Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Didem Karakas | -- | 2490 | 2023-06-15 20:10:31 | | | |
| 2 | Catherine Yang | Meta information modification | 2490 | 2023-06-16 02:35:45 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Karakas, D.; Ozpolat, B. LncRNAs in the Regulation of Translation. Encyclopedia. Available online: https://encyclopedia.pub/entry/45679 (accessed on 27 July 2026).
Karakas D, Ozpolat B. LncRNAs in the Regulation of Translation. Encyclopedia. Available at: https://encyclopedia.pub/entry/45679. Accessed July 27, 2026.
Karakas, Didem, Bulent Ozpolat. "LncRNAs in the Regulation of Translation" Encyclopedia, https://encyclopedia.pub/entry/45679 (accessed July 27, 2026).
Karakas, D., & Ozpolat, B. (2023, June 15). LncRNAs in the Regulation of Translation. In Encyclopedia. https://encyclopedia.pub/entry/45679
Karakas, Didem and Bulent Ozpolat. "LncRNAs in the Regulation of Translation." Encyclopedia. Web. 15 June, 2023.
Copy Citation
Long non-coding RNAs (lncRNAs), a group of non-protein coding RNAs with lengths of more than 200 nucleotides, exert their effects by binding to DNA, mRNA, microRNA, and proteins and regulate gene expression at the transcriptional, post-transcriptional, translational, and post-translational levels.
non-coding RNAs
long non-coding RNAs
ncRNAs
translation
cancer
1. Overview of Protein Translation Process
Protein translation is a highly complex process, comprising three steps (initiation, elongation, translation) and each step requires dynamic and efficient interactions between a great number of proteins, RNAs and ribosome.
The initiation process consists of two main steps. The first step involves the formation of the pre-initiation complex, and the second step is the assembling of this complex to the large subunit of the ribosome [1]. The initiation step begins with the formation of a ternary complex (eIF2-GTP-Met-tRNA), then the complex binds to small subunit (40S) of ribosome and assembles a pre-initiation complex by binding to other initiation factors (eIF1, eIF1A, eIF3, and eIF5) [2][3]. Before the pre-initiation complex directs to the 5′ end of mRNA, eIF4F complex, which is formed by eIF4E (cap-binding protein), eIF4G (scaffold protein) and eIF4A (helicase), bind to the 5′ end of mRNA to unwind and activate it [1][4]. The formation of eIF4F complex is maintained by some other initiation factors, eIF4B and eIF3. The pre-initiation complex then scans the mRNA until it recognizes a start codon [5]. Once the start codon is recognized, eIF5 and eIF5B promote hydrolysis of eIF2-bound GTP, releasing of eIFs from the complex and joining to the large subunit of the ribosome [6]. Following the initiation step of translation, met-tRNA reaches the P (peptidyl)-site of the 80S ribosome awaiting amino acids for elongation of the peptide chain.
The elongation step of translation requires the recruitment of aminoacyl-tRNA to the A (aminoacyl)-site of ribosome through GTP-bound eukaryotic elongation factor 1A (eEF1A). Although there is no base-pairing between tRNA anticodon and A-site codon, tRNA generates a codon-anticodon helix by remodeling itself [7] and stabilizes the ternary complex (aa-tRNA-eIF1A-GTP) [8]. Base-pairing interactions between A-site codon and aa-tRNA anticodon induce hydrolysis of GTP by eEF1A, which is then released from the A-site of the ribosome. eEF1A-GDP complex is recycled by eEF1B. Following the transfer of aa-tRNA to the A-site, a conformational change occurs in the ribosome which facilitates the formation of peptide bond between the aa-tRNA and the tRNA carrying the Met-tRNA at the P site. A GTPase (eEF2) binds to the A-site of the ribosome, hydrolyzes GTP and stimulates a conformational change in the ribosome resulting in movement of the ribosome one codon further. After the translocation of the ribosome, the A-site becomes empty and can accept the next aa-tRNAs to start a new cycle of elongation [7].
The last step of protein translation is termination, which begins when a stop codon (UAA, UGA, or UAG) reaches the A-site of the ribosome. Two types of release factors, eRF1 and eRF3, are involved in the termination process [9][10][11]. eRF1 is responsible for the recognition of stop codon and stimulation of peptide release, while eRF3 binds to eRF1 and triggers eRF1-mediated peptide release via GTPase activity [11][12]. The ternary complex (eRF1-eRF3-GTP) then binds to the ribosomal pre-termination complex and eRF3 hydrolyses GTP to release polypeptide [13].
2. Regulation of Translational Factors by LncRNAs
2.1. Inhibitory Roles of LncRNAs in Translation through Regulation of Translation Factors
A growing body of evidence demonstrates that lncRNAs can regulate each step of translation by regulating the expression and the function of translation factors. For instance, lncRNA GAS5 is involved in the regulation of apoptosis and cell proliferation. A study performed with lymphoma cells showed that GAS5 interacts with the translation initiation complex, eIF4F, by directly binding to eIF4E and decreasing the translation of c-Myc [14]. Similarly, lncRNA RP1-5O6.5 has been shown to interact with eIF4E and prevents binding of eIF4E to eIF4G, leading to inhibition of translation of p27kip1, which negatively regulates Snail levels in breast cancer cells [15]. lncRNAs SNHG1 and SNGH4 are capable of binding to eIF4E and dysregulate the function of eIF4E in mantle cell lymphoma cells [16]. In the other example, lncRNA treRNA has been shown to interact with ribonucleoproteins (RNPs) (hnRNP K, FXR1, FXR2, PUF60, and SF3B3) and form treRNA-RNP complex which suppresses the translation efficiency of E-cadherin by binding eIF4G1 [17]. A brain-specific lncRNA, BC1, has been reported to interact with eIF4A and poly(A)-binding protein (PABP) and negatively regulate translation process [18][19]. lncRNA GAPLINC is overexpressed in non-small lung cancer cells and it increases eEF2K expression (a negative regulator of eEF2) by acting as a sponge for miR-661 [20]. In the other study, lncRNA FOXD1-AS1 was shown to bind to eIF5A, however it did not change the mRNA expression levels, suggesting that FOXD1-AS1 can involve in the post-translational regulation [21]. Overall, these studies suggested that lncRNAs can play an important inhibitory roles in mRNA translation through regulation of translation factors.
2.2. LncRNAs Positively Regulate Protein Translation
Some lncRNAs have been reported to positively regulate protein translation. For instance, lncRNA SRA enhanced Wnt/β-catenin signaling pathway by increasing the expression of eIF4E-binding protein 1 (eIF4E-BP1) and contributed to the aggressive characteristics of endometrial cancer [22]. Another study showed that lncRNA MCM3AP-AS1 enhances the expression of eIF4E by acting as a sponge for miR15a, which supresses eIF4E expression and contributes to doxorubicin resistance in Burkitt lymphoma cells through MCM3AP-AS1/miR-15a/eIF4E axis [23]. Similarly, lncRNA SNHG12 enhanced the invasion of human vascular smooth muscle cells by serving as a sponge of miR-766-5p and influencing the miR-766-5p/eIF5A axis [24]. In the other study, a Y-linked lncRNA, LINC00278, was found to encode a micropeptide called YY1BM which led to a decrease in the expression of negative regulator of translation, eEF2K [25]. The functions of lncRNAs on translational factors are summarized in Table 1.
Table 1. The list of long non-coding RNAs (lncRNAs) involved in regulation of translational factors [14][15][16][17][18][19][20][21][22][23][24][25].
| LncRNA | Translation Factor | Function | Reference |
|---|---|---|---|
| GAS5 | Binds to eIF4E and prevents formation of initiation complex (eIF4F) | Decreases translation of c-Myc | [14] |
| RP1-5O6.5 | Interacts with eIF4E and prevents binding to eIF4G | Promotes breast cancer metastasis by inhibiting translation of p27Kip1 | [15] |
| SNHG1 and SNGH4 | Bind to eIF4E and dysregulate it | Enhance translation and contribute aggressiveness of lymphoma cells | [16] |
| treRNA | Promotes the formation of a treRNA-associated protein (treRNP) complex and suppresses translation by binding to eEIF4G1 | treRNP complex reduces translation efficiency of E-cadherin and decreases tumor metastasis | [17] |
| BC1 | Interacts with eIF4A and poly(A)-binding protein (PABP) | Represses translation | [18][19] |
| GAPLINC | Positively regulates eEF2K expression by sponging miR-661 | Promotes tumorigenesis of non-small cell lung cancer cells | [20] |
| SRA | Binds and increases the expression of eIF4E-binding protein 1 (eIF4E-BP1) | Increases the activity of Wnt/ β-catenin signaling and promotes aggressive characteristics of endometrial cancer | [22] |
| MCM3AP-AS1 | Positively regulates the expression of eIF4E by using miR15a as a sponge | Promotes translation and contributes doxorubicin resistance | [23] |
| SNGH12 | Binds to miR-766-5p, which is a negative regulator of eIF5A | Targets miR-766-5p/eIF5A axis and enhances invasion of vascular smooth muscle cells | [24] |
| LNC00278 | Decreases eEF2K expression | Micropeptide of lncRNA, YY1BM, represses the eEF2K/eEF2 axis | [25] |
3. LncRNAs Involved in Signaling Pathways Regulating Protein Translation
The PI3K/AKT/mTOR is one of the major signaling pathways known to regulate vital cellular processes including cell proliferation, growth, survival, metabolism and protein translation. The role of PI3K/AKT/mTOR and MAPK pathways in the regulation of translational machinery are well documented and they are frequently overactivated in most types of cancer [26]. Both pathways involve the mechanistic target of rapamycin (mTOR) to regulate a variety of components of the translational machinery in homeostasis, their dysregulation results in aberrant translation which is often detected in diabetes, neurological disorders, and cancer [27][28][29][30]. The MAPK family consists of a serine/threonine kinases, that includes ERKs, JNKs and p38/SAPKs [31]. Especially the MAPK/ERK signaling pathway is amongst the most well-studied, signaling and dysregulating one-third of all human cancers [32].
PI3K/AKT/mTOR pathway regulates cell growth and proliferation by phosphorylating two downstream targets which are 4E-BP1 and ribosomal protein S6 kinase (S6Ks). mTOR complex I (mTORC1) controls translational activation by phosphorylating eIF4E inhibitor, 4E-BP1, which releases eIF4E to interact with initiation complex (eIF4F) [33]. S6K protein requires sequential phosphorylations at multiple serine/threonine sites and mTORC1 regulates its activation by phosphorylation. Once S6K is activated, it phosphorylates and activates eIF4B, which increases the recruitment of eIF4B to eEF4A and enhances translation [34]. Besides, S6K and mTORC1 signaling pathways can phosphorylate EF2-Kinase (EF2K) and decrease its sensitivity to Ca/Calmoduline for its activation [35]. Similarly, eEF2K activity is negatively regulated by MAPKs and their downstream effectors, reducing phosphorylation of eEF2, leading to increased translation by promoting peptide elongation phase of protein systhesis [36][37]. Considering the significant regulatory roles of PI3K/AKT/mTOR and MAPK signaling pathways in protein translation, regulation of their activity by lncRNAs indicate that the lncRNAs are involved in controlling protein translation through regulation of these key signaling pathways. For instance, lncRNA UASR1 promotes cell growth and migration of breast cancer cells by regulating AKT/mTOR pathway [38]. In these cells, active mediators of this pathway such as p-AKT, p-TSC2, p-4EBP1 and p-p70S6K are increased by overexpression of UASR1. Thus, UASR1 plays an oncogenic role in breast cancer cells through activation of the AKT/mTOR signaling pathway. Another lncRNA H19 is overexpressed in colorectal cancer tissues and it promotes the activity of PI3K/AKT pathway by acting as a ceRNA and regulating some components of this pathway. H19 regulates various cancer-related mRNAs (such as (AKT3, CSF1, MET, COL1A1) by competitively sponging various miRNAs. Knockdown of H19 reduced protein level of MET, ZEB1, and COL1A1 in vitro [39]. The other study showed that H19 inhibits mTORC1-mediated 4E-BP1 phosphorylation, but it does not affect the activation of S6K1 [40]. lncRNA CASC9 has been shown to suppress apoptosis and promote aggressiveness of oral squamous cell carcinoma cells by activating the AKT/mTOR pathway [41].
In contrast, some lncRNAs might negatively regulate the abovementioned pathways. For instance, lncRNA FER1L4 suppresses cell proliferation and metastasis through downregulating the expressions of PI3K and AKT in lung cancer cells [42]. Overall, lncRNAs can regulate signaling pathways involved in translational control that is an integral part of these survival adaptive pathways in normal and cancer cells. Some of these regulatory lncRNAs and their functions on signaling pathways are summarized in Table 2.
Table 2. lncRNAs in the regulation of signaling pathways and their roles in various cancers [43][44][45][46][47][48][49][50][51][52].
| LncRNA | Target | Function | Reference |
|---|---|---|---|
| MALAT1 | mTOR signaling | Improves glucose metabolism to contribute aggressiveness in hepatocellular carcinoma cells | [43] |
| HOXB-AS3 | PI3K/AKT signaling | Increases proliferation, migration, and invasion of lung cancer cells | [44] |
| AK023391 | PI3K/AKT signaling | Promotes tumorigenesis and invasion of gastric cancer | [45] |
| LOC101928316 | PI3K/AKT/mTOR signaling | Inhibits cell proliferation, invasion and tumorigenesis of gastric cancer cells | [46] |
| UCA1 | PI3K/AKT signaling | Promotes cell proliferation and inhibits apoptosis in retinoblastoma cells | [47] |
| OECC | PI3K/AKT/mTOR signaling | Increases proliferation, migration and invasion of lung cancer cells | [48] |
| GAS5 | PTEN/PI3K/AKT signaling | Suppresses proliferation and invasion of osteosarcoma cells and promotes PTEN expression by sponging miR-23a-3p | [49] |
| LINC01503 | MAPK/ERK signaling | Increases proliferation and tumor forming-ability of hepatocellular carcinoma cells | [50] |
| ST8SIA6-AS1 | p38 MAPK signaling | Promotes proliferation, migration and invasion of breast cancer cells | [51] |
| FENDRR | p38 MAPK signaling | Inhibits cell proliferation and induces apoptosis in hepatocellular carcinoma cells | [52] |
4. LncRNAs in Cancer
4.1. LncRNAs Can Contribute Hallmarks of Cancer
Deregulation of mRNA translation is commonly observed in malignant cells and is considered as a critical factor contributing to cancer initiation, tumorigenesis, and progression. Because lncRNAs play critical roles in the regulation of a wide range of cellular processes, their dysregulation is associated with cell proliferation, survival, tumorigenesis and progression of various cancers, and aberrant expression of lncRNAs can contribute to the hallmarks of cancer. Reprograming of the translation machinery in cancer cells is important function of the key oncogenic signalings, promoting cellular transformation. Increased activity of translational machinery has been shown to be critical in many cancer cells, including breast [53], pancreatic [54], liver [55], and colorectal cancer [56], and leukemia [57]. Thus, lncRNA-mediated regulation of protein translation plays an important role in promoting oncogenic signaling, and specific targeting of these lncRNAs holds promise for developing highly targeted therapies in cancer and other human diseases. Figure 1 illustrates some of the lncRNAs that are involved in tumorigenesis and cancer progression.

Figure 1. Some lncRNAs are identified to be involved in aggressive characteristics of some common types of cancers [17][58][59][60][61][62][63][64][65][66][67][68][69][70][71][72][73][74][75][76][77][78][79][80][81][82][83][84][85][86][87][88][89][90][91][92][93][94][95][96][97][98][99][100][101][102][103].
4.2. The Functions of LncRNAs in Regulating Translation of Cancer-Related Proteins
As mentioned above, various lncRNAs are involved in the regulation of hallmarks of cancer, suggesting that they have potential regulatory roles in cancer-related protein tranlation.
An example of a lncRNA that is well-known to be associated with cancer is MALAT1. MALAT1 was shown to upregulate the expression of glycolytic genes which contributes the aggressive characteristics of hepatocellular carcinoma cells. MALAT1 regulated the glucose metabolism of hepatocellular carcinoma cells by enhancing translation of metabolic transcription factor TCF7L2 through mTORC1–4EBP1 axis [43]. lncRNA NEAT1 represents another example of lncRNAs that contribute to the aggressiveness of non-small cell lung cancer by enhancing eIF4G2 via miR-582-5p sponging effects [104]. Similarly, lncRNA RP11-284P20.2 enhanced c-met mRNA translation by recruiting eIF3b to c-met and thus promoted proliferation and invasion of hepatocellular carcinoma cells [105]. In prostate cancer, lncRNA UCA1 levels were found to be positively correlated with eIF4G1 levels. UCA1 enhances eIF4G1 levels via sponging miR-331-3p, while knockingdown of UCA1 sensitizes prostate cancer cells to radiotherapy by suppressing eIF4G1 expression via miR-331-3p/eIF4G1 axis [106]. In another study, lncRNA GAPLINC increased the eEF2K expression by serving as a sponge for miR-661, thereby promoted proliferation and progression of non-small cell lung cancer [20].
lncRNAs can also regulate translation process by interacting with the ribosome or ribosome-related proteins. For example, lncRNA ZFAS1 was shown to interact with a small 40S subunit of the ribosome in breast cancer cells. The study showed that ZFAS1 did not regulate translation process directly. Instead, the lncRNA was increased during the ribosome biogenesis indicating its role in regulating the ribosome production and assembly [107]. In neuroblastoma cells, it was shown that lncNB1 enhanced E2F1 protein synthesis and N-Myc stability by binding the ribosomal protein RPL35 [108].
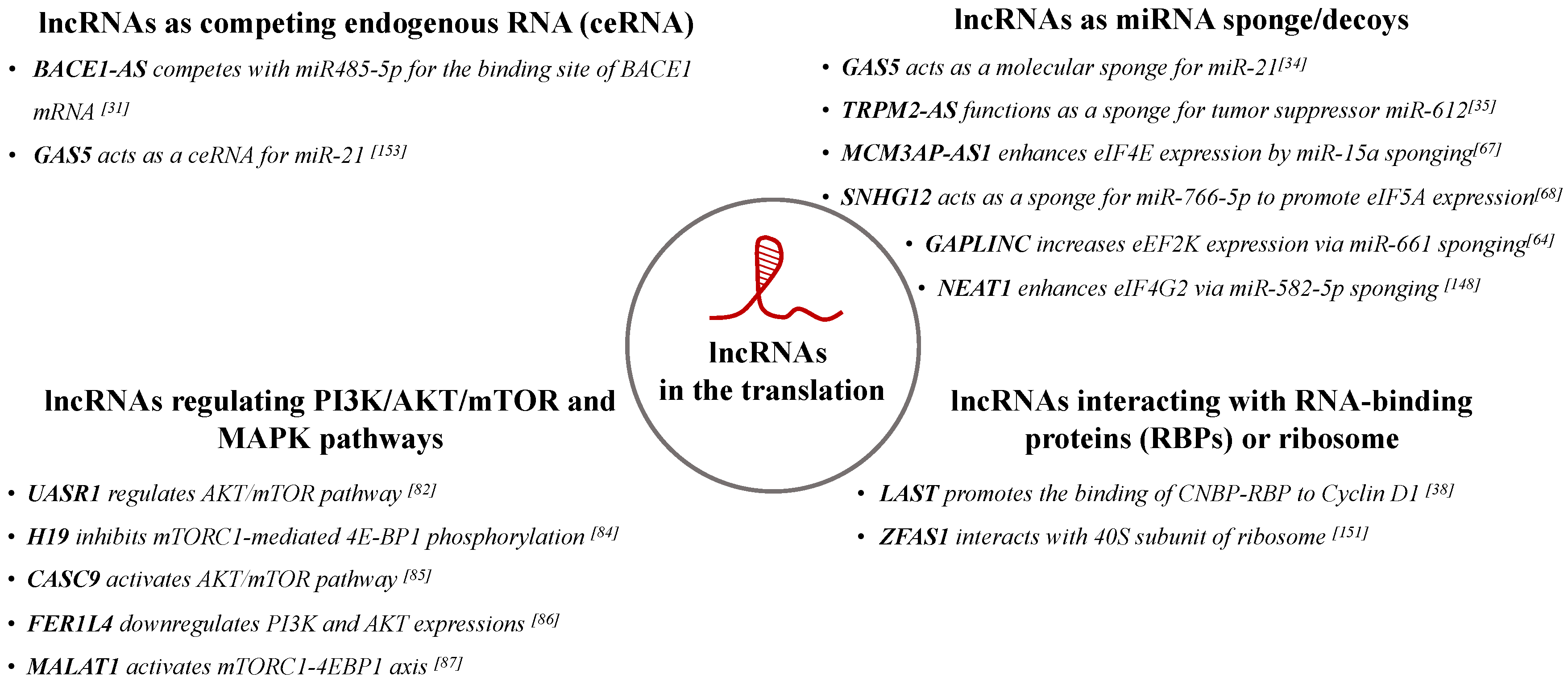
Overall, an emerging body of evidence suggests that lncRNAs play important roles in the regulation of protein translation process. They can enhance or suppress translation via several mechanisms, including through interacting with the ribosome-associated proteins, sponging miRNAs, and competing with endogenous RNAs. Their mechanisms of action and some examples are summarized in Figure 2.
References
- Jackson, R.J.; Hellen, C.U.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127.
- Dever, T.E. Gene-specific regulation by general translation factors. Cell 2002, 108, 545–556.
- Ali, M.U.; Ur Rahman, M.S.; Jia, Z.; Jiang, C. Eukaryotic translation initiation factors and cancer. Tumor Biol. 2017, 39, 1010428317709805.
- Grifo, J.A.; Tahara, S.M.; Morgan, M.A.; Shatkin, A.J.; Merrick, W.C. New initiation factor activity required for globin mRNA translation. J. Biol. Chem. 1983, 258, 5804–5810.
- Hinnebusch, A.G. Molecular mechanism of scanning and start codon selection in eukaryotes. Microbiol. Mol. Biol. Rev. 2011, 75, 434–467.
- Llácer, J.L.; Hussain, T.; Saini, A.K.; Nanda, J.S.; Kaur, S.; Gordiyenko, Y.; Kumar, R.; Hinnebusch, A.G.; Lorsch, J.R.; Ramakrishnan, V. Translational initiation factor eIF5 replaces eIF1 on the 40S ribosomal subunit to promote start-codon recognition. eLife 2018, 7, e39273.
- Voorhees, R.M.; Ramakrishnan, V. Structural basis of the translational elongation cycle. Annu. Rev. Biochem. 2013, 82, 203–236.
- Rodnina, M.V.; Gromadski, K.B.; Kothe, U.; Wieden, H.J. Recognition and selection of tRNA in translation. FEBS Lett. 2005, 579, 938–942.
- Stansfield, I.; Jones, K.M.; Kushnirov, V.V.; Dagkesamanskaya, A.R.; Poznyakovski, A.I.; Paushkin, S.V.; Nierras, C.R.; Cox, B.S.; Ter-Avanesyan, M.D.; Tuite, M.F. The products of the SUP45 (eRF1) and SUP35 genes interact to mediate translation termination in Saccharomyces cerevisiae. EMBO J. 1995, 14, 4365–4373.
- Zhouravleva, G.; Frolova, L.; Le Goff, X.; Le Guellec, R.; Inge-Vechtomov, S.; Kisselev, L.; Philippe, M. Termination of translation in eukaryotes is governed by two interacting polypeptide chain release factors, eRF1 and eRF3. EMBO J. 1995, 14, 4065–4072.
- Alkalaeva, E.Z.; Pisarev, A.V.; Frolova, L.Y.; Kisselev, L.L.; Pestova, T.V. In vitro reconstitution of eukaryotic translation reveals cooperativity between release factors eRF1 and eRF3. Cell 2006, 125, 1125–1136.
- Salas-Marco, J.; Bedwell, D.M. GTP hydrolysis by eRF3 facilitates stop codon decoding during eukaryotic translation termination. Mol. Cell Biol. 2004, 24, 7769–7778.
- Frolova, L.; Le Goff, X.; Zhouravleva, G.; Davydova, E.; Philippe, M.; Kisselev, L. Eukaryotic polypeptide chain release factor eRF3 is an eRF1- and ribosome-dependent guanosine triphosphatase. RNA 1996, 2, 334–341.
- Hu, G.; Lou, Z.; Gupta, M. The long non-coding RNA GAS5 cooperates with the eukaryotic translation initiation factor 4E to regulate c-Myc translation. PLoS ONE 2014, 9, e107016.
- Jia, X.; Shi, L.; Wang, X.; Luo, L.; Ling, L.; Yin, J.; Song, Y.; Zhang, Z.; Qiu, N.; Liu, H.; et al. KLF5 regulated lncRNA RP1 promotes the growth and metastasis of breast cancer via repressing p27kip1 translation. Cell Death Dis. 2019, 10, 373.
- Hu, G.; Witzig, T.E.; Gupta, M. Novel Long Non-Coding RNA, SNHG4 Complex With Eukaryotic Initiation Factor-4E and Regulate Aberrant Protein Translation In Mantle Cell Lymphoma: Implications For Novel Biomarker. Blood 2013, 122, 81.
- Gumireddy, K.; Li, A.; Yan, J.; Setoyama, T.; Johannes, G.J.; Orom, U.A.; Tchou, J.; Liu, Q.; Zhang, L.; Speicher, D.W.; et al. Identification of a long non-coding RNA-associated RNP complex regulating metastasis at the translational step. EMBO J. 2013, 32, 2672–2684.
- Wang, H.; Iacoangeli, A.; Popp, S.; Muslimov, I.A.; Imataka, H.; Sonenberg, N.; Lomakin, I.B.; Tiedge, H. Dendritic BC1 RNA: Functional role in regulation of translation initiation. J. Neurosci. 2002, 22, 10232–10241.
- Wang, H.; Iacoangeli, A.; Lin, D.; Williams, K.; Denman, R.B.; Hellen, C.U.; Tiedge, H. Dendritic BC1 RNA in translational control mechanisms. J. Cell Biol. 2005, 171, 811–821.
- Gu, H.; Chen, J.; Song, Y.; Shao, H. Gastric Adenocarcinoma Predictive Long Intergenic Non-Coding RNA Promotes Tumor Occurrence and Progression in Non-Small Cell Lung Cancer via Regulation of the miR-661/eEF2K Signaling Pathway. Cell Physiol. Biochem. 2018, 51, 2136–2147.
- Gao, Y.F.; Liu, J.Y.; Mao, X.Y.; He, Z.W.; Zhu, T.; Wang, Z.B.; Li, X.; Yin, J.Y.; Zhang, W.; Zhou, H.H.; et al. LncRNA FOXD1-AS1 acts as a potential oncogenic biomarker in glioma. CNS Neurosci. Ther. 2020, 26, 66–75.
- Park, S.A.; Kim, L.K.; Kim, Y.T.; Heo, T.H.; Kim, H.J. Long non-coding RNA steroid receptor activator promotes the progression of endometrial cancer via Wnt/β-catenin signaling pathway. Int. J. Biol. Sci. 2020, 16, 99–115.
- Guo, C.; Gong, M.; Li, Z. Knockdown of lncRNA MCM3AP-AS1 Attenuates Chemoresistance of Burkitt Lymphoma to Doxorubicin Treatment via Targeting the miR-15a/EIF4E Axis. Cancer Manag. Res. 2020, 12, 5845–5855.
- Liu, W.; Che, J.; Gu, Y.; Song, L. LncRNA SNHG12 promotes proliferation and migration of vascular smooth muscle cells via targeting miR-766-5p/EIF5A. Res. Square 2020.
- Wu, S.; Zhang, L.; Deng, J.; Guo, B.; Li, F.; Wang, Y.; Wu, R.; Zhang, S.; Lu, J.; Zhou, Y. A Novel Micropeptide Encoded by Y-Linked LINC00278 Links Cigarette Smoking and AR Signaling in Male Esophageal Squamous Cell Carcinoma. Cancer Res. 2020, 80, 2790–2803.
- Janku, F.; Yap, T.A.; Meric-Bernstam, F. Targeting the PI3K pathway in cancer: Are we making headway? Nat. Rev. Clin. Oncol. 2018, 15, 273–291.
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys. Acta 2010, 1802, 396–405.
- Fruman, D.A.; Chiu, H.; Hopkins, B.D.; Bagrodia, S.; Cantley, L.C.; Abraham, R.T. The PI3K Pathway in Human Disease. Cell 2017, 170, 605–635.
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618.
- Xu, F.; Na, L.; Li, Y.; Chen, L. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci. 2020, 10, 54.
- Morrison, D.K. MAP kinase pathways. Cold Spring Harb. Perspect. Biol. 2012, 4, a011254.
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290.
- Heesom, K.J.; Denton, R.M. Dissociation of the eukaryotic initiation factor-4E/4E-BP1 complex involves phosphorylation of 4E-BP1 by an mTOR-associated kinase. FEBS Lett. 1999, 457, 489–493.
- Holz, M.K.; Ballif, B.A.; Gygi, S.P.; Blenis, J. mTOR and S6K1 mediate assembly of the translation preinitiation complex through dynamic protein interchange and ordered phosphorylation events. Cell 2005, 123, 569–580.
- Browne, G.J.; Proud, C.G. A novel mTOR-regulated phosphorylation site in elongation factor 2 kinase modulates the activity of the kinase and its binding to calmodulin. Mol. Cell. Biol. 2004, 24, 2986–2997.
- Knebel, A.; Morrice, N.; Cohen, P. A novel method to identify protein kinase substrates: eEF2 kinase is phosphorylated and inhibited by SAPK4/p38delta. EMBO J. 2001, 20, 4360–4369.
- Knebel, A.; Haydon, C.E.; Morrice, N.; Cohen, P. Stress-induced regulation of eEF2 kinase by SB203580-sensitive and -insensitive pathways. Biochem. J. 2002, 367, 525–532.
- Cao, Z.; Wu, P.; Su, M.; Ling, H.; Khoshaba, R.; Huang, C.; Gao, H.; Zhao, Y.; Chen, J.; Liao, Q.; et al. Long non-coding RNA UASR1 promotes proliferation and migration of breast cancer cells through the AKT/mTOR pathway. J. Cancer 2019, 10, 2025–2034.
- Zhong, M.E.; Chen, Y.; Zhang, G.; Xu, L.; Ge, W.; Wu, B. LncRNA H19 regulates PI3K-Akt signal pathway by functioning as a ceRNA and predicts poor prognosis in colorectal cancer: Integrative analysis of dysregulated ncRNA-associated ceRNA network. Cancer Cell Int. 2019, 19, 148.
- Wu, Z.R.; Yan, L.; Liu, Y.T.; Cao, L.; Guo, Y.H.; Zhang, Y.; Yao, H.; Cai, L.; Shang, H.B.; Rui, W.W.; et al. Inhibition of mTORC1 by lncRNA H19 via disrupting 4E-BP1/Raptor interaction in pituitary tumours. Nat. Commun. 2018, 9, 4624.
- Yang, Y.; Chen, D.; Liu, H.; Yang, K. Increased expression of lncRNA CASC9 promotes tumor progression by suppressing autophagy-mediated cell apoptosis via the AKT/mTOR pathway in oral squamous cell carcinoma. Cell Death Dis. 2019, 10, 41.
- Gao, X.; Wang, N.; Wu, S.; Cui, H.; An, X.; Yang, Y. Long non-coding RNA FER1L4 inhibits cell proliferation and metastasis through regulation of the PI3K/AKT signaling pathway in lung cancer cells. Mol. Med. Rep. 2019, 20, 182–190.
- Malakar, P.; Stein, I.; Saragovi, A.; Winkler, R.; Stern-Ginossar, N.; Berger, M.; Pikarsky, E.; Karni, R. Long Noncoding RNA MALAT1 Regulates Cancer Glucose Metabolism by Enhancing mTOR-Mediated Translation of TCF7L2. Cancer Res. 2019, 79, 2480–2493.
- Jiang, W.; Kai, J.; Li, D.; Wei, Z.; Wang, Y.; Wang, W. lncRNA HOXB-AS3 exacerbates proliferation, migration, and invasion of lung cancer via activating the PI3K-AKT pathway. J. Cell Physiol. 2020, 235, 7194–7203.
- Huang, Y.; Zhang, J.; Hou, L.; Wang, G.; Liu, H.; Zhang, R.; Chen, X.; Zhu, J. LncRNA AK023391 promotes tumorigenesis and invasion of gastric cancer through activation of the PI3K/Akt signaling pathway. J. Exp. Clin. Cancer Res. 2017, 36, 194.
- Li, C.; Liang, G.; Yang, S.; Sui, J.; Wu, W.; Xu, S.; Ye, Y.; Shen, B.; Zhang, X.; Zhang, Y. LncRNA-LOC101928316 contributes to gastric cancer progression through regulating PI3K-Akt-mTOR signaling pathway. Cancer Med. 2019, 8, 4428–4440.
- Yuan, Z.; Li, Z. Long noncoding RNA UCA1 facilitates cell proliferation and inhibits apoptosis in retinoblastoma by activating the PI3K/Akt pathway. Transl. Cancer Res. 2020, 9.
- Zou, Y.; Zhang, B.; Mao, Y.; Zhang, H.; Hong, W. Long non-coding RNA OECC promotes cell proliferation and metastasis through the PI3K/Akt/mTOR signaling pathway in human lung cancer. Oncol. Lett. 2019, 18, 3017–3024.
- Liu, J.; Chen, M.; Ma, L.; Dang, X.; Du, G. LncRNA GAS5 Suppresses the Proliferation and Invasion of Osteosarcoma Cells via the miR-23a-3p/PTEN/PI3K/AKT Pathway. Cell Transplant. 2020, 29, 963689720953093.
- Wang, M.R.; Fang, D.; Di, M.P.; Guan, J.L.; Wang, G.; Liu, L.; Sheng, J.Q.; Tian, D.A.; Li, P.Y. Long non-coding RNA LINC01503 promotes the progression of hepatocellular carcinoma via activating MAPK/ERK pathway. Int. J. Med. Sci. 2020, 17, 1224–1234.
- Fang, K.; Hu, C.; Zhang, X.; Hou, Y.; Gao, D.; Guo, Z.; Li, L. LncRNA ST8SIA6-AS1 promotes proliferation, migration and invasion in breast cancer through the p38 MAPK signalling pathway. Carcinogenesis 2020, 41, 1273–1281.
- Qian, G.; Jin, X.; Zhang, L. LncRNA FENDRR Upregulation Promotes Hepatic Carcinoma Cells Apoptosis by Targeting miR-362-5p Via NPR3 and p38-MAPK Pathway. Cancer Biother. Radiopharm. 2020, 35, 629–639.
- Zhang, T.; Hu, H.; Yan, G.; Wu, T.; Liu, S.; Chen, W.; Ning, Y.; Lu, Z. Long Non-Coding RNA and Breast Cancer. Technol. Cancer Res. Treat. 2019, 18, 1533033819843889.
- Zhou, W.; Chen, L.; Li, C.; Huang, R.; Guo, M.; Ning, S.; Ji, J.; Guo, X.; Lou, G.; Jia, X.; et al. The multifaceted roles of long noncoding RNAs in pancreatic cancer: An update on what we know. Cancer Cell Int. 2020, 20, 41.
- Huang, Z.; Zhou, J.K.; Peng, Y.; He, W.; Huang, C. The role of long noncoding RNAs in hepatocellular carcinoma. Mol. Cancer 2020, 19, 77.
- Siddiqui, H.; Al-Ghafari, A.; Choudhry, H.; Al Doghaither, H. Roles of long non-coding RNAs in colorectal cancer tumorigenesis: A Review. Mol. Clin. Oncol. 2019, 11, 167–172.
- Gao, J.; Wang, F.; Wu, P.; Chen, Y.; Jia, Y. Aberrant LncRNA Expression in Leukemia. J. Cancer 2020, 11, 4284–4296.
- Cheng, D.; Fan, J.; Ma, Y.; Zhou, Y.; Qin, K.; Shi, M.; Yang, J. LncRNA SNHG7 promotes pancreatic cancer proliferation through ID4 by sponging miR-342-3p. Cell Biosci. 2019, 9, 28.
- Wei, W.; Liu, Y.; Lu, Y.; Yang, B.; Tang, L. LncRNA XIST Promotes Pancreatic Cancer Proliferation Through miR-133a/EGFR. J. Cell Biochem. 2017, 118, 3349–3358.
- Berteaux, N.; Lottin, S.; Monté, D.; Pinte, S.; Quatannens, B.; Coll, J.; Hondermarck, H.; Curgy, J.J.; Dugimont, T.; Adriaenssens, E. H19 mRNA-like noncoding RNA promotes breast cancer cell proliferation through positive control by E2F1. J. Biol. Chem. 2005, 280, 29625–29636.
- Tang, J.; Li, Y.; Sang, Y.; Yu, B.; Lv, D.; Zhang, W.; Feng, H. LncRNA PVT1 regulates triple-negative breast cancer through KLF5/beta-catenin signaling. Oncogene 2018, 37, 4723–4734.
- Zhong, H.; Yang, J.; Zhang, B.; Wang, X.; Pei, L.; Zhang, L.; Lin, Z.; Wang, Y.; Wang, C. LncRNA GACAT3 predicts poor prognosis and promotes cell proliferation in breast cancer through regulation of miR-497/CCND2. Cancer Biomark. 2018, 22, 787–797.
- Zhang, M.; Wu, W.B.; Wang, Z.W.; Wang, X.H. lncRNA NEAT1 is closely related with progression of breast cancer via promoting proliferation and EMT. Eur. Rev. Med. Pharmacol. Sci. 2017, 21, 1020–1026.
- You, Z.; Liu, C.; Wang, C.; Ling, Z.; Wang, Y.; Wang, Y.; Zhang, M.; Chen, S.; Xu, B.; Guan, H.; et al. LncRNA CCAT1 Promotes Prostate Cancer Cell Proliferation by Interacting with DDX5 and MIR-28-5P. Mol. Cancer Ther. 2019, 18, 2469–2479.
- Yang, B.; Gao, G.; Wang, Z.; Sun, D.; Wei, X.; Ma, Y.; Ding, Y. Long non-coding RNA HOTTIP promotes prostate cancer cells proliferation and migration by sponging miR-216a-5p. Biosci. Rep. 2018, 38, BSR20180566.
- Zhang, Y.; Su, X.; Kong, Z.; Fu, F.; Zhang, P.; Wang, D.; Wu, H.; Wan, X.; Li, Y. An androgen reduced transcript of LncRNA GAS5 promoted prostate cancer proliferation. PLoS ONE 2017, 12, e0182305.
- Park, S.M.; Choi, E.Y.; Bae, D.H.; Sohn, H.A.; Kim, S.Y.; Kim, Y.J. The LncRNA EPEL Promotes Lung Cancer Cell Proliferation Through E2F Target Activation. Cell Physiol. Biochem. 2018, 45, 1270–1283.
- Nie, F.Q.; Sun, M.; Yang, J.S.; Xie, M.; Xu, T.P.; Xia, R.; Liu, Y.W.; Liu, X.H.; Zhang, E.B.; Lu, K.H.; et al. Long noncoding RNA ANRIL promotes non-small cell lung cancer cell proliferation and inhibits apoptosis by silencing KLF2 and P21 expression. Mol. Cancer Ther. 2015, 14, 268–277.
- Feng, C.; Zhao, Y.; Li, Y.; Zhang, T.; Ma, Y.; Liu, Y. LncRNA MALAT1 Promotes Lung Cancer Proliferation and Gefitinib Resistance by Acting as a miR-200a Sponge. Arch. Bronconeumol. 2019, 55, 627–633.
- Wu, R.; Li, L.; Bai, Y.; Yu, B.; Xie, C.; Wu, H.; Zhang, Y.; Huang, L.; Yan, Y.; Li, X.; et al. The long noncoding RNA LUCAT1 promotes colorectal cancer cell proliferation by antagonizing Nucleolin to regulate MYC expression. Cell Death Dis. 2020, 11, 908.
- Wang, Y.; Lu, Z.; Wang, N.; Feng, J.; Zhang, J.; Luan, L.; Zhao, W.; Zeng, X. Long noncoding RNA DANCR promotes colorectal cancer proliferation and metastasis via miR-577 sponging. Exp. Mol. Med. 2018, 50, 1–17.
- Liu, Z.; Sun, M.; Lu, K.; Liu, J.; Zhang, M.; Wu, W.; De, W.; Wang, Z.; Wang, R. The long noncoding RNA HOTAIR contributes to cisplatin resistance of human lung adenocarcinoma cells via downregualtion of p21(WAF1/CIP1) expression. PLoS ONE 2013, 8, e77293.
- Pan, Y.; Chen, J.; Tao, L.; Zhang, K.; Wang, R.; Chu, X.; Chen, L. Long noncoding RNA ROR regulates chemoresistance in docetaxel-resistant lung adenocarcinoma cells via epithelial mesenchymal transition pathway. Oncotarget 2017, 8, 33144–33158.
- Wang, Y.; Zhang, D.; Wu, K.; Zhao, Q.; Nie, Y.; Fan, D. Long noncoding RNA MRUL promotes ABCB1 expression in multidrug-resistant gastric cancer cell sublines. Mol. Cell Biol. 2014, 34, 3182–3193.
- Chen, J.; Zhang, K.; Song, H.; Wang, R.; Chu, X.; Chen, L. Long noncoding RNA CCAT1 acts as an oncogene and promotes chemoresistance in docetaxel-resistant lung adenocarcinoma cells. Oncotarget 2016, 7, 62474–62489.
- Jiang, Y.Z.; Liu, Y.R.; Xu, X.E.; Jin, X.; Hu, X.; Yu, K.D.; Shao, Z.M. Transcriptome Analysis of Triple-Negative Breast Cancer Reveals an Integrated mRNA-lncRNA Signature with Predictive and Prognostic Value. Cancer Res. 2016, 76, 2105–2114.
- Ren, J.; Ding, L.; Zhang, D.; Shi, G.; Xu, Q.; Shen, S.; Wang, Y.; Wang, T.; Hou, Y. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theranostics 2018, 8, 3932–3948.
- Fang, Z.; Chen, W.; Yuan, Z.; Liu, X.; Jiang, H. LncRNA-MALAT1 contributes to the cisplatin-resistance of lung cancer by upregulating MRP1 and MDR1 via STAT3 activation. Biomed. Pharmacother. 2018, 101, 536–542.
- Lan, W.G.; Xu, D.H.; Xu, C.; Ding, C.L.; Ning, F.L.; Zhou, Y.L.; Ma, L.B.; Liu, C.M.; Han, X. Silencing of long non-coding RNA ANRIL inhibits the development of multidrug resistance in gastric cancer cells. Oncol. Rep. 2016, 36, 263–270.
- Zhu, J.; Zhang, R.; Yang, D.; Li, J.; Yan, X.; Jin, K.; Li, W.; Liu, X.; Zhao, J.; Shang, W.; et al. Knockdown of Long Non-Coding RNA XIST Inhibited Doxorubicin Resistance in Colorectal Cancer by Upregulation of miR-124 and Downregulation of SGK1. Cell Physiol. Biochem. 2018, 51, 113–128.
- Liu, Y.; Wang, J.; Dong, L.; Xia, L.; Zhu, H.; Li, Z.; Yu, X. Long Noncoding RNA HCP5 Regulates Pancreatic Cancer Gemcitabine (GEM) Resistance By Sponging Hsa-miR-214-3p To Target HDGF. Oncol. Ther. 2019, 12, 8207–8216.
- Zhang, X.; Zhao, P.; Wang, C.; Xin, B. SNHG14 enhances gemcitabine resistance by sponging miR-101 to stimulate cell autophagy in pancreatic cancer. Biochem. Biophys. Res. Commun. 2019, 510, 508–514.
- Chang, K.C.; Diermeier, S.D.; Yu, A.T.; Brine, L.D.; Russo, S.; Bhatia, S.; Alsudani, H.; Kostroff, K.; Bhuiya, T.; Brogi, E.; et al. MaTAR25 lncRNA regulates the Tensin1 gene to impact breast cancer progression. Nat. Commun. 2020, 11, 6438.
- Zhang, Y.; Huang, W.; Yuan, Y.; Li, J.; Wu, J.; Yu, J.; He, Y.; Wei, Z.; Zhang, C. Long non-coding RNA H19 promotes colorectal cancer metastasis via binding to hnRNPA2B1. J. Exp. Clin. Cancer Res. 2020, 39, 141.
- Xing, Z.; Lin, A.; Li, C.; Liang, K.; Wang, S.; Liu, Y.; Park, P.K.; Qin, L.; Wei, Y.; Hawke, D.H.; et al. lncRNA directs cooperative epigenetic regulation downstream of chemokine signals. Cell 2014, 159, 1110–1125.
- Leng, X.; Ding, X.; Wang, S.; Fang, T.; Shen, W.; Xia, W.; You, R.; Xu, K.; Yin, R. Long noncoding RNA AFAP1-AS1 is upregulated in NSCLC and associated with lymph node metastasis and poor prognosis. Oncol. Lett. 2018, 16, 727–732.
- Deng, S.J.; Chen, H.Y.; Ye, Z.; Deng, S.C.; Zhu, S.; Zeng, Z.; He, C.; Liu, M.L.; Huang, K.; Zhong, J.X.; et al. Hypoxia-induced LncRNA-BX111 promotes metastasis and progression of pancreatic cancer through regulating ZEB1 transcription. Oncogene 2018, 37, 5811–5828.
- Luo, Y.; Wang, Q.; Teng, L.; Zhang, J.; Song, J.; Bo, W.; Liu, D.; He, Y.; Tan, A. LncRNA DANCR promotes proliferation and metastasis in pancreatic cancer by regulating miRNA-33b. FEBS Open Bio 2020, 10, 18–27.
- Zhang, M.; Zhao, Y.; Zhang, Y.; Wang, D.; Gu, S.; Feng, W.; Peng, W.; Gong, A.; Xu, M. LncRNA UCA1 promotes migration and invasion in pancreatic cancer cells via the Hippo pathway. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 1770–1782.
- Sun, F.; Wu, K.; Yao, Z.; Mu, X.; Zheng, Z.; Sun, M.; Wang, Y.; Liu, Z.; Zhu, Y. Long Noncoding RNA PVT1 Promotes Prostate Cancer Metastasis by Increasing NOP2 Expression via Targeting Tumor Suppressor MicroRNAs. OncoTargets Ther. 2020, 13, 6755–6765.
- Yang, M.H.; Hu, Z.Y.; Xu, C.; Xie, L.Y.; Wang, X.Y.; Chen, S.Y.; Li, Z.G. MALAT1 promotes colorectal cancer cell proliferation/migration/invasion via PRKA kinase anchor protein 9. Biochim. Biophys. Acta 2015, 1852, 166–174.
- Gupta, R.A.; Shah, N.; Wang, K.C.; Kim, J.; Horlings, H.M.; Wong, D.J.; Tsai, M.C.; Hung, T.; Argani, P.; Rinn, J.L.; et al. Long non-coding RNA HOTAIR reprograms chromatin state to promote cancer metastasis. Nature 2010, 464, 1071–1076.
- Ling, H.; Spizzo, R.; Atlasi, Y.; Nicoloso, M.; Shimizu, M.; Redis, R.S.; Nishida, N.; Gafà, R.; Song, J.; Guo, Z.; et al. CCAT2, a novel noncoding RNA mapping to 8q24, underlies metastatic progression and chromosomal instability in colon cancer. Genome Res. 2013, 23, 1446–1461.
- Tee, A.E.; Liu, B.; Song, R.; Li, J.; Pasquier, E.; Cheung, B.B.; Jiang, C.; Marshall, G.M.; Haber, M.; Norris, M.D.; et al. The long noncoding RNA MALAT1 promotes tumor-driven angiogenesis by up-regulating pro-angiogenic gene expression. Oncotarget 2016, 7, 8663–8675.
- Fu, W.M.; Lu, Y.F.; Hu, B.G.; Liang, W.C.; Zhu, X.; Yang, H.D.; Li, G.; Zhang, J.F. Long noncoding RNA Hotair mediated angiogenesis in nasopharyngeal carcinoma by direct and indirect signaling pathways. Oncotarget 2016, 7, 4712–4723.
- Yuan, S.X.; Yang, F.; Yang, Y.; Tao, Q.F.; Zhang, J.; Huang, G.; Yang, Y.; Wang, R.Y.; Yang, S.; Huo, X.S.; et al. Long noncoding RNA associated with microvascular invasion in hepatocellular carcinoma promotes angiogenesis and serves as a predictor for hepatocellular carcinoma patients’ poor recurrence-free survival after hepatectomy. Hepatology 2012, 56, 2231–2241.
- Zhang, J.X.; Chen, Z.H.; Chen, D.L.; Tian, X.P.; Wang, C.Y.; Zhou, Z.W.; Gao, Y.; Xu, Y.; Chen, C.; Zheng, Z.S.; et al. LINC01410-miR-532-NCF2-NF-kB feedback loop promotes gastric cancer angiogenesis and metastasis. Oncogene 2018, 37, 2660–2675.
- Cai, H.; Liu, X.; Zheng, J.; Xue, Y.; Ma, J.; Li, Z.; Xi, Z.; Li, Z.; Bao, M.; Liu, Y. Long non-coding RNA taurine upregulated 1 enhances tumor-induced angiogenesis through inhibiting microRNA-299 in human glioblastoma. Oncogene 2017, 36, 318–331.
- Zhao, J.; Du, P.; Cui, P.; Qin, Y.; Hu, C.; Wu, J.; Zhou, Z.; Zhang, W.; Qin, L.; Huang, G. LncRNA PVT1 promotes angiogenesis via activating the STAT3/VEGFA axis in gastric cancer. Oncogene 2018, 37, 4094–4109.
- Lin, X.; Yang, F.; Qi, X.; Li, Q.; Wang, D.; Yi, T.; Yin, R.; Zhao, X.; Zhong, X.; Bian, C. LncRNA DANCR promotes tumor growth and angiogenesis in ovarian cancer through direct targeting of miR-145. Mol. Carcinog. 2019, 58, 2286–2296.
- Guo, Z.; Wang, X.; Yang, Y.; Chen, W.; Zhang, K.; Teng, B.; Huang, C.; Zhao, Q.; Qiu, Z. Hypoxic Tumor-Derived Exosomal Long Noncoding RNA UCA1 Promotes Angiogenesis via miR-96-5p/AMOTL2 in Pancreatic Cancer. Mol. Ther. Nucl. Acids 2020, 22, 179–195.
- Jiang, X.; Yan, Y.; Hu, M.; Chen, X.; Wang, Y.; Dai, Y.; Wu, D.; Wang, Y.; Zhuang, Z.; Xia, H. Increased level of H19 long noncoding RNA promotes invasion, angiogenesis, and stemness of glioblastoma cells. J. Neurosurg. 2016, 124, 129–136.
- Zhang, C.; Luo, Y.; Cao, J.; Wang, X.; Miao, Z.; Shao, G. Exosomal lncRNA FAM225A accelerates esophageal squamous cell carcinoma progression and angiogenesis via sponging miR-206 to upregulate NETO2 and FOXP1 expression. Cancer Med. 2020, 9, 8600–8611.
- Zhang, X.; Sun, Z.; Zou, Y. LncRNA NEAT1 exacerbates non-small cell lung cancer by upregulating EIF4G2 via miR-582-5p sponging. Arch. Biol. Sci. 2020, 72, 243–252.
- Fang, Q.L.; Zhou, J.Y.; Xiong, Y.; Xiong, Y.; Xie, C.R.; Wang, F.Q.; Li, Y.T.; Yin, Z.Y.; Luo, G.H. Long non-coding RNA RP11-284P20.2 promotes cell proliferation and invasion in hepatocellular carcinoma by recruiting EIF3b to induce c-met protein synthesis. Biosci. Rep. 2020, 40, BSR20200297.
- Hu, M.; Yang, J. Down-regulation of lncRNA UCA1 enhances radiosensitivity in prostate cancer by suppressing EIF4G1 expression via sponging miR-331-3p. Cancer Cell Int. 2020, 20, 449.
- Hansji, H.; Leung, E.Y.; Baguley, B.C.; Finlay, G.J.; Cameron-Smith, D.; Figueiredo, V.C.; Askarian-Amiri, M.E. ZFAS1: A long noncoding RNA associated with ribosomes in breast cancer cells. Biol. Direct. 2016, 11, 62.
- Liu, P.Y.; Tee, A.E.; Milazzo, G.; Hannan, K.M.; Maag, J.; Mondal, S.; Atmadibrata, B.; Bartonicek, N.; Peng, H.; Ho, N.; et al. The long noncoding RNA lncNB1 promotes tumorigenesis by interacting with ribosomal protein RPL35. Nat. Commun. 2019, 10, 5026.
- Faghihi, M.A.; Zhang, M.; Huang, J.; Modarresi, F.; Van der Brug, M.P.; Nalls, M.A.; Cookson, M.R.; St-Laurent, G., 3rd; Wahlestedt, C. Evidence for natural antisense transcript-mediated inhibition of microRNA function. Genome Biol. 2010, 11, R56.
- Li, W.; Zhai, L.; Wang, H.; Li, W.; Zhai, L.; Wang, H.; Liu, C.; Zhang, J.; Chen, W.; Wei, Q. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget 2016, 7, 27778–27786.
- Xiao, J.; Lin, L.; Luo, D.; Shi, L.; Chen, W.; Fan, H.; Li, Z.; Ma, X.; Ni, P.; Yang, L.; et al. Long noncoding RNA TRPM2-AS acts as a microRNA sponge of miR-612 to promote gastric cancer progression and radioresistance. Oncogenesis 2020, 9, 29.
- Cao, L.; Zhang, P.; Li, J.; Wu, M. LAST, a c-Myc-inducible long noncoding RNA, cooperates with CNBP to promote CCND1 mRNA stability in human cells. Elife 2017, 6, e30433.
- Song, J.; Ahn, C.; Chun, C.H.; Jin, E.J. A long non-coding RNA, GAS5, plays a critical role in the regulation of miR-21 during osteoarthritis. J. Orthop. Res. 2014, 32, 1628–1635.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
907
Revisions:
2 times
(View History)
Update Date:
16 Jun 2023
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No