids [28][44].
There is a global decrease in osteoblastogenesis and an increase in adipocytogenesis with aging, ovariectomy and other causes of osteoporosis or disease [45][46][47][48]. Indeed, reinforcing the role of adipocytes in the loss of dormancy, perivascular cells express an adipocyte-skewed expression profile that promotes proliferation [49]. Mature adipocytes activate extracellular receptor kinase (ERK or MAP kinase) signaling in multiple myeloma cells [50]. Overall vascular density and leakiness increase and sinusoidal notch receptor (Notch) activity decreases with aging [28]. These effects are accompanied by an overall increase in inflammation and secretory senescence, characterized by increased levels of inflammatory cytokines IL-1, IL-3, IL-6, tumor necrosis factor (TNF)α, interferon (INF)γ and transforming growth factor (TGF)-β, which regulate myeloid skewing [28].
The role of osteoblasts in the maintenance of the undifferentiated quiescent HSC state is supported by experiments in which a preosteoblast knockout was sufficient to induce leukemia [51][52][53], and its replacement restored normal hematopoiesis [54]. Many adhesion molecules, growth factors and chemokines interact to maintain HSCs quiescence in their niche and mobilize them to the vascular niche as needed [33][35][36][37]. These include bone morphogenic proteins (BMPs), TGF-β2 [32] and FGF-2 [55][56][57]. FGF-2 is important for the maintenance [55], self-renewal [56][57][58] and myeloid differentiation of HSCs [59][60], but blocks erythroid and B lymphocyte differentiation [61] and myeloid differentiation at high concentrations [62].
2.2. The Metastatic BM Niches and DTC Dormancy Signaling
Most metastases die in the hostile microenvironment of the BM, but some survive and enter a state of dormancy [63]. Early arriving DTCs that survive the microenvironmental effects generate a metastatic niche, combine with late metastases and potentially remain quiescent or in an ultra-slow cycling mesenchymal state in the HSC niche [64][65][66] for periods lasting up to decades [32][67].
The fate of the cancer cells depends on the opposing efforts of the microenvironment and the cancer cells. The microenvironment endeavors to suppress the cancer cells, while the cancer cells exert their efforts to turn MSCs and fibroblasts into cancer-promoting cells [68]. The cancer cells attempt to generate a pre-metastatic niche with the potential to support cancer cell colonization through the modulation of MSC through microvesicles [69] and through the secretion of inflammatory cytokines, which recruit BM-derived cells and form an inflammatory milieu that supports colonization [70]. They also secrete factors that enhance bone resorption, such as lysyl oxidase (LOX), a collagen crosslinking enzyme produced primarily by hypoxic ER− cancer cells, parathyroid hormone-related peptide (PTHrP), osteopontin (OPN) and CC-chemokine ligand (CCL)-2, directly promoting bone resorption and extracellular matrix (ECM) remodeling, making the niche more permissive to DTCs [71]. However, metastatic cancer cells also process structural proteins such as fibronectin in the microenvironment, which in turn induce quiescence and survival signaling in the cancer cells [67]. Ultimately, the scarcity of micrometastases is the most likely reason why the cancer cells’ attempts at modifying the niche to promote cancer growth are overwhelmed by the collective suppressive effects of the cellular, structural and soluble factors of the niche [68]. DTCs in the BM interact with a wide array of cell types, proteins, proteoglycans, growth factors and cytokines endemic to the hematopoietic microenvironment, which, together with their biophysical and bioenergetic characteristics, regulate dormancy and eventually participate in reawakening [23].
Cancer cell dormancy in a niche can be considered an adaptive state, guided by the thermodynamics of local energy minima, mechanical confinement [72] and hypoxia [73]. The preparation of the metastatic niche may be aided by the presence of VEGF receptor (VEGFR)1+ hematopoietic progenitor cells [74], as well as the deposition of extracellular matrix by micrometastases [75]. In the metastatic HSC niche, BC cells interact with cellular, structural and soluble factors to initiate dormancy [74][76][77][78][79][80], including NG2+/Nestin+ mesenchymal stem cell-initiated TGF-β2 and BMP7 signaling [81]. Cells include MSCs, fibroblasts, osteoblasts, adipocytes, Nestin+ endothelial cells, T-cells and macrophages. Structural factors include fibronectin, p-selectin, thrombospondin and HSPG. Soluble factors include Bmp4, Bmp6, Bmp7, kit ligand, TGF-β1 and β2; Dickkopf-related protein 1 (Dkk1) and Dkk3, thombospondin2 (Thbs2) found in the BM secretome [82][83] and FGF-2 [74][76][77][78][79][84]. FGF-2 is deposited on stromal HSPGs [85], which are needed for FGFR dimerization [86] and are able to induce dormancy [87]. Fibronectin, an integral element of the endosteum [88], also induces dormancy [89][90] and can prevent transformation [91]. Signaling initiated by structural proteins in the BM also depends on their variable structural organization [89], tensile strength and mechanical signaling [92].
Dormant BM micrometastases have marked genetic heterogeneity [93][94]. However, most express the hyaluronan receptor CD44 and about half of the cells are also CD24−, identifying them as having tumor-initiating characteristics that enable them to regrow into breast tumors [95] and express a stem cell program [96]. Signaling initiated by osteoblast interactions seems to maintain tumor-initiating properties in DTCs [97].
Maintenance of stemness was also reported to be mediated by FGFR2 [98] and FGFR-initiated signaling through Akt/Sry-related HMG-box (Sox)2 [99], stem cell-like chromatin rearrangement through the inhibition of cyclin-dependent kinase (CDK)4/6 and upregulation of programmed cell death protein (PD)-1 [100][101], protecting dormant cells from immune elimination. A key niche factor for inducing and maintaining ER+ BC dormancy is FGF-2, which is synthesized and exported by stromal fibroblasts and subsequently deposited on HSPGs overlying the stroma [84][85]. FGF-2 also plays a key role in the maintenance of HSC dormancy, as noted above [98][99]. HSPGs are necessary for the dimerization of FGF receptors [86] and for maintenance [102], multipotency [103] and osteogenic differentiation of MSCs [104]. Similarly to its role in HSCs, FGF-2 also supports the dormancy of hormone receptor-positive BC cells through dual signaling by FGF-2-induced re-expression of integrin α5β1, which binds to microenvironmental fibronectin [84][105][106][107]. FGF-2 inhibits breast cancer cell proliferation and response to chemotherapy through the activation of ERK [108], phosphoinositol-3 kinase (PI3K) [84][105] and intracellular TGF-β-mediated upregulation of cyclin-dependent kinase inhibitors p21Waf1, p27Kip1 and 15INK4b [109][110], inactivation of CDK2 and CDK4, and dephosphorylation of retinoblastoma protein (Rb) [109], mechanisms which have been confirmed in subsequent investigations [100][101]. Dormant cells maintain a characteristic, large, spread out, non-motile epithelial phenotype through dual FGF-2 and fibronectin-activated integrin α5β1 signaling [106]. The phenotype is due to the inhibition of Ras homology family member (Rho)A by the Rho GTPase activating protein (Gap) 26 (GRAF), a resulting cortical actin rearrangement and an omnidirectional activation of FAK [106]. Fibronectin, which is deposited abundantly in the BM microenvironment [88], suppresses the malignant phenotype [89][91] and collaborates with integrin α5β1 to establish the premetastatic niche [67]. ER− cells are not inhibited by FGF-2 and do not utilize the fibronectin-FGF-2 dual signaling model to become dormant [84]. However, stromal MSCs do inhibit ER− BC cells in a transwell co-culture model through the transfer of micro (mi)RNAs 127, -197, -222 and -223 [111][112][113] or SDF-1a [114] and decrease in CXCL12 levels [112]. This interaction is reciprocal, as metastatic breast cancer cells in the bone marrow microenvironment participate in remodeling the niche to sustain their dormancy [115]. In addition to its role in BC [116], FGF-2 also promotes stemness in benign prostate cells [98] and pancreatic cancer [99], and induces dormancy in ER+ BC cells [84]. Quiescent cancer micrometastases express dormancy signatures [64][65][117] similar to those modulating normal stem cell quiescence [64][66]. FGF-2 acts in concert with structural proteins in the microenvironment, where dormant micrometastases become anchored in place by binding to microenvironmental proteins and cellular components [84][90].
The MSC niche, the vascular niche and the immune niche provide support for metastatic BC cell survival and dormancy through a variety of mechanisms (Table 1) [32]. Metastatic cells survive in the BM hematopoietic microenvironment in close proximity to stromal cells in the endosteum, where they occupy the hematopoietic stem cell niche [118], as well as in the perivascular endothelium [8]. The mesenchymal stem cell niche activates multiple signaling pathways in dormant cells via receptors Mer tyrosine kinase (MERTK), AXL and its ligand GAS6 [22][119], TGF-β2 through TGF-β receptor 3 and BMP receptor 2 via SMAD family members (SMAD)1 and 5, basic helix–loop–helix family member E41 (DEC2), the metastasis suppressor gene N-Myc downstream-regulated (NDRG)1, BMP4 and 7 through BMP receptor (R)2, activated p38 MAP kinase (p38) [76] and its downstream target mitogen- and stress-activated kinase 1 (MSK1) [120], activin receptor-like kinase (Alk)5 [121], inactivated ERK, and cyclin-dependent kinase inhibitors p21Waf1 and p27Kip1 [32][80][122]. Other stem cell niche signals also regulate dormancy [74][76][77][79], potentially by antagonizing oncogene signaling [78].
The BM is a hypoxic environment [123], a factor implicated in the induction of dormancy [124] by repressing the leukemia inhibitory factor (LIF)-signal transducer and activator of transcription (STAT)3. Primary tumor hypoxia presets primary tumor cells with a program supporting dormancy, which manifests after dissemination to the metastatic niche [125]. Redox signaling in the microenvironment also generates enabling conditions for dormancy signaling, remodeling of the microenvironment, reprogramming of DTC dormancy signaling and maintenance of epithelial-mesenchymal transition (EMT) and stemness [126]. Microenvironmental redox signaling also generates therapeutic resistance in dormant cells through vigorous induction of antioxidant mechanisms to counter cytotoxin-induced oxidative stress, apoptosis, autophagy and oncogenic bypass signaling [126]. Conversely, redox signaling can also play a role in reawakening [126].
Other factors involved in inducing dormancy are retinoic acid, leukemia inhibitory factor (LIF), wingless-related integration site (Wnt) family members, miR-126 and DNA methylation (reviewed by Risson et al., 2020) [80]. Stroma also produces exosomes overexpressing miR-23b [111] or other miRNAs [112][113] that are transferred to DTCs, which also endow dormancy signaling [82][111][112][127]. Wnt5a non-canonical Wnt signaling induces dormancy in prostate cancer cells in vitro and in vivo in the BM osteoblast niche in a reversible manner via receptor tyrosine kinase-like orphan receptor 2 (ROR2)-activation of siah E3 ubiquitin protein ligase 2 (SIAH2) expression, which represses canonical Wnt/β-catenin tumor stem cell and tumor progression signaling [128]. Wnt family members regulate MSCs in their niche in the BM stroma, where Wnt5a localizes with cells that are leucocyte common antigen (CD45)+, which are transmembrane protein tyrosine phosphatases located on most hematopoietic cells, and CD45− mesenchymal stem cell marker (STRO-1)+ mesenchymal progenitor cells, whereas canonical Wnt is associated with the underlying stroma matrix [129]. Wnt3a expands the pool of MSCs capable of generating colony-forming unit-fibroblasts (CFU-F) and CFU-osteoblasts (O), whereas Wnt5a maintains the pool of cell numbers, CFU-Fs and CFU-Os, suggesting a potential dual role of Wnt5a in the maintenance of MSCs in the BM and in enhancing osteogenesis [129].
The BM microenvironment has a low oxygen tension, which predisposes cancer cells to fuse with MSCs and other cells in the microenvironment, and, in fact, the fusion of BC cells with MSCs can induce dormancy [130], as well as a spectrum of other functions in BC cells [131]. One study suggests that cancer cells cannibalize stromal mesenchymal cells to become dormant [132]. The BM interstitial pH ranges from 6.7 to 7.5 (7.0–7.3 within a 10% to 90% confidence interval), with a mean value of 7.1, which is slightly more acidic than the blood serum that is close to 7.4 [42]. The BM oxygen tension is <1–6% (~7 mm Hg–43 mm Hg), as compared to most normal tissues of 2% to 9% (14–65 mm Hg) [73]. This hypoxic, acidic microenvironment generates a redox imbalance, which, combined with a slightly hypertonic medium and TGF-β and BMP signaling, is sufficient to drive cancer cells to a gene expression pattern with characteristic features of the dormancy signature [133].
In the osteoblast niche, data suggest that metastatic malignant cells usurp the HSC niche to create an abnormal niche that is unable to support normal hematopoiesis [49][118][122][134]. BC cells with a stem cell phenotype compete with HSCs in the endosteal niche and remain dormant in a Notch-dependent manner by spindle-shaped N-cadherin+ CD45− osteoblasts (SNO cells) [135]. Micrometastases survive in the HSC niche close to endosteal stromal cells [118], where preosteoblasts support their survival and chemoresistance [9], partially mediated by Jagged1 [136]. As noted, it is the preosteoblasts that likely support dormancy [34], but as osteoblasts differentiate, they connect with cancer cells through gap junctions, increase their intracellular calcium levels and potentially promote colonization [39]. Jagged-1/Notch signaling regulates tumor stem cell development, epithelial-to-mesenchymal transition, and immune cell homeostasis during minimal residual disease, and plays a role in the recurrence of minimal residual disease in primary tumors [137]. However, some osteoblasts in the HSC niche become “educated” by arriving cancer cells to support the dormant state [138]. These “educated” osteoblasts express RUNX2/osteocalcin (OCN)/OPN, are negative for IL-6 and α-smooth muscle actin (αSMA), and have new properties where they acquire the capacity to suppress both triple-negative and ER+ breast cancer cell proliferation [139]. They increase cancer cell p21Waf1 expression [138], regulate ERK 1 and 2 signaling and inhibit S-phase entry [139]. These effects are mediated by the secretion of small extracellular vesicles enriched for miR-148a-3p [139]. These data underscore the reciprocal relationship between the cancer cells educating the metastatic microenvironment in the premetastatic and dormancy niches, and the dormancy-endowing effects of the niche on cancer cells.
In the vascular niche, non-sprouting endothelial cells produce ECM components such as thrombospondin-1 (TSP1), which may induce dormancy [32]. The endothelial Duffy antigen receptor for chemokines (DARC) may induce dormancy in cancer cells by binding to the metastasis suppressor cluster of differentiation 82 (KAI1), inhibiting proliferation through p21Waf1 and downregulating T-Box transcription factor 2 (TBX2) [140]. Signaling mechanisms associated with micrometastasis dormancy include von Willebrand factor (vWF) [8], VCAM1 [8], CXCL 1 and 2 [141], BMP7 [76], TGFβ-2 [77], canonical nuclear factor (NF)κB combined with ER signaling in ER+ BC cells [142], nuclear receptor subfamily 2 group F member (NR2F)1 [79] and zing finger protein (ZFP)281 [143]. Dormant stem cell signaling through phosphatase and tensin homolog (PTEN) maintains a dominant role in tumorigenic stimuli [78]. Perivascular periaxin (Prx)1+ MSCs express CXCL12 and maintain quiescence and chemoresistance of leukemic stem cells, in contrast to their effects on HSCs, suggesting a more complex mechanism that differentiates the roles of the endothelial niche in malignant vs. normal hematopoietic stem cell maintenance [144].
However, signaling in the vascular endothelial niche is not all dormancy-inducing in malignant cells [145]. Analogous to the case of hematopoietic stem cells that receive pro-differentiating signals once they translocate to the endothelial niche, cancer cell micrometastases can receive context-specific proliferative signals in their interactions with endothelial cell tips mediated through TGF-β1 and periostin [145]. Indeed, the effects of periostin appear to be context-specific, as some of its effects on HSC are linked to stem cell maintenance in the endosteal niche, as noted above [40]. Endothelial cells can promote a stem-like phenotype in some solid tumor cancer cells through the activation of the hedgehog pathway through Gli-1 [146]. Gli-1 expression is high in breast cancer and contributes to therapeutic resistance in both ER+ [147] and ER− BC cells [148][149] through Wnt signaling [148]. Endothelial cell L1 cell adhesion molecule (L1CAM) ligands may induce the proliferation of L1CAM+ DTCs [80]. Endothelial leukocyte adhesion molecule 1 (E-selectin) signaling in endothelial cells induces a non-canonical mesenchymal–epithelial transition (MET) phenotype in cancer cells, which begin to express EpCam and cytokeratin 14 (CK14) while continuing to express mesenchymal gene expression factors including snail family transcriptional repressor (Snail) 1/2, twist family bHLH transcription factor (Twist) 1/2, zinc finger E-box binding homeobox (Zeb) 1/2 and cancer stem cell marker Sox 2/9 [150]. These programs permit the regrowth of dormant micrometastases [150]. The conditions for the recurrence of cells expressing mesenchymal programs are discussed below [107]. Quiescent DTCs in the BM lack the epithelial marker E-cadherin [151], but do not undergo a phenotypic appearance of EMT [152]. This is corroborated by in vitro data supporting a model for continued mesenchymal signaling in dormant cells with an apparent epithelial phenotype [106][107], as discussed below. However, once these cells are stimulated to undergo MET, they begin to proliferate once again. Micrometastatic sites can serve as launching pads for colonization and re-metastasis [153].
The immune niche contains macrophages and CD4+ and CD8+ cells that may induce dormancy [152] through INFγ [32]. Quiescent cancer cells in distant organs that have tumor-initiating capacity express DKK-1, which inhibits Wnt, enhancing the downregulation of Natural Killer (NK) cell activators and death ligands, and evading killing by NK cells [154]. In addition to homing, SDF-1/CXCR4 may promote survival through Src through Akt and TNF resistance through TNF-related apoptosis-inducing ligand (TRAIL) [155]. Secretion of SDF-1α by BM MSCs may maintain quiescence in breast cancer micrometastases by downregulating the truncated neurokinin receptor-1 (NK1R-Tr) expression [114].
Signaling intrinsic to the cancer cell is also likely to contribute to the dormant state by expressing metastasis suppressor genes that contribute to dormancy without affecting the growth of cells in the originating primary tumor [156]. The tyrosine kinase receptor TIE2, which induces dormancy in hematopoietic stem cells, also induces cell cycle arrest in breast cancer cells through CDK inhibitors CDKN1A (p21Waf1) and CDKN1B (p27Kip1) in vitro, decreases osteolytic metastases and response to antimetabolites in mice, and is associated with delayed time to metastasis in breast cancer patients [157]. Expression of the metastasis suppressor genes KISS-1, metastasis suppressor Kangai-1 (KAI1), mitogen-activated protein kinase (MKK)4/7 and NM23 nucleoside diphosphate kinase 1 (Nm23-H1) by cancer cells also promotes tumor dormancy at the metastatic site [156]. Signaling through indoleamine 2,3-dioxygenase 1 (IDO1) through the mammalian target of rapamycin (mTOR) and control of nonderepressible-2 kinase has been linked to cellular quiescence [158]. IDO1, which is a heme-containing enzyme that mediates the rate-limiting step in the metabolism of l-tryptophan to kynurenine, has been explored as a potential immunotherapeutic target in oncology [159]. An inhibitor of this pathway has been found to have an acceptable toxicological spectrum in animal studies [159].
Other mechanisms of inducing reversible dormancy functions through epigenetic modifications, such as repressive histones [160] or the downregulation of suppressor gene promoter methylation enzymes [161], have been explored. These effects are analogous to evolutionary mechanisms that ensure the survival of organisms in environmentally disadvantageous circumstances [162]. They can also originate in the primary tumor, where epigenetic modifications in some of the cells enable them to enter dormancy in a distant microenvironment by cancer-associated fibroblasts (CAF) with altered p53 functions [163]. In tumor cells, the downregulation of DNA methyltransferase (DNMT)1 expression results in silencing a transcription network regulating the G1-S transition, including forkhead box (FOX)M1, FOXD, FOXL, early growth response (EGR)1/2/3, peroxisome proliferator-activated receptor (PPAR)γ, ETS Like-1 protein Elk-1 (ELK1) and Jun family members [64][162]. However, the dormancy-associated genes p53, DEC2, nuclear receptor subfamily 2 group F member (NR2F)1 and retinoic acid receptor (RAR)β, which are often silenced in proliferating cancer, are upregulated in dormant cells [64][162]. NR2F1 and RARβ together direct the removal of acetyl groups from histone H3 by histone deacetylases (HDACs) and are associated with dormant DTCs in patients [79][164][165]. In contrast, NR2F1 induces the methylation of H3 residues histone H3 (H3)K4, H3K9 and H3K27 and decreases the expression of growth-promoting SOX9 [162]. Histone H4 methylation is necessary for breast cancer dormancy in the lungs [166]. Epigenetics also affect dormancy and proliferation by governing the processing of coding mRNA alternative isoforms and non-coding RNAs, including micro-RNAs and long non-coding RNAs [167][168].
The overall effect of the metastatic microenvironment is to impose a reversible state of dormancy on the microscopic disseminated tumor-initiating cells. This effect is mediated through cancer cell interactions with structural, soluble, cellular and biophysical elements of the microenvironment that initiate signaling through a variety of receptors and sensors outlined above in order to change gene expression and phenotypic patterns to induce a dormant, cytotoxin-resistant state.
Table 1. Mechanisms of breast cancer dormancy in the bone marrow.
| Vehicle | Signaling | References | |
|---|---|---|---|
| Endosteal niche |
MSCs |
MERTK, AXL, TGFβR3, BMPR2, Alk5, NDRG1, ERK, p38, p21WAF1, p27Kip1, 15INK4b, PI3K, RhoA/GRAF, integrin α5β1, FGF-2, HSPG, fibronectin | [22][32][67][74][76][77][79][80][84][85][86][88][89][91][100][101][105][106][107][108][109][110][111][112][113][114][115][116][117][118][119][120][121][122] |
| Inhibition of oncogene signaling | [78] | ||
| Non-canonical Wnt5a signaling, SIAH2, repression of β-catenin, LIF, RA | [80][128][129] | ||
| Hypoxia, acidic pH | LIF, STAT3, TGFβ, BMP signaling | [73][125][133] | |
| Redox signaling | [126] | ||
| Exosomes | miR-23b, -126, 127, -148a, -3p -197, -222, -223 | [80][81][111][112][113][127][139] | |
| Fusion with and cannibalizing MSCs | SDF-1a, decreased CXCL12 | [112][114][130][132] | |
| Microenvironmental remodeling | [115] | ||
| Preosteoblasts, SNO cells | Notch, Jagged1 | [34][135][136] | |
| Vascular niche | Endothelial cells | TSP1 | [32] |
| DARC, KAI1, p21Waf1, downregulated TBX2 |
[140] | ||
| vWF, VCAM1, CXCL 1 and 2, BMP7, TGFβ-2, NFκB combined with ER in ER+ BC, NR2F1, ZFP281, PTEN | [8][77][78][79][141][142][143] |
||
| Prx1+ MSCs | CXCL12 | [144] | |
| Immune niche | CD4+ and CD8+ cells | INFγ | [32][152] |
| NK cells | DKK-1, inhibited canonical Wnt | [154] | |
| SDF-1/CXCR4 | Src, Akt, TRAIL, downregulated NK1R-Tr | [114][155] | |
| Cancer cell-intrinsic effects | TIE2 | p21Waf1, p27Kip1 | [156][157] |
| KAI1, MKK4/7, Nm23-H1 | [156] | ||
| IDO1 | mTOR | [158] | |
| Epigenetics | Repressive histones | altered p53 functions | [160][161][163] |
| downregulation of suppressor gene promoter methylation enzymes | |||
| downregulation of DNMT1 |
silencing of a transcription network FOXM1, FOXD, FOXL EGR1/2/3, PPARγ, ELK1, Jun family upregulating p53, DEC2, NR2F1, RARβ |
[64][162] |
|
| NR2F1, RARβ | removal of acetyl groups from histone H3, HDACs | [79][164][165] | |
| NR2F1 |
induced methylation of H3 residues H3K4, H3K9, H3K27, decreased expression of growth-promoting SOX9 | [162] |
|
| processing alternative coding mRNA isoforms, non-coding RNAs, miRNAs, lnRNAs | [167][168] |
References
- American Cancer Society Facts and Figures. 2021. Available online: https://www.cancer.org/research/cancer-facts-statistics/all-cancer-facts-figures/cancer-facts-figures-2021.html (accessed on 6 June 2022).
- Braun, S.; Pantel, K.; Muller, P.; Janni, W.; Hepp, F.; Kentenich, C.R.; Gastroph, S.; Wischnik, A.; Dimpfl, T.; Kindermann, G.; et al. Cytokeratin-positive cells in the bone marrow and survival of patients with stage I, II, or III breast cancer. N. Engl. J. Med. 2000, 342, 525–533.
- Hartkopf, A.D.; Brucker, S.Y.; Taran, F.A.; Harbeck, N.; von Au, A.; Naume, B.; Pierga, J.Y.; Hoffmann, O.; Beckmann, M.W.; Rydén, L.; et al. Disseminated tumour cells from the bone marrow of early breast cancer patients: Results from an international pooled analysis. Eur. J. Cancer 2021, 154, 128–137.
- Klein, C.A. Parallel progression of primary tumours and metastases. Nat. Rev. Cancer 2009, 9, 302–312.
- Huang, Z.; Wu, T.; Liu, A.Y.; Ouyang, G. Differentiation and transdifferentiation potentials of cancer stem cells. Oncotarget 2015, 6, 39550–39563.
- Braun, S.; Kentenich, C.; Janni, W.; Hepp, F.; de Waal, J.; Willegroth, F.; Sommer, H.; Pantel, K. Lack of an effect of adjuvant chemotherapy on the elimination of single dormant tumor cells in bone marrow of high risk breast cancer patients. J. Clin. Onc. 2000, 18, 80–86.
- Naumov, G.N.; Townson, J.L.; MacDonald, I.C.; Wilson, S.M.; Bramwell, V.H.; Groom, A.C.; Chambers, A.F. Ineffectiveness of doxorubicin treatment on solitary dormant mammary carcinoma cells or late-developing metastases. Breast Cancer Res. Treat. 2003, 82, 199–206.
- Carlson, P.; Dasgupta, A.; Grzelak, C.A.; Kim, J.; Barrett, A.; Coleman, I.M.; Shor, R.E.; Goddard, E.T.; Dai, J.; Schweitzer, E.M.; et al. Targeting the perivascular niche sensitizes disseminated tumour cells to chemotherapy. Nat. Cell Biol. 2019, 21, 238–250.
- Hughes, R.; Chen, X.; Cowley, N.; Ottewell, P.D.; Hawkins, R.J.; Hunter, K.D.; Hobbs, J.K.; Brown, N.J.; Holen, I. Osteoblast-derived paracrine and juxtacrine signals protect disseminated breast cancer cells from stress. Cancers 2021, 13, 1366.
- Brenner, S.; Whiting-Theobald, N.; Kawai, T.; Linton, G.F.; Rudikoff, A.G.; Choi, U.; Ryser, M.F.; Murphy, P.M.; Sechler, J.M.; Malech, H.L. CXCR4-transgene expression significantly improves marrow engraftment of cultured hematopoietic stem cells. Stem Cells 2004, 22, 1128–1133.
- Taichman, R.S.; Cooper, C.; Keller, E.T.; Pienta, K.J.; Taichman, N.S.; McCauley, L.K. Use of the stromal cell-derived factor-1/CXCR4 pathway in prostate cancer metastasis to bone. Cancer Res. 2002, 62, 1832–1837.
- Sun, S.; Guo, Z.; Xiao, X.; Liu, B.; Liu, X.; Tang, P.H.; Mao, N. Isolation of mouse marrow mesenchymal progenitors by a novel and reliable method. Stem Cells. 2003, 21, 527–535.
- Wang, J.; Loberg, R.; Taichman, R.S. The pivotal role of CXCL12 (SDF-1)/CXCR4 axis in bone metastasis. Cancer Metastasis Rev. 2006, 25, 573–587.
- Shiozawa, Y.; Havens, A.M.; Jung, Y.; Ziegler, A.M.; Pedersen, E.A.; Wang, J.; Wang, J.; Lu, G.; Roodman, G.D.; Loberg, R.D.; et al. Annexin II/annexin II receptor axis regulates adhesion, migration, homing, and growth of prostate cancer. J. Cell. Biochem. 2008, 105, 370–380.
- Jung, Y.; Wang, J.; Song, J.; Shiozawa, Y.; Wang, J.; Havens, A.; Wang, Z.; Sun, Y.-X.; Emerson, S.G.; Krebsbach, P.H.; et al. Annexin II expressed by osteoblasts and endothelial cells regulates stem cell adhesion, homing, and engraftment following transplantation. Blood 2007, 110, 82–90.
- Jung, Y.; Shiozawa, Y.; Wang, J.; Patel, L.R.; Havens, A.M.; Song, J.; Krebsbach, P.H.; Roodman, G.D.; Taichman, R.S. Annexin-2 is a regulator of stromal cell-derived factor-1/cxcl12 function in the hematopoietic stem cell endosteal niche. Exp. Hematol. 2011, 39, 151–166.e1.
- Jung, Y.; Wang, J.; Lee, E.; McGee, S.; Berry, J.E.; Yumoto, K.; Dai, J.; Keller, E.T.; Shiozawa, Y.; Taichman, R.S. Annexin 2-cxcl12 interactions regulate metastatic cell targeting and growth in the bone marrow. Mol. Cancer Res. MCR 2015, 13, 197–207.
- Chu, K.; Cheng, C.J.; Ye, X.; Lee, Y.C.; Zurita, A.J.; Chen, D.T.; Yu-Lee, L.Y.; Zhang, S.; Yeh, E.T.; Hu, M.C.; et al. Cadherin-11 promotes the metastasis of prostate cancer cells to bone. Mol. Cancer Res. 2008, 6, 1259–1267.
- Tamura, D.; Hiraga, T.; Myoui, A.; Yoshikawa, H.; Yoneda, T. Cadherin-11-mediated Interactions with Bone Marrow Stromal/Osteoblastic Cells Support Selective Colonization of Breast Cancer Cells in Bone. Int. J. Oncol. 2008, 33, 17–24.
- Kang, Y.; Siegel, P.M.; Shu, W.; Drobnjak, M.; Kakonen, S.M.; Cordon-Cardo, C.; Guise, T.A.; Massague, J. A multigenic program mediating breast cancer metastasis to bone. Cancer Cell 2003, 3, 537–549.
- Li, X.Q.; Lu, J.T.; Tan, C.C.; Wang, Q.S.; Feng, Y.M. RUNX2 promotes breast cancer bone metastasis by increasing integrin alpha5-mediated colonization. Cancer Lett. 2016, 380, 78–86.
- Shiozawa, Y.; Pedersen, E.A.; Patel, L.R.; Ziegler, A.M.; Havens, A.M.; Jung, Y.; Wang, J.; Zalucha, S.; Loberg, R.D.; Pienta, K.J.; et al. Gas6/axl axis regulates prostate cancer invasion, proliferation, and survival in the bone marrow niche. Neoplasia 2010, 12, 116–127.
- Wieder, R. Stromal Co-Cultivation for Modeling Breast Cancer Dormancy in the Bone Marrow. Cancers 2022, 14, 3344.
- Beerman, I.; Luis, T.C.; Singbrant, S.; Lo Celso, C.; Mendez-Ferrer, S. The evolving view of the hematopoietic stem cell niche. Exp. Hematol. 2017, 50, 22–26.
- Calvi, L.M.; Adams, G.B.; Weibrecht, K.W.; Weber, J.M.; Olson, D.P.; Knight, M.C.; Martin, R.P.; Schipani, E.; Divieti, P.; Bringhurst, F.R.; et al. Osteoblastic cells regulate the haematopoietic stem cell niche. Nature 2003, 425, 841–846.
- Kiel, M.J.; Yilmaz, O.H.; Iwashita, T.; Yilmaz, O.H.; Terhorst, C.; Morrison, S.J. SLAM family receptors distinguish hematopoietic stem and progenitor cells and reveal endothelial niches for stem cells. Cell 2005, 121, 1109–1121.
- Lo Celso, C.; Fleming, H.E.; Wu, J.W.; Zhao, C.X.; Miake-Lye, S.; Fujisaki, J.; Cote, D.; Rowe, D.W.; Lin, C.P.; Scadden, D.T. Live-animal tracking of individual haematopoietic stem/progenitor cells in their niche. Nature 2009, 457, 92–96.
- Ho, Y.H.; Mendez-Ferrer, S. Microenvironmental contributions to hematopoietic stem cell aging. Haematologica 2020, 105, 38–46.
- Kent, D.G.; Copley, M.R.; Benz, C.; Wohrer, S.; Dykstra, B.J.; Ma, E.; Cheyne, J.; Zhao, Y.; Bowie, M.B.; Zhao, Y.; et al. Prospective isolation and molecular characterization of hematopoietic stem cells with durable self-renewal potential. Blood 2009, 113, 6342–6350.
- Grover, A.; Sanjuan-Pla, A.; Thongjuea, S.; Carrelha, J.; Giustacchini, A.; Gambardella, A.; Macaulay, I.; Mancini, E.; Luis, T.C.; Mead, A.; et al. Single-cell RNA sequencing reveals molecular and functional platelet bias of aged haematopoietic stem cells. Nat. Commun. 2016, 7, 11075.
- Carrelha, J.; Meng, Y.; Kettyle, L.M.; Luis, T.C.; Norfo, R.; Alcolea, V.; Boukarabila, H.; Grasso, F.; Gambardella, A.; Grover, A.; et al. Hierarchically related lineage-restricted fates of multipotent haematopoietic stem cells. Nature 2018, 554, 106–111.
- Sosa, M.S.; Bragado, P.; Aguirre-Ghiso, J.A. Mechanisms of disseminated cancer cell dormancy: An awakening field. Nat. Rev. Cancer 2014, 14, 611–622.
- Li, Z.; Li, L. Understanding hematopoietic stem-cell microenvironments. Trends Biochem. Sci. 2006, 31, 589–595.
- Morrison, S.J.; Scadden, D.T. The bone marrow niche for haematopoietic stem cells. Nature 2014, 505, 327–334.
- Chitteti, B.R.; Cheng, Y.H.; Kacena, M.A.; Srour, E.F. Hierarchical organization of osteoblasts reveals the significant role of CD166 in hematopoietic stem cell maintenance and function. Bone 2013, 54, 58–67.
- Hooker, R.A.; Chitteti, B.R.; Egan, P.H.; Cheng, Y.H.; Himes, E.R.; Meijome, T.; Srour, E.F.; Fuchs, R.K.; Kacena, M.A. Activated leukocyte cell adhesion molecule (ALCAM or CD166) modulates bone phenotype and hematopoiesis. J. Musculoskelet. Neuronal Interact. 2015, 15, 83–94.
- Panaroni, C.; Fulzele, K.; Saini, V.; Chubb, R.; Pajevic, P.D.; Wu, J.Y. PTH Signaling in Osteoprogenitors Is Essential for B-Lymphocyte Differentiation and Mobilization. J. Bone Miner. Res. 2015, 30, 2273–2286.
- He, Q.; Scott Swindle, C.; Wan, C.; Flynn, R.J.; Oster, R.A.; Chen, D.; Zhang, F.; Shu, Y.; Klug, C.A. Enhanced Hematopoietic Stem Cell Self-Renewal-Promoting Ability of Clonal Primary Mesenchymal Stromal/Stem cells Versus Their Osteogenic Progeny. Stem Cells 2017, 35, 473–484.
- Wang, H.; Tian, L.; Liu, J.; Goldstein, A.; Bado, I.; Zhang, W.; Arenkiel, B.R.; Li, Z.; Yang, M.; Du, S.; et al. The Osteogenic Niche Is a Calcium Reservoir of Bone Micrometastases and Confers Unexpected Therapeutic Vulnerability. Cancer Cell 2018, 34, 823–839.e7.
- Khurana, S.; Schouteden, S.; Manesia, J.K.; Santamaria-Martinez, A.; Huelsken, J.; Lacy-Hulbert, A.; Verfaillie, C.M. Outside-in integrin signalling regulates haematopoietic stem cell function via Periostin-Itgav axis. Nat. Commun. 2016, 7, 13500.
- Kawamori, Y.; Katayama, Y.; Asada, N.; Minagawa, K.; Sato, M.; Okamura, A.; Shimoyama, M.; Nakagawa, K.; Okano, T.; Tanimoto, M.; et al. Role for vitamin D receptor in the neuronal control of the hematopoietic stem cell niche. Blood 2010, 116, 5528–5535.
- Yeh, S.A.; Hou, J.; Wu, J.W.; Yu, S.; Zhang, Y.; Belfield, K.D.; Camargo, F.D.; Lin, C.P. Quantification of bone marrow interstitial pH and calcium concentration by intravital ratiometric imaging. Nat. Commun. 2022, 13, 393.
- Maryanovich, M.; Zahalka, A.H.; Pierce, H.; Pinho, S.; Nakahara, F.; Asada, N.; Wei, Q.; Wang, X.; Ciero, P.; Xu, J.; et al. Adrenergic nerve degeneration in bone marrow drives aging of the hematopoietic stem cell niche. Nat. Med. 2018, 24, 782–791.
- Mendez-Ferrer, S.; Michurina, T.V.; Ferraro, F.; Mazloom, A.R.; Macarthur, B.D.; Lira, S.A.; Scadden, D.T.; Ma’ayan, A.; Enikolopov, G.N.; Frenette, P.S. Mesenchymal and haematopoietic stem cells form a unique bone marrow niche. Nature 2010, 466, 829–834.
- Kim, M.; Kim, C.; Choi, Y.S.; Kim, M.; Park, C.; Suh, Y. Age-related alterations in mesenchymal stem cells related to shift in differentiation from osteogenic to adipogenic potential: Implication to age-associated bone diseases and defects. Mech. Ageing Dev. 2012, 133, 215–225.
- Lai, P.; Song, Q.; Yang, C.; Li, Z.; Liu, S.; Liu, B.; Li, M.; Deng, H.; Cai, D.; Jin, D.; et al. Loss of Rictor with aging in osteoblasts promotes age-related bone loss. Cell Death Dis. 2016, 7, e2408.
- Sui, B.; Hu, C.; Liao, L.; Chen, Y.; Zhang, X.; Fu, X.; Zheng, C.; Li, M.; Wu, L.; Zhao, X.; et al. Mesenchymal progenitors in osteopenias of diverse pathologies: Differential characteristics in the common shift from osteoblastogenesis to adipogenesis. Sci. Rep. 2016, 6, 30186.
- Nehlin, J.O.; Jafari, A.; Tencerova, M.; Kassem, M. Aging and lineage allocation changes of bone marrow skeletal (stromal) stem cells. Bone 2019, 123, 265–273.
- Tikhonova, A.N.; Dolgalev, I.; Hu, H.; Sivaraj, K.K.; Hoxha, E.; Cuesta-Dominguez, A.; Pinho, S.; Akhmetzyanova, I.; Gao, J.; Witkowski, M.; et al. The bone marrow microenvironment at single-cell resolution. Nature 2019, 569, 222–228.
- Trotter, T.N.; Gibson, J.T.; Sherpa, T.L.; Gowda, P.S.; Peker, D.; Yang, Y. Adipocyte-Lineage Cells Support Growth and Dissemination of Multiple Myeloma in Bone. Am. J. Pathol. 2016, 186, 3054–3063.
- Raaijmakers, M.H.; Mukherjee, S.; Guo, S.; Zhang, S.; Kobayashi, T.; Schoonmaker, J.A.; Ebert, B.L.; Al-Shahrour, F.; Hasserjian, R.P.; Scadden, E.O.; et al. Bone progenitor dysfunction induces myelodysplasia and secondary leukaemia. Nature 2010, 464, 852–857.
- Kode, A.; Manavalan, J.S.; Mosialou, I.; Bhagat, G.; Rathinam, C.V.; Luo, N.; Khiabanian, H.; Lee, A.; Murty, V.V.; Friedman, R.; et al. Leukaemogenesis induced by an activating beta-catenin mutation in osteoblasts. Nature 2014, 506, 240–244.
- Bowers, M.; Zhang, B.; Ho, Y.; Agarwal, P.; Chen, C.C.; Bhatia, R. Osteoblast ablation reduces normal long-term hematopoietic stem cell self-renewal but accelerates leukemia development. Blood 2015, 125, 2678–2688.
- Krevvata, M.; Silva, B.C.; Manavalan, J.S.; Galan-Diez, M.; Kode, A.; Matthews, B.G.; Park, D.; Zhang, C.A.; Galili, N.; Nickolas, T.L.; et al. Inhibition of leukemia cell engraftment and disease progression in mice by osteoblasts. Blood 2014, 124, 2834–2846.
- Gabbianelli, M.; Sargiacomo, M.; Pelosi, E.; Testa, U.; Isacchi, G.; Peschle, C. “Pure” human hematopoietic progenitors: Permissive action of basic fibroblast growth factor. Science 1990, 249, 1561–1564.
- Yeoh, J.S.; van Os, R.; Weersing, E.; Ausema, A.; Dontje, B.; Vellenga, E.; de Haan, G. Fibroblast growth factor-1 and -2 preserve long-term repopulating ability of hematopoietic stem cells in serum-free cultures. Stem Cells 2006, 24, 1564–1572.
- Itkin, T.; Kaufmann, K.B.; Gur-Cohen, S.; Ludin, A.; Lapidot, T. Fibroblast growth factor signaling promotes physiological bone remodeling and stem cell self-renewal. Curr. Opin. Hematol. 2013, 20, 237–244.
- Abdallah, B.M.; Alzahrani, A.M.; Abdel-Moneim, A.M.; Ditzel, N.; Kassem, M. A simple and reliable protocol for long-term culture of murine bone marrow stromal (mesenchymal) stem cells that retained their in vitro and in vivo stemness in long-term culture. Biol. Proced. Online 2019, 21, 3.
- Gabrilove, J.L.; Wong, G.; Bollenbacher, E.; White, K.; Kojima, S.; Wilson, E.L. Basic fibroblast growth factor counteracts the suppressive effect of transforming growth factor-beta 1 on human myeloid progenitor cells. Blood 1993, 81, 909–915.
- Gabrilove, J.L.; White, K.; Rahman, Z.; Wilson, E.L. Stem cell factor and basic fibroblast growth factor are synergistic in augmenting committed myeloid progenitor cell growth. Blood 1994, 83, 907–910.
- Buono, M.; Visigalli, I.; Bergamasco, R.; Biffi, A.; Cosma, M.P. Sulfatase modifying factor 1-mediated fibroblast growth factor signaling primes hematopoietic multilineage development. J. Exp. Med. 2010, 207, 1647–1660.
- Wilson, E.L.; Rifkin, D.B.; Kelly, F.; Hannocks, M.J.; Gabrilove, J.L. Basic fibroblast growth factor stimulates myelopoiesis in long-term human bone marrow cultures. Blood 1991, 77, 954–960.
- Weidenfeld, K.; Schif-Zuck, S.; Abu-Tayeh, H.; Kang, K.; Kessler, O.; Weissmann, M.; Neufeld, G.; Barkan, D. Dormant tumor cells expressing LOXL2 acquire a stem-like phenotype mediating their transition to proliferative growth. Oncotarget 2016, 7, 71362–71377.
- Adam, A.P.; George, A.; Schewe, D.; Bragado, P.; Iglesias, B.V.; Ranganathan, A.C.; Kourtidis, A.; Conklin, D.S.; Aguirre-Ghiso, J.A. Computational identification of a p38sapk-regulated transcription factor network required for tumor cell quiescence. Cancer Res. 2009, 69, 5664–5672.
- Kim, R.S.; Avivar-Valderas, A.; Estrada, Y.; Bragado, P.; Sosa, M.S.; Aguirre-Ghiso, J.A.; Segall, J.E. Dormancy signatures and metastasis in estrogen receptor positive and negative breast cancer. PLoS ONE 2012, 7, e35569.
- Cheung, T.H.; Rando, T.A. Molecular regulation of stem cell quiescence. Nature Rev. Mo. Cell Biol. 2013, 14, 329–340.
- Giancotti, F.G. Mechanisms governing metastatic dormancy and reactivation. Cell 2013, 155, 750–764.
- Wieder, R. Fibroblasts as Turned Agents in Cancer Progression. Cancers 2023, 15, 2014.
- Dong, Q.; Liu, X.; Cheng, K.; Sheng, J.; Kong, J.; Liu, T. Pre-metastatic niche formation in different organs induced by tumor extracellular vesicles. Front. Cell Dev. Biol. 2021, 9, 733627.
- Li, R.; Wen, A.; Lin, J. Pro-inflammatory cytokines in the formation of the pre-metastatic niche. Cancers 2020, 12, 3752.
- Sanmartin, M.C.; Borzone, F.R.; Giorello, M.B.; Pacienza, N.; Yannarelli, G.; Chasseing, N.A. Bone marrow/bone pre-metastatic niche for breast cancer cells colonization: The role of mesenchymal stromal cells. Crit. Rev. Oncol. Hematol. 2021, 164, 103416.
- Bakhshandeh, S.; Werner, C.; Fratzl, P.; Cipitria, A. Microenvironment-mediated cancer dormancy: Insights from metastability theory. Proc. Natl. Acad. Sci. USA 2022, 119, 46118.
- Johnson, R.W.; Sowder, M.E.; Giaccia, A.J. Hypoxia and Bone Metastatic Disease. Curr. Osteoporos. Rep. 2017, 15, 231–238.
- Kaplan, R.N.; Riba, R.D.; Zacharoulis, S.; Bramley, A.H.; Vincent, L.; Costa, C.; MacDonald, D.D.; Jin, D.K.; Shido, K.; Kerns, S.A.; et al. VEGFR1-positive haematopoietic bone marrow progenitors initiate the pre-metastatic niche. Nature 2005, 438, 820–827.
- Fang, C.; Kang, Y. Cellular Plasticity in Bone Metastasis. Bone 2020, 158, 115693.
- Kobayashi, A.; Okuda, H.; Xing, F.; Pandey, P.R.; Watabe, M.; Hirota, S.; Pai, S.K.; Liu, W.; Fukuda, K.; Chambers, C.; et al. Bone morphogenetic protein 7 in dormancy and metastasis of prostate cancer stem-like cells in bone. J. Exp. Med. 2011, 208, 2641–2655.
- Bragado, P.; Estrada, Y.; Parikh, F.; Krause, S.; Capobianco, C.; Farina, H.G.; Schewe, D.M.; Aguirre-Ghiso, J.A. TGF-beta2 dictates disseminated tumour cell fate in target organs through TGF-beta-RIII and p38alpha/beta signalling. Nat. Cell Biol. 2013, 15, 1351–1361.
- White, A.C.; Khuu, J.K.; Dang, C.Y.; Hu, J.; Tran, K.V.; Liu, A.; Gomez, S.; Zhang, Z.; Yi, R.; Scumpia, P.; et al. Stem cell quiescence acts as a tumour suppressor in squamous tumours. Nat. Cell Biol. 2014, 16, 99–107.
- Sosa, M.S.; Parikh, F.; Maia, A.G.; Estrada, Y.; Bosch, A.; Bragado, P.; Ekpin, E.; George, A.; Zheng, Y.; Lam, H.M.; et al. NR2F1 controls tumour cell dormancy via SOX9- and RARbeta-driven quiescence programmes. Nat. Commun. 2015, 6, 6170.
- Risson, E.; Nobre, A.R.; Maguer-Satta, V.; Aguirre-Ghiso, J.A. The current paradigm and challenges ahead for the dormancy of disseminated tumor cells. Nat. Cancer 2020, 1, 672–680.
- Nobre, A.R.; Risson, E.; Singh, D.K.; di Martino, J.; Cheung, J.F.; Wang, J.; Johnson, J.; Russnes, H.G.; Bravo-Cordero, J.J.; Birbrair, A.; et al. Bone Marrow NG2+/Nestin+ mesenchymal stem cells drive DTC dormancy via TGFβ2. Nat. Cancer 2021, 2, 327–339.
- Gao, X.; Zhang, M.; Tang, Y.; Liang, X. Cancer cell dormancy: Mechanisms and implications of cancer recurrence and metastasis. Onco Targets Ther. 2017, 10, 5219–5228.
- Singh, A.; Veeriah, V.; Xi, P.; Labella, R.; Chen, J.; Romeo, S.G.; Ramasamy, S.K.; Kusumbe, A.P. Angiocrine signals regulate quiescence and therapy resistance in bone metastasis. JCI Insight 2019, 4, 125679.
- Korah, R.; Boots, M.; Wieder, R. Integrin α5β1 promotes survival of growth-arrested breast cancer cells: An in vitro paradigm for breast cancer dormancy in bone marrow. Cancer Res. 2004, 64, 4514–4522.
- Brunner, G.; Nguyen, H.; Gabrilove, J.; Rifkin, D.B.; Wilson, E.L. Basic fibroblast growth factor expression in human bone marrow and peripheral blood cells. Blood 1993, 81, 631–638.
- Hassan, N.; Greve, B.; Espinoza-Sanchez, N.A.; Gotte, M. Cell-surface heparan sulfate proteoglycans as multifunctional integrators of signaling in cancer. Cell. Signal. 2021, 77, 109822.
- Guereno, M.; Delgado Pastore, M.; Lugones, A.C.; Cercato, M.; Todaro, L.; Urtreger, A.; Peters, M.G. Glypican-3 (GPC3) inhibits metastasis development promoting dormancy in breast cancer cells by p38 MAPK pathway activation. Eur. J. Cell Biol. 2020, 99, 151096.
- Nilsson, S.K.; Debatis, M.E.; Dooner, M.S.; Madri, J.A.; Quesenberry, P.J.; Becker, P.S. Immunofluorescence characterization of key extracellular matrix proteins in murine bone marrow in situ. J. Histochem. Cytochem. 1998, 46, 371–377.
- Barney, L.E.; Hall, C.L.; Schwartz, A.D.; Parks, A.N.; Sparages, C.; Galarza, S.; Platt, M.O.; Mercurio, A.M.; Peyton, S.R. Tumor cell-organized fibronectin maintenance of a dormant breast cancer population. Sci. Adv. 2020, 6, eaaz4157.
- Barkan, D.; Kleinman, H.; Simmons, J.L.; Asmussen, H.; Kamaraju, A.K.; Hoenorhoff, M.J.; Liu, Z.Y.; Costes, S.V.; Cho, E.H.; Lockett, S.; et al. Inhibition of metastatic outgrowth from single dormant tumor cells by targeting the cytoskeleton. Cancer Res. 2008, 68, 6241–6250.
- Giancotti, F.G.; Ruoslahti, E. Elevated levels of the alpha 5 beta 1 fibronectin receptor suppress the transformed phenotype of Chinese hamster ovary cells. Cell 1990, 60, 849–859.
- Montagner, M.; Dupont, S. Mechanical Forces as Determinants of Disseminated Metastatic Cell Fate. Cells 2020, 9, 250.
- Klein, C.A.; Blankenstein, T.J.; Schmidt-Kittler, O.; Petronio, M.; Polzer, B.; Stoecklein, N.H.; Riethmuller, G. Genetic heterogeneity of single disseminated tumour cells in minimal residual cancer. Lancet 2002, 360, 683–689.
- Magbanua, M.J.M.; Rugo, H.S.; Hauranieh, L.; Roy, R.; Scott, J.H.; Lee, J.C.; Hsiao, F.; Sosa, E.V.; Van’t Veer, L.; Esserman, L.J.; et al. Genomic and expression profiling reveal molecular heterogeneity of disseminated tumor cells in bone marrow of early breast cancer. NPJ Breast Cancer 2018, 4, 31.
- Balic, M.; Lin, H.; Young, L.; Hawes, D.; Giuliano, A.; McNamara, G.; Datar, R.H.; Cote, R.J. Most Early Disseminated Cancer Cells Detected in Bone Marrow of Breast Cancer Patients Have a Putative Breast Cancer Stem Cell Phenotype. Clin. Cancer Res. 2006, 12, 5615–5621.
- Lawson, D.A.; Bhakta, N.R.; Kessenbrock, K.; Prummel, K.D.; Yu, Y.; Takai, K.; Zhou, A.; Eyob, H.; Balakrishnan, S.; Wang, C.Y.; et al. Single-cell analysis reveals a stem-cell program in human metastatic breast cancer cells. Nature 2015, 526, 131–135.
- Sistigu, A.; Musella, M.; Galassi, C.; Vitale, I.; De Maria, R. Tuning Cancer Fate: Tumor Microenvironment’s Role in Cancer Stem Cell Quiescence and Reawakening. Front. Immunol. 2020, 11, 2166.
- Huang, Y.; Hamana, T.; Liu, J.; Wang, C.; An, L.; You, P.; Chang, J.Y.; Xu, J.; Jin, C.; Zhang, Z.; et al. Type 2 Fibroblast Growth Factor Receptor Signaling Preserves Stemness and Prevents Differentiation of Prostate Stem Cells from the Basal Compartment. J. Biol. Chem. 2015, 290, 17753–17761.
- Quan, M.Y.; Guo, Q.; Liu, J.; Yang, R.; Bai, J.; Wang, W.; Cai, Y.; Han, R.; Lv, Y.Q.; Ding, L.; et al. An FGFR/AKT/SOX2 Signaling Axis Controls Pancreatic Cancer Stemness. Front. Cell Dev. Biol. 2020, 8, 287.
- Goel, S.; DeCristo, M.J.; Watt, A.C.; BrinJones, H.; Sceneay, J.; Li, B.B.; Khan, N.; Ubellacker, J.M.; Xie, S.; Metzger-Filho, O.; et al. CDK4/6 inhibition triggers anti-tumour immunity. Nature 2017, 548, 471–475.
- Watt, A.C.; Cejas, P.; DeCristo, M.J.; Metzger-Filho, O.; Lam, E.Y.N.; Qiu, X.; BrinJones, H.; Kesten, N.; Coulson, R.; Font-Tello, A.; et al. CDK4/6 inhibition reprograms the breast cancer enhancer landscape by stimulating AP-1 transcriptional activity. Nat. Cancer 2021, 2, 34–48.
- Yoon, K.A.; Son, Y.; Choi, Y.J.; Kim, J.H.; Cho, J.Y. Fibroblast growth factor 2 supports osteoblastic niche cells during hematopoietic homeostasis recovery after bone marrow suppression. Cell Commun. Signal. 2017, 15, 25.
- Bae, S.H.; Ryu, H.; Rhee, K.J.; Oh, J.E.; Baik, S.K.; Shim, K.Y.; Kong, J.H.; Hyun, S.Y.; Pack, H.S.; Im, C.; et al. L-ascorbic acid 2-phosphate and fibroblast growth factor-2 treatment maintains differentiation potential in bone marrow-derived mesenchymal stem cells through expression of hepatocyte growth factor. Growth Factors 2015, 33, 71–78.
- Ito, T.; Sawada, R.; Fujiwara, Y.; Tsuchiya, T. FGF-2 increases osteogenic and chondrogenic differentiation potentials of human mesenchymal stem cells by inactivation of TGF-beta signaling. Cytotechnology 2008, 56, 1–7.
- Najmi, S.; Korah, R.; Chandra, R.; Abdellatif, M.; Wieder, R. Flavopiridol blocks integrin-mediated survival in dormant breast cancer cells. Clin. Cancer Res. 2005, 11, 2038–2046.
- Barrios, J.; Wieder, R. Dual FGF-2 and intergrin α5β1 signaling mediate GRAF-induced RhoA inactivation in a model of breast cancer dormancy. Cancer Microenviron. 2009, 2, 33–47.
- Tivari, S.; Lu, H.; Dasgpta, T.; De Lorenzo, M.S.; Wieder, R. Reawakening of dormant estrogen-dependent human breast cancer cells by bone marrow stroma secretory senescence. Cell Commun. Signal. 2018, 16, 48.
- Fenig, E.; Wieder, R.; Paglin, S.; Wang, H.; Persaud, R.; Haimovitz-Friedman, A.; Fuks, Z.; Yahalom, J. Basic fibroblast growth factor confers growth inhibition and Mitogen-activated Protein Kinase activation in human breast cancer cells. Clin. Cancer Res. 1997, 3, 135–142.
- Wang, H.; Rubin, M.; Fenig, E.; DeBlasio, T.; Mendelsohn, J.; Yahalom, J.; Wieder, R. Basic FGF causes growth arrest in MCF-7 human breast cancer cells while inducing both mitogenic and inhibitory G1 events. Cancer Res. 1997, 57, 1750–1757.
- Fenig, E.; Kanfi, Y.; Wang, Q.; Beery, E.; Livnat, T.; Wasserman, L.; Lilling, G.; Yahalom, J.; Wieder, R.; Nordenberg, J. Role of transforming growth factor beta in the growth inhibition of human breast cancer cells by basic fibroblast growth factor. Breast Cancer Res. Treat. 2001, 70, 27–37.
- Ono, M.; Kosala, N.; Tominaga, N.; Yoshioka, Y.; Takeshita, F.; Takahashi, R.U.; Yoshida, M.; Tsuda, H.; Tamura, K.; Ochiya, T. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci. Signal. 2014, 7, ra63.
- Lim, P.K.; Bliss, S.A.; Patel, S.A.; Taborga, M.; Dave, M.A.; Gregory, L.A.; Greco, S.J.; Bryan, M.; Patel, P.S.; Rameshwar, P. Gap junction-mediated import of microRNA from bone marrow stromal cells can elicit cell cycle quiescence in breast cancer cells. Cancer Res. 2011, 71, 1550–1560.
- Greco, S.J.; Rameshwar, P. Analysis of the transfer of circulating microRNA between cells mediated by gap junction. Methods Mol. Biol. 2013, 1024, 87–96.
- Zhou, Y.; Zuo, D.; Wang, M.; Zhang, Y.; Yu, M.; Yang, J.; Yao, Z. Effect of truncated neurokinin-1 receptor expression changes on the interaction between human breast cancer and bone marrow-derived mesenchymal stem cells. Genes Cells 2014, 19, 676–691.
- Di Martino, J.S.; Akhter, T.; Bravo-Cordero, J.J. Remodeling the ECM: Implications for metastasis and tumor dormancy. Cancers 2021, 13, 4916.
- Kim, S.; Dubrovska, A.; Salamone, R.J.; Walker, J.R.; Grandinetti, K.B.; Bonamy, G.M.; Orth, A.P.; Elliott, J.; Porta, D.G.; Garcia-Echeverria, C.; et al. FGFR2 promotes breast tumorigenicity through maintenance of breast tumor-initiating cells. PLoS ONE 2013, 8, e51671.
- Coller, H.A.; Sang, L.; Roberts, J.M. A new description of cellular quiescence. PLoS Biol. 2006, 4, e83.
- Shiozawa, Y.; Pedersen, E.A.; Havens, A.M.; Jung, Y.; Mishra, A.; Joseph, J.; Kim, J.K.; Patel, L.R.; Ying, C.; Ziegler, A.M.; et al. Human prostate cancer metastases target the hematopoietic stem cell niche to establish footholds in mouse bone marrow. J. Clin. Investig. 2011, 121, 1298–1312.
- Yumoto, K.; Eber, M.R.; Wang, J.; Cackowski, F.C.; Decker, A.M.; Lee, E.; Nobre, A.R.; Aguirre-Ghiso, J.A.; Jung, Y.; Taichman, R.S. Axl is required for TGF-β2-induced dormancy of prostate cancer cells in the bone marrow. Sci. Rep. 2016, 6, 36520.
- Gawrzak, S.; Rinaldi, L.; Gregorio, S.; Arenas, E.J.; Salvador, F.; Urosevic, J.; Figueras-Puig, C.; Rojo, F.; Del Barco Barrantes, I.; Cejalvo, J.M.; et al. MSK1 regulates luminal cell differentiation and metastatic dormancy in ER+ breast cancer. Nat. Cell Biol. 2018, 20, 211–221.
- Marlow, R.; Honeth, G.; Lombardi, S.; Cariati, M.; Hessey, S.; Pipili, A.; Mariotti, V.; Buchupalli, B.; Foster, K.; Bonnet, D.; et al. A novel model of dormancy for bone metastatic breast cancer cells. Cancer Res. 2013, 73, 6886–6899.
- Cackowski, F.C.; Eber, M.R.; Rhee, J.; Decker, A.M.; Yumoto, K.; Berry, J.E.; Lee, E.; Shiozawa, Y.; Jung, Y.; Aguirre-Ghiso, J.A.; et al. Mer Tyrosine Kinase Regulates Disseminated Prostate Cancer Cellular Dormancy. J. Cell Biochem. 2017, 118, 891–902.
- Spencer, J.A.; Ferraro, F.; Roussakis, E.; Klein, A.; Wu, J.; Runnels, J.M.; Zaher, W.; Mortensen, L.J.; Alt, C.; Turcotte, R.; et al. Direct measurement of local oxygen concentration in the bone marrow of live animals. Nature 2014, 508, 269–273.
- Johnson, R.W.; Finger, E.C.; Olcina, M.M.; Vilalta, M.; Aguilera, T.; Miao, Y.; Merkel, A.R.; Johnson, J.R.; Sterling, J.A.; Wu, J.Y.; et al. Induction of LIFR confers a dormancy phenotype in breast cancer cells disseminated to the bone marrow. Nat. Cell Biol. 2016, 18, 1078–1089.
- Fluegen, G.; Avivar-Valderas, A.; Wang, Y.; Padgen, M.R.; Williams, J.K.; Nobre, A.R.; Calvo, V.; Cheung, J.F.; Bravo-Cordero, J.J.; Entenberg, D.; et al. Phenotypic heterogeneity of disseminated tumour cells is preset by primary tumour hypoxic microenvironments. Nat. Cell Biol. 2017, 19, 120–132.
- Qin, S.; Li, B.; Ming, H.; Nice, E.C.; Zou, B.; Huang, C. Harnessing redox signaling to overcome therapeutic-resistant cancer dormancy. Biochim. Et Biophys. Acta Rev. Cancer 2022, 1877, 188749.
- Bliss, S.A.; Sinha, G.; Sandiford, O.A.; Williams, L.M.; Engelberth, D.J.; Guiro, K.; Isenalumhe, L.L.; Greco, S.J.; Ayer, S.; Bryan, M.; et al. Mesenchymal Stem Cell-Derived Exosomes Stimulate Cycling Quiescence and Early Breast Cancer Dormancy in Bone Marrow. Cancer Res. 2016, 76, 5832–5844.
- Ren, D.; Dai, Y.; Yang, Q.; Zhang, X.; Guo, W.; Ye, L.; Huang, S.; Chen, X.; Lai, Y.; Du, H.; et al. Wnt5a induces and maintains prostate cancer cells dormancy in bone. J. Exp. Med. 2019, 216, 428–449.
- Baksh, D.; Tuan, R.S. Canonical and non-canonical Wnts differentially affect the development potential of primary isolate of human bone marrow mesenchymal stem cells. J. Cell. Physiol. 2007, 212, 817–826.
- Melzer, C.; Ohe, J.v.d.; Luo, T.; Hass, R. Spontaneous Fusion of MSC with Breast Cancer Cells Can Generate Tumor Dormancy. Int. J. Mol. Sci. 2021, 22, 5930.
- Melzer, C.; Ohe, J.v.d.; Hass, R. Altered Tumor Plasticity after Different Cancer Cell Fusions with MSC. Int. J. Mol. Sci. 2020, 21, 8347.
- Bartosh, T.J.; Ullah, M.; Zeitouni, S.; Beaver, J.; Prockop, D.J. Cancer cells enter dormancy after cannibalizing mesenchymal stem/stromal cells (MSCs). Proc. Natl. Acad. Sci. USA 2016, 113, E6447–E6456.
- Bui, A.T.; Laurent, F.; Havard, M.; Dautry, F.; Tchénio, T. SMAD signaling and redox imbalance cooperate to induce prostate cancer cell dormancy. Cell Cycle 2015, 14, 1218–1231.
- Colmone, A.; Amorim, M.; Pontier, A.L.; Wang, S.; Jablonski, E.; Sipkins, D.A. Leukemic cells create bone marrow niches that disrupt the behavior of normal hematopoietic progenitor cells. Science 2008, 322, 1861–1865.
- Capulli, M.; Hristova, D.; Valbret, Z.; Carys, K.; Arjan, R.; Maurizi, A.; Masedu, F.; Cappariello, A.; Rucci, N.; Teti, A. Notch2 pathway mediates breast cancer cellular dormancy and mobilisation in bone and contributes to haematopoietic stem cell mimicry. Br. J. Cancer 2019, 121, 157–171.
- Zheng, H.; Bae, Y.; Kasimir-Bauer, S.; Tang, R.; Chen, J.; Ren, G.; Yuan, M.; Esposito, M.; Li, W.; Wei, Y.; et al. Therapeutic Antibody Targeting Tumor- and Osteoblastic Niche-Derived Jagged1 Sensitizes Bone Metastasis to Chemotherapy. Cancer Cell 2017, 32, 731–747.
- Janghorban, M.; Yang, Y.; Zhao, N.; Hamor, C.; Nguyen, T.M.; Zhang, X.H.; Rosen, J.M. Single-cell analysis unveils the role of the tumor immune microenvironment and notch signaling in dormant minimal residual disease. Cancer Res. 2022, 82, 885–899.
- Kolb, A.D.; Shupp, A.B.; Mukhopadhyay, D.; Marini, F.C.; Bussard, K.M. Osteoblasts are “educated” by crosstalk with metastatic breast cancer cells in the bone tumor microenvironment. Breast Cancer Res. 2019, 21, 31.
- Shupp, A.B.; Neupane, M.; Agostini, L.C.; Ning, G.; Brody, J.R.; Bussard, K.M. Stromal-derived extracellular vesicles suppress proliferation of bone metastatic cancer cells mediated by ERK2. Mol. Cancer Res. MCR 2021, 19, 1763–1777.
- Bandyopadhyay, S.; Zhan, R.; Chaudhuri, A.; Watabe, M.; Pai, S.K.; Hirota, S.; Hosobe, S.; Tsukada, T.; Miura, K.; Takano, Y.; et al. Interaction of KAI1 on tumor cells with DARC on vascular endothelium leads to metastasis suppression. Nat. Med. 2006, 12, 933–938.
- Acharyya, S.; Oskarsson, T.; Vanharanta, S.; Malladi, S.; Kim, J.; Morris, P.G.; Manova-Todorova, K.; Leversha, M.; Hogg, N.; Seshan, V.E.; et al. A CXCL1 paracrine network links cancer chemoresistance and metastasis. Cell 2012, 150, 165–178.
- El-Shennawy, L.; Dubrovskyi, O.; Kastrati, I.; Danes, J.M.; Zhang, Y.; Whiteley, H.E.; Creighton, C.J.; Frasor, J. Coactivation of estrogen receptor and iKKbeta induces a dormant metastatic phenotype in ER-positive breast cancer. Cancer Res. 2018, 78, 974–984.
- Nobre, A.R.; Dalla, E.; Yang, J.; Huang, X.; Wullkopf, L.; Risson, E.; Razghandi, P.; Anton, M.L.; Zheng, W.; Seoane, J.A.; et al. ZFP281 drives a mesenchymal-like dormancy program in early disseminated breast cancer cells that prevents metastatic outgrowth in the lung. Nat. Cancer 2022, 3, 1165–1180.
- Agarwal, P.; Isringhausen, S.; Li, H.; Paterson, A.J.; He, J.; Gomariz, A.; Nagasawa, T.; Nombela-Arrieta, C.; Bhatia, R. Mesenchymal niche-specific expression of Cxcl12 controls quiescence of treatment-resistant leukemia stem cells. Cell Stem Cell 2019, 24, 769–784.
- Ghajar, C.M.; Peinado, H.; Mori, H.; Matei, I.R.; Evason, K.J.; Brazier, H.; Almeida, D.; Koller, A.; Hajjar, K.A.; Stainier, D.Y.; et al. The perivascular niche regulates breast tumour dormancy. Nat. Cell Biol. 2013, 15, 807–817.
- Yan, G.N.; Yang, L.; Lv, Y.F.; Shi, Y.; Shen, L.L.; Yao, X.H.; Guo, Q.N.; Zhang, P.; Cui, Y.H.; Zhang, X.; et al. Endothelial cells promote stem-like phenotype of glioma cells through activating the Hedgehog pathway. J. Pathol. 2014, 234, 11–22.
- Kurebayashi, J.; Koike, Y.; Ohta, Y.; Saitoh, W.; Yamashita, T.; Kanomata, N.; Moriya, T. Anti-cancer stem cell activity of a hedgehog inhibitor gant61 in estrogen receptor-positive breast cancer cells. Cancer Sci. 2017, 108, 918–930.
- Arnold, K.M.; Pohlig, R.T.; Sims-Mourtada, J. Co-activation of hedgehog and wnt signaling pathways is associated with poor outcomes in triple negative breast cancer. Oncol. Lett. 2017, 14, 5285–5292.
- Jeng, K.S.; Jeng, C.J.; Sheen, I.S.; Wu, S.H.; Lu, S.J.; Wang, C.H.; Chang, C.F. Glioma-associated oncogene homolog inhibitors have the potential of suppressing cancer stem cells of breast cancer. Int. J. Mol. Sci. 2018, 19, 1375.
- Esposito, M.; Mondal, N.; Greco, T.M.; Wei, Y.; Spadazzi, C.; Lin, S.C.; Zheng, H.; Cheung, C.; Magnani, J.L.; Lin, S.H.; et al. Bone vascular niche E-selectin induces mesenchymal–epithelial transition and Wnt activation in cancer cells to promote bone metastasis. Nat. Cell Biol. 2019, 21, 627–639.
- Harper, K.L.; Sosa, M.S.; Entenberg, D.; Hosseini, H.; Cheung, J.F.; Nobre, R.; Avivar-Valderas, A.; Nagi, C.; Girnius, N.; Davis, R.J.; et al. Mechanism of early dissemination and metastasis in Her2+ mammary cancer. Nature 2016, 540, 588–592.
- Pommier, A.; Anaparthy, N.; Memos, N.; Kelley, Z.L.; Gouronnec, A.; Yan, R.; Auffray, C.; Albrengues, J.; Egeblad, M.; Iacobuzio-Donahue, C.A.; et al. Unresolved endoplasmic reticulum stress engenders immune-resistant, latent pancreatic cancer metastases. Science 2018, 360, eaao4908.
- Satcher, R.L.; Zhang, X.H. Evolving cancer-niche interactions and therapeutic targets during bone metastasis. Nat. Rev. Cancer 2022, 22, 85–101.
- Malladi, S.; Macalinao, D.G.; Jin, X.; He, L.; Basnet, H.; Zou, Y.; de Stanchina, E.; Massague, J. Metastatic latency and immune evasion through autocrine inhibition of WNT. Cell 2016, 165, 45–60.
- Zhang, X.H.; Wang, Q.; Gerald, W.; Hudis, C.A.; Norton, L.; Smid, M.; Foekens, J.A.; Massague, J. Latent bone metastasis in breast cancer tied to Src-dependent survival signals. Cancer Cell 2009, 16, 67–78.
- Horak, C.E.; Lee, J.H.; Marshall, J.C.; Shreeve, S.M.; Steeg, P.S. The role of metastasis suppressor genes in metastatic dormancy. APMIS 2008, 116, 586–601.
- Drescher, F.; Juárez, P.; Arellano, D.L.; Serafín-Higuera, N.; Olvera-Rodriguez, F.; Jiménez, S.; Licea-Navarro, A.F.; Fournier, P.G. TIE2 Induces Breast Cancer Cell Dormancy and Inhibits the Development of Osteolytic Bone Metastases. Cancers 2020, 12, 868.
- Yang, C.; Ng, C.T.; Li, D.; Zhang, L. Targeting indoleamine 2,3-dioxygenase 1: Fighting cancers via dormancy regulation. Front. Immunol. 2021, 12, 725204.
- Hamilton, M.M.; Mseeh, F.; McAfoos, T.J.; Leonard, P.G.; Reyna, N.J.; Harris, A.L.; Xu, A.; Han, M.; Soth, M.J.; Czako, B.; et al. Discovery of iacs-9779 and iacs-70465 as potent inhibitors targeting indoleamine 2,3-dioxygenase 1 (ido1) apoenzyme. J. Med. Chem. 2021, 64, 11302–11329.
- Kapoor, A.; Goldberg, M.S.; Cumberland, L.K.; Ratnakumar, K.; Segura, M.F.; Emanuel, P.O.; Menendez, S.; Vardabasso, C.; Leroy, G.; Vidal, C.I.; et al. The histone variant macroH2A suppresses melanoma progression through regulation of cdk8. Nature 2010, 468, 1105–1109.
- Zhang, J.; Yang, C.; Wu, C.; Cui, W.; Wang, L. DNA methyltransferases in cancer: Biology, paradox, aberrations, and targeted therapy. Cancers 2020, 12, 2123.
- Sosa, M.S.; Bernstein, E.; Aguirre-Ghiso, J.A. Epigenetic Regulation of Cancer Dormancy as a Plasticity Mechanism for Metastasis Initiation. In Tumor Dormancy and Recurrence. Cancer Drug Discovery and Development; Wang, Y., Crea, F., Eds.; Humana Press: Totowa, NJ, USA, 2017.
- Arandkar, S.; Furth, N.; Elisha, Y.; Nataraj, N.B.; van der Kuip, H.; Yarden, Y.; Aulitzky, W.; Ulitsky, I.; Geiger, B.; Oren, M. Altered p53 functionality in cancer-associated fibroblasts contributes to their cancer-supporting features. Proc. Natl. Acad. Sci. USA 2018, 11, 6410–6415.
- Borgen, E.; Rypdal, M.C.; Sosa, M.S.; Renolen, A.; Schlichting, E.; Lonning, P.E.; Synnestvedt, M.; Aguirre-Ghiso, J.A.; Naume, B. NR2F1 stratifies dormant disseminated tumor cells in breast cancer patients. Breast Cancer Res. 2018, 20, 120.
- Suh, Y.A.; Lee, H.Y.; Virmani, A.; Wong, J.; Mann, K.K.; Miller, W.H.; Gazdar, A.; Kurie, J.M. Loss of retinoic acid receptor beta gene expression is linked to aberrant histone h3 acetylation in lung cancer cell lines. Cancer Res. 2002, 62, 3945–3949.
- Gao, H.; Chakraborty, G.; Lee-Lim, A.P.; Mavrakis, K.J.; Wendel, H.G.; Giancotti, F.G. Forward genetic screens in mice uncover mediators and suppressors of metastatic reactivation. Proc. Natl. Acad. Sci. USA 2014, 111, 16532–16537.
- Robinson, N.J.; Parker, K.A.; Schiemann, W.P. Epigenetic plasticity in metastatic dormancy: Mechanisms and therapeutic implications. Ann. Transl. Med. 2020, 8, 903.
- Parker, K.A.; Robinson, N.J.; Schiemann, W.P. The role of RNA processing and regulation in metastatic dormancy. Semin. Cancer Biol. 2022, 78, 23–34.
More
Information
Subjects:
Oncology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
891
Revisions:
2 times
(View History)
Update Date:
20 Jun 2023
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No