 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Eric Hallerman | -- | 4329 | 2023-06-13 19:49:42 | | | |
| 2 | Fanny Huang | -445 word(s) | 3884 | 2023-06-14 07:25:33 | | |
Video Upload Options
Species of the genus Pseudoplatystoma, the long-whiskered catfishes, are important in commercial and recreational fisheries in South America, and some species have become key to regional aquaculture. Some species of the genus are under pressure due to overfishing and the negative impacts of dams.
1. Introduction
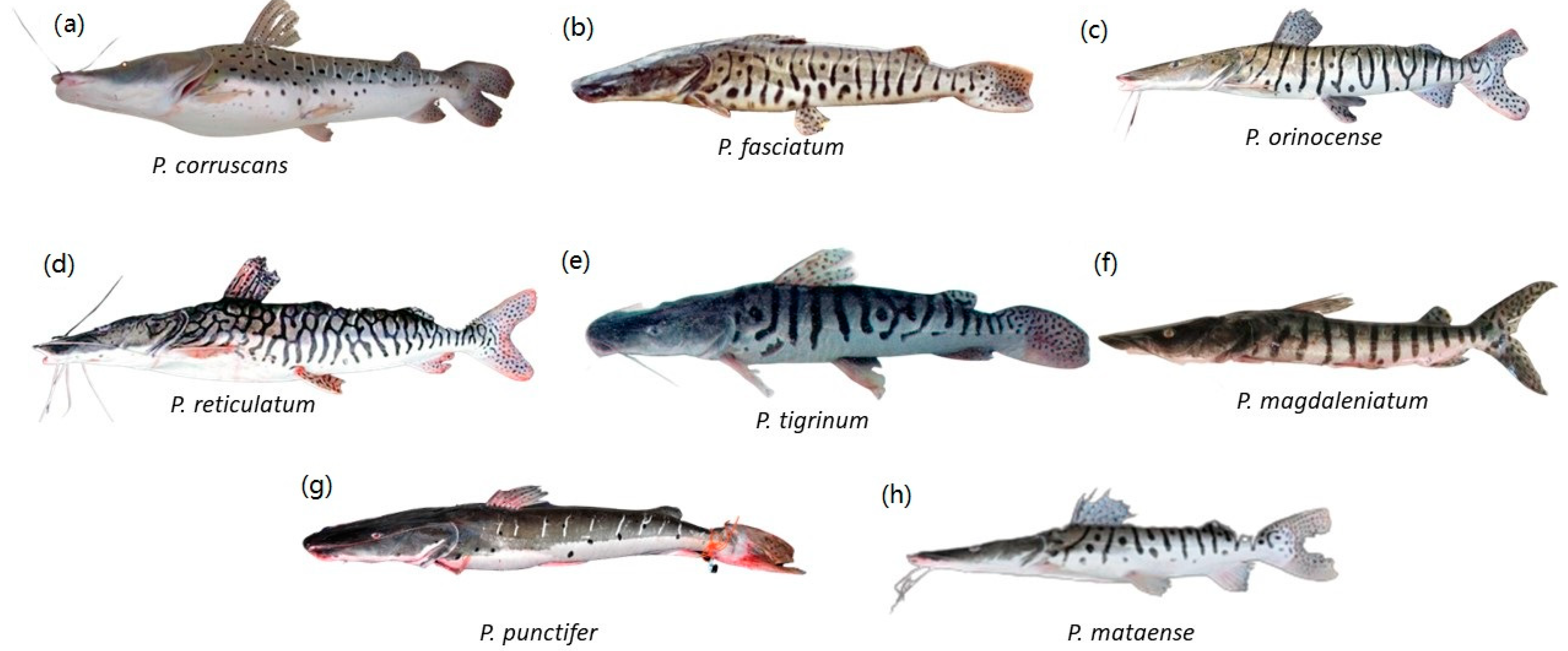
2. What Species Are in the Genus, and Where Are They Distributed?
2.1. Natural History
2.2. Systematics and Taxonomy
2.3. Distributions
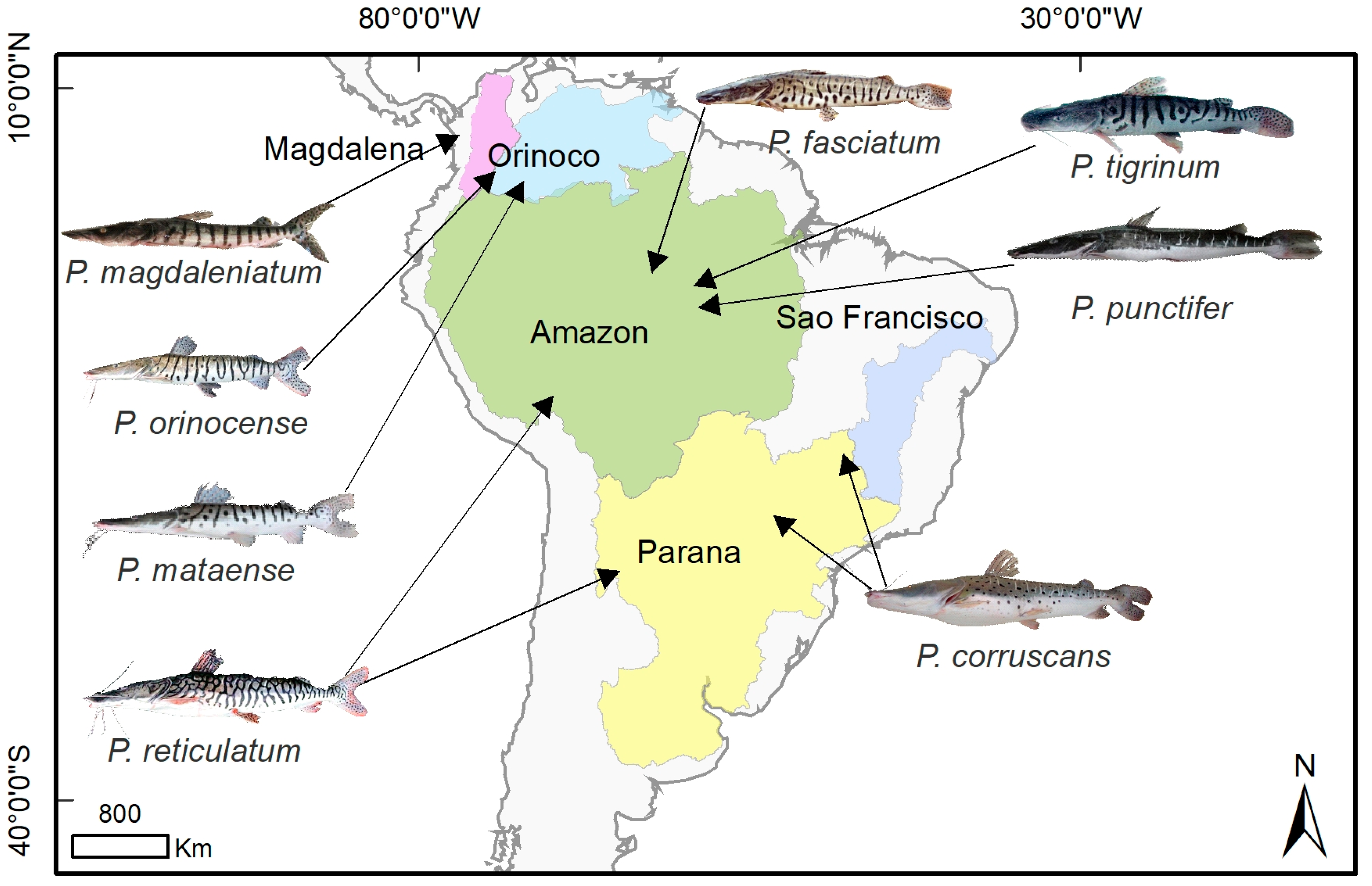
3. What Are the Life Histories and Ecologies of Pseudoplatystoma Species?
3.1. Movement Ecology
3.2. Reproduction
| Species | Size at First Maturation (cm) | Spawning Season | Spawning Type | Oocyte Diameter (mm) | Ref. |
|---|---|---|---|---|---|
| P. corruscans | 62.92 | Rising water | Total | 0.9 | [32][48] |
| P. reticulatum | 57.84 | Rising water | Total | - | [48] |
| P. tigrinum | 69.4 | Rising water | Total | - | [45][53] |
| P. fasciatum | 53 | Rising water | Total | 0.64 | [31][53] |
| P.magdaleniatum | 82 | Rising water | - | 0.9 | [47] |
| P. tigrinum | 69.4 | High water | - | - | [42] |
| P. fasciatum | 53 | Rising water | - | 0.64 | [42] |
3.3. Growth
4. Threats to Conservation of Pseudoplatystoma Species
The main threats to Pseudoplatystoma species are (1) overfishing and (2) alterations in and obstruction of river flow due to the construction of hydropower dams [64]. Other threats—such as industrial, domestic, and agricultural pollution, deforestation, introduced species, hybridization with farmed fish, and genetic erosion—may also affect populations of Pseudoplatystoma species, but in our view, they are not less pressing.
Dams interrupt streamflow and generate longitudinal and lateral hydrological changes in river ecosystems [65][66][67] that can be reflected subsequently in decreased fisheries yields. The most apparent effects of placing dams on rivers are the formation of a new lentic environment upstream from the dam and a tailwater environment downstream from the dam [66]. In general, six effects of dams on fisheries are (1) alteration of flood regime; (2) change in nutrient flow; (3) change in water temperature; (4) disruption of fish migration; (5) change in fish community composition; and (6) change of water chemistry [67]. For Pseudoplatystoma species, the alteration of the flood regime and disruption of migration are particularly important [68].
Alteration of the flood regime disrupts lateral connectivity, as dams cause the permanent inundation of upstream floodplains and restrict the seasonal movement of littoral zones that are part of the flood pulse [66]. Further, downstream of the dam, floodplains are not sufficiently inundated in terms of the depth, duration, and seasonality of flooding, which could result in a significant decline in Pseudoplatystoma populations and fisheries. That is, disruption of floodplain inundation is likely to affect the life cycle of Pseudoplatystoma, including disruption of migration, recruitment, and survival of cohorts. Dams blocking upstream and downstream migration can cause the decline and even extirpation of species that depend on longitudinal movements along the stream during certain phases of their life cycle [65][66]. Migratory fishes require different environments for the main stages of their life cycle: reproduction, juvenile production, growth, and sexual maturation (review in [64]). As a barrier to upstream migration, a dam prevents adults from reaching spawning grounds during the breeding season, resulting in recruitment failure and eventual extirpation of the population above the dam. Downstream migration past the dam may also be difficult or impossible for many fish species. Fish migrating into the reservoir from tributary streams may be unable to find their way to the dam and subsequently pass downstream through discharge structures. In particular, the modification of the downstream river-flow regime by an impoundment can have various adverse effects on Pseudoplatystoma fish species, including loss of stimuli for migration, loss of migration routes and spawning grounds, decreased survival of eggs and juveniles, decreased recruitment to the mature life stage and diminished fisheries production [67].
In addition to obstruction of river flow due to dams, overfishing is especially evident in parts of the São Francisco and Paraná rivers [68], particularly regarding P. corruscans stocks [68][69][70][71]. This species is subject to overfishing in Minas Gerais State, Brazil. P. corruscans harvest in the São Francisco River has manifested indications of collapse [71][72]. In the 1950s, the capture of thousands of P. corruscans with low fishery effort was the norm [73]. More recently, however, the yield of P. corruscans caught in a vital fishing area has declined from 10.3 kg fish/day in 1987 to 0.8 kg fish/day in 1999 [74]. In the Amazon basin, some authors have suggested that P. tigrinum and P. fasciatum are declining in mean catches and captured at small sizes [69][75][76][77].
With continued research, understanding of the biology and ecology of this genus would fill knowledge gaps and contribute to effective fisheries management and conservation of the eight species of the genus Pseudoplatystoma.
References
- Arthington, A.H.; Dulvy, N.K.; Gladstone, W.; Winfield, I.J. Fish Conservation in Freshwater and Marine Realms: Status, Threats and Management. Aquat. Conserv. 2016, 26, 838–857.
- Pelayo-Villamil, P.; Astor Guisande, C.; Vari, R.P.; Manjarr Es-Hern Andez, A.; Garc Ia-Rosell, E.; Gonz Alez-Dacosta, J.; Urgen Heine, J.; Gonz Alez Vilas, L.; Patti, B.; Mar Ia Quinci, E.; et al. Global Diversity Patterns of Freshwater Fishes-Potential Victims of Their Own Success Diversity and Distributions. Divers. Distrib. 2015, 21, 345–356.
- Pinder, A.C.; Britton, J.R.; Harrison, A.J.; Nautiyal, P.; Bower, S.D.; Cooke, S.J.; Lockett, S.; Everard, M.; Katwate, U.; Ranjeet, K.; et al. Mahseer. Fishes of the World: Status, Challenges and Opportunities for Conservation. Rev. Fish Biol. Fish. 2019, 29, 417–452.
- Fricke, R.; Eschmeyer, W.N.; van der Laan, R. Eschmeyer’s Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 16 May 2023).
- Dudgeon, D.; Arthington, A.H.; Gessner, M.O.; Kawabata, Z.I.; Knowler, D.J.; Lévêque, C.; Naiman, R.J.; Prieur-Richard, A.H.; Soto, D.; Stiassny, M.L.J.; et al. Freshwater Biodiversity: Importance, Threats, Status and Conservation Challenges. Biol. Rev. Camb. Philos. Soc. 2006, 81, 163–182.
- Cooke, S.J.; Paukert, C.; Hogan, Z. Endangered River Fish: Factors Hindering Conservation and Restoration. Endanger. Species Res. 2012, 17, 179–191.
- Oliveira, C.; Foresti, A.F.; Hilsdorf, A.A.W.S. Genetics of Neotropical Fish: From Chromosomes to Populations. Fish. Physiol. Biochem. 2009, 35, 81–100.
- Ahmed, S.F.; Kumar, P.S.; Kabir, M.; Zuhara, F.T.; Mehjabin, A.; Tasannum, N.; Hoang, A.T.; Kabir, Z.; Mofijur, M. Threats, Challenges and Sustainable Conservation Strategies for Freshwater Biodiversity. Environ. Res. 2022, 214, 113808.
- Nelson, J.S. Fishes of the World, 3rd ed.; J. Wiley and Sons: New York, NY, USA, 1994; ISBN 0471547131.
- Lundberg, J.G.; Pérez, M.H.S.; Dahdul, W.M.; Aguilera, O.A. The Amazonian Neogene Fish Fauna. In Amazonia, Landscape and Species Evolution: A Look into the Past; Wiley-Blackwell: Hoboken, NJ, USA, 2010; pp. 281–301.
- Nuñez, J.; Dugué, R.; Corcuy Arana, N.; Duponchelle, F.; Renno, J.F.; Raynaud, T.; Hubert, N.; Legendre, M. Induced Breeding and Larval Rearing of Surubí, Pseudoplatystoma fasciatum (Linnaeus, 1766), from the Bolivian Amazon. Aquac. Res. 2008, 39, 764–776.
- Dantas, H.L.; Arcanjo, M.; Neto, S.; Kelly, K.; Oliveira, C.; Severi, W.; Mendonça Diniz, F.; Raquel, M.; Coimbra, M.; Moura, M.R. Genetic Diversity of Captive and Wild Threatened Catfish Pseudoplatystoma corruscans in the São Francisco River. Rev. Fish. Sci. 2013, 21, 237–246.
- Mello, P.H.; Venturieri, R.L.L.; Honji, R.M.; Moreira, R.G. Threatened Fishes of the World: Pseudoplatystoma corruscans (Agassiz, 1829) (Siluriformes: Pimelodidae). Environ. Biol. Fish. 2009, 85, 359–360.
- Armas, M.; Ortega, H.; García-Vasquez, A.; García-Dávila, C.; Vargas, G.; Nuñez, J.; Renno, J.-F.; Duponchelle, F. Age Validation and Contrasted Growth Performances of Pseudoplatystoma punctifer (Siluriformes: Pimelodidae) in Two River Systems of the Western Amazon. Neotrop. Ichthyol. 2022, 20, e210099.
- Santos, R.E.; Pinto-Coelho, R.M.; Fonseca, R.; Simões, N.R.; Zanchi, F.B. The Decline of Fisheries on the Madeira River, Brazil: The High Cost of the Hydroelectric Dams in the Amazon Basin. Fish. Manag. Ecol. 2018, 25, 380–391.
- Agostinho, A.A.; Pelicice, F.M.; Petry, A.C.; Gomes, L.C.; Júlio, H.F. Fish Diversity in the Upper Paraná River Basin: Habitats, Fisheries, Management and Conservation. Aquat. Ecosyst. Health Manag. 2007, 10, 174–186.
- Peluso, L.M.; Mateus, L.; Penha, J.; Bailly, D.; Cassemiro, F.; Suárez, Y.; Fantin-Cruz, I.; Kashiwaqui, E.; Lemes, P. Climate Change Negative Effects on the Neotropical Fishery Resources May Be Exacerbated by Hydroelectric Dams. Sci. Total Environ. 2022, 828, 154485.
- Cunha-Machado, A.S.; Farias, I.P.; Hrbek, T.; Lizarazo, M.D.E.; Alves-Gomes, J.A.; Formiga, K.; Da Silva Batista, J. Genetic Differentiation and Gene Flow of the Amazonian Catfish Pseudoplatystoma punctifer across the Madeira River Rapids Prior to the Construction of Hydroelectric Dams. Hydrobiologia 2022, 849, 29–46.
- Mojica, J.; Valderrama, M.; Jimenez-Segura, L.; Alonso, J.C. Pseudoplatystoma magdaleniatum. IUCN Red List Threatened Species 2016, e.T58439165A61474168.
- Brambilla, L.; Toledo, M.J.; Ibarra, D.A. First Fossil Record of Pseudoplatystoma corruscans (Siluriformes, Pimelodidae) from the Late Pleistocene, Santa Fe, Argentina. J. S. Am. Earth Sci. 2021, 105, 102987.
- Vallone, E.; Ignacio Vezzosi, R.; Luis Cione, A. First Fossil Fish (Teleostei, Siluriformes) from the Late Pleistocene of Santa Fe Province, Argentina Mammal Paleoneurology View Project Cranio-Appendicular Anatomy of American Fossil and Living Deers (Mammalia, Cervidae): Systematic and Phylogenetic Implications of the Most Representative Taxa from the Litoral Region View Project. Alcheringa Austral J. Palaeontol. 2017, 41, 369–377.
- Hilsdorf, A.W.S.; Hallerman, E.M. Genetic Resources of Freshwater Neotropical Fishes. In Genetic Resources of Neotropical Fishes; Springer International Publishing: Cham, Switzerland, 2017; pp. 119–210.
- Cassemiro, F.A.S.; Albert, J.S.; Antonelli, A.; Menegotto, A.; Wüest, R.O.; Cerezer, F.; Coelho, M.T.P.; Reis, R.E.; Tan, M.; Tagliacollo, V.; et al. Landscape Dynamics and Diversification of the Megadiverse South American Freshwater Fish Fauna. Proc. Natl. Acad. Sci. USA 2023, 120, e2211974120.
- Géry, J. The Fresh-Water Fishes of South. Am. Biogeogr. Ecol. S. Am. 1969, 2, 828–848.
- Hubert, N.; Renno, J.F. Historical Biogeography of South American Freshwater Fishes. J. Biogeogr. 2006, 33, 1414–1436.
- Buitrago-Suárez, U.A.; Burr, B.M. Taxonomy of the Catfish Genus Pseudoplatystoma Bleeker (Siluriformes: Pimelodidae) with Recognition of Eight Species. Zootaxa 2007, 1512, 1–38.
- Torrico, J.P.; Hubert, N.; Desmarais, E.; Duponchelle, F.; Nuñez Rodriguez, J.; Montoya-Burgos, J.; Garcia Davila, C.; Carvajal-Vallejos, F.M.; Grajales, A.A.; Bonhomme, F.; et al. Molecular Phylogeny of the Genus Pseudoplatystoma (Bleeker, 1862): Biogeographic and Evolutionary Implications. Mol. Phylogenet. Evol. 2009, 51, 588–594.
- Carvalho-Costa, L.F.; Piorski, N.M.; Willis, S.C.; Galetti, P.M.; Ortí, G. Molecular Systematics of the Neotropical Shovelnose Catfish Genus Pseudoplatystoma Bleeker 1862 Based on Nuclear and MtDNA Markers. Mol. Phylogenet. Evol. 2011, 59, 177–194.
- Lundberg, J.G.; Sullivan, J.P.; Hardman, M. Phylogenetics of the South American Catfish Family Pimelodidae (Teleostei: Siluriformes) Using Nuclear and Mitochondrial Gene Sequences. Proc. Acad. Nat. Sci. Phila. 2011, 161, 153–189.
- García-Dávila, C.; Duponchelle, F.; Castro-Ruiz, D.; Villacorta, J.; Quérouil, S.; Chota-Macuyama, W.; Nú, J.; Römer, U.; Carvajal-Vallejos, F.; Renno, J.-F.; et al. Molecular Identification of a Cryptic Species in the Amazonian Predatory Catfish Genus Pseudoplatystoma (Bleeker, 1962) from Peru. Genetica 2013, 141, 347–358.
- Loubens, G.; Panfili, J. Biologie de Pseudoplatystoma fasciatum et P. tigrinum (Teleostei: Pimelodidae) Dans Le Bassin Du Mamoré (Amazonie Bolivienne). Ichthyol. Explor. Freshw. 2000, 11, 13–34.
- Carolsfeld, J.; Harvey, B.; Ross, C.; Baer, A. (Eds.) Migratory Fishes of South. America; World Fisheries Trust: Victoria, BC, USA, 2003; ISBN 9780444536433.
- Barthem, R.; Goulding, M. Os Bagres Balizadores: Ecologia, Migração e Conservação de Peixes Amazônicos; Tefé: Sociedade Civil Mamirauá; CNPq: Tefe, Brazil, 1997.
- Diaz-Sarmineto, J.A.; Alvarez-León, R. Migratory Fishes of the Colombian Amazon. In Migratory Fishes of South America Chapter 7; Carolsfeld, J., Harvey, B., Ross, C., Baer, A., Eds.; World Fisheries Trust: Victoria, BC, USA, 2003; pp. 303–344.
- Goulding, M.; Lowe-McConnell, R.H. (Eds.) The Fishes and the Forest; University of California Press: Berkeley, CA, USA, 1980.
- Araujo-Lima, C.A.R.M.; Oliveira, E.C. Transport of Larval Fish in the Amazon. J. Fish. Biol. 1998, 53, 297–306.
- Pavlov, S.; Nezdoliy, V.; Urteaga, A.; Sanches, O. Downstream Migration of Juvenile Fishes in the Rivers. J. Ichthyol./Vopr. Ikhtiologii 1995, 35, 227–248.
- Barthem, R.B.; Goulding, M.; Leite, R.G.; Cañas, C.; Forsberg, B.; Venticinque, E.; Petry, P.; Ribeiro, M.L.D.B.; Chuctaya, J.; Mercado, A. Goliath Catfish Spawning in the Far Western Amazon Confirmed by the Distribution of Mature Adults, Drifting Larvae and Migrating Juveniles. Sci. Rep. 2017, 7, 41784.
- Hahn, L.; Martins, E.G.; Nunes, L.D.; da Câmara, L.F.; Machado, L.S.; Garrone-Neto, D. Biotelemetry Reveals Migratory Behaviour of Large Catfish in the Xingu River, Eastern Amazon. Sci. Rep. 2019, 9, 8464.
- Godinho, A.L.; Kynard, B.; Godinho, H.P. Migration and Spawning of Female Surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environ. Biol. Fish. 2007, 80, 421–433.
- Avigliano, E.; Pouilly, M.; Bouchez, J.; Domanico, A.; Sánchez, S.; Llamazares Vegh, S.; Clavijo, C.; Scarabotti, P.; Facetti, J.F.; Caffetti, J.D.; et al. Strontium Isotopes (87Sr/86Sr) Reveal the Life History of Freshwater Migratory Fishes in the La Plata Basin. River Res. Appl. 2020, 36, 1985–2000.
- Mariac, C.; Renno, J.-F.; Garcia-Davila, C.; Vigouroux, Y.; Mejia, E.; Angulo, C.; Ruiz, D.C.; Estivals, G.; Nolorbe, C.; Vasquez, A.G.; et al. Species-Level Ichthyoplankton Dynamics for 97 Fishes in Two Major River Basins of the Amazon Using Quantitative Metabarcoding. Mol. Ecol. 2021, 31, 1627–1648.
- Crepaldi, D.V.; Faria, P.M.C.; Teixeira, E.D.A.; Ribeiro, L.P.; Augusto, Â.; Costa, P.; Chemim De Melo, D.; Paula, A.; Cintra, R.; Prado, S.D.A.; et al. Estructura Histologica de los ovarios de Pseudoplatystoma corruscans (Agassiz, 1829) Pimelodidae, Siluriformes. Boletim do Instituto de Pesca 1996, 23, 203–212.
- Brito, M.F.G.; Bazzoli, N. Reproduction of the Surubim Catfish (Pisces, Pimelodidae) in the São Francisco River, Pirapora Region, Minas Gerais, Brazil. Arq. Bras. Med. Vet. Zootec. 2003, 55, 624–633.
- Pérez, A.; Castillo, O.; Barbarino, A.; Fabré, N. Aspectos Reproductivos Del Bagre Rayado Pseudoplatystoma tigrinum (Siluriformes, Pimelodidae) En La Cuenca Del Río Apure, Venezuela. Zootecnia Trop. 2012, 30, 251–262.
- Jiménez-Segura, L.F.; Palacio, J.; López, R. Caracteristicas Biologicas del Blanquillo Sorobim cuspicaudas Littmann, Burr y Nass, 2000 y Bagre rayado Pseudoplatystoma magdaleniatum Buitrago-Suarez y Burr 2007 (Siluriformes: Pimelodidae) Relacionadas con su Reproducction en la Cuenca Media del Rio Magdalena, Colombia. Actual. Biol. 2009, 31, 53–66.
- Barzotto, E.; Oliveira, M.; Mateus, L. Reproductive Biology of Pseudoplatystoma corruscans (Spix and Agassiz, 1829) and Pseudoplatystoma reticulatum (Eigenmann and Eigenmann, 1889), Two Species of Fisheries Importance in the Cuiabá River Basin, Brazil. J. Appl. Ichthyol. 2017, 33, 29–36.
- Rizzo, E.; Godinho, H.P. Superfície de Ovos de Peixes Characiformes e Siluriformes. In Águas, Peixes e Pescadores do São Francisco das Minas Gerais; Godinho, H.P., Godinho, A.L., Eds.; PUC Minas: Belo Horizonte, Brazil, 2003; pp. 115–132.
- Reid, J. La Biología de Los Bagres Rayados Pseudoplatystoma fasciatum y P. tigrinum En La Cuenca Del Río Apure, Venezuela. Revista Unellez de Ciencia y Tecnología 1983, 1.1, 13–41.
- Resende, A.; Kawakami, E.; Catella, C.; Lima, F.; Shirley Da, N.; Palmeira, S.; Aparecida, R.; Pereira, C.; De, M.; Lima, S. Biologia Do Curimbatá (Prochilodus lineatus), Pintado (Pseudoplatystoma corruscans) e Cachara (Pseudoplatystoma fasciatum) Na Bacia Hidrográfica Do Rio Miranda. Infoteca.Cnptia.Embrapa.br, 1995; 75p, (EMBRAPA-CPAP. Boletim de Pesquisa, 02).
- Padilla Pérez, P.P.; Alcántara Bocanegra, F.; Orbe, R.I. Reproducción Inducida de La Doncella Pseudoplatystoma fasciatum y Desarrollo Embrionario-Larval. Folia amazónica 2001, 12, 141–154.
- Garcia, V.A.; Montreuil, V. Rodriguez R Talla de Primera Maduracion y Epoca de Desove de La “Doncella”, Pseudoplatystoma fasciatum (Linnaeus), y Del “Tigre Zungaro”, Pseudoplatystoma tigrinum (Valenciennes), En La Amazonia Peruana. Boletin del Museo Para Emilio Goeldi Séria Zoologia 2001, 17, 3–13.
- Núñez, J.; Castro, D.; Fernández, C.; Dugué, R.; Chu-Koo, F.; Duponchelle, F.; García, C.; Renno, J.F. Hatching Rate and Larval Growth Variations in Pseudoplatystoma punctifer: Maternal and Paternal Effects. Aquac. Res. 2011, 42, 764–775.
- Andrade, F.F.; Lima, A.F.; Assumpção, L.; Makrakis, S.; Kasai, R.I.D.; Makrakis, M.C. Characterization of the Early Development of Pseudoplatystoma reticulatum Eigenmann & Eigenmann, 1889 (Siluriformes: Pimelodidae) from the Paraguay River Basin. Neotrop. Ichthyol. 2016, 14.
- Erard, C.; Keith, P.; Le Bail, P.Y.; Planquette, P. Atlas Des Poissons d’eau Douce de Guyane. Revue d’Écologie (La Terre et La Vie) 2002, 57, 185–187.
- Pérez, A.; Fabré, N.N. Life-History Characteristics of Pseudoplatystoma Metaense (Teleostei: Siluriformes: Pimelodidae) from the Northwestern Orinoco River Basin. Neotrop. Ichthyol. 2018, 16, 160140.
- Hauser, M.; Doria, C.R.C.; Melo, L.R.C.; Santos, A.R.; Ayala, D.M.; Nogueira, L.D.; Amadio, S.; Fabré, N.; Torrente-Vilara, G.; García-Vásquez, Á.; et al. Age and Growth of the Amazonian Migratory Catfish Brachyplatystoma russeauxii in the Madeira River Basin before the Construction of Dams. Neotrop. Ichthyol. 2018, 16, 1127.
- González, S.Á.R.; Mendoza, J.; Arocha, F.; Márquez, A. Edad y Crecimiento Del Bagre Rayado Pseudoplatystoma orinocoense Del Orinoco Medio En Venezuela. Zootec. Trop. 2010, 28, 283–293.
- Cutrim, L.; Batista, V.d.S. Determinação de Idade e Crescimento Do Mapará (Hypophthalmus marginatus) Na Amazônia Central. Acta Amazon. 2005, 35, 85–92.
- Pérez, A.; Fabré, N.N. Seasonal Growth and Life History of the Catfish Calophysus macropterus (Lichtenstein, 1819) (Siluriformes: Pimelodidae) from the Amazon Floodplain. J. Appl. Ichthyol. 2009, 25, 343–349.
- Arantes, C.C.; Castello, L.; Stewart, D.J.; Cetra, M.; Queiroz, H.L. Population Density, Growth and Reproduction of Arapaima in an Amazonian River-Floodplain. Ecol. Freshw. Fish. 2010, 19, 455–465.
- Henderson, P.A. The Growth of Tropical Fishes. Fish. Physiol. 2005, 21, 85–100.
- Lagler, K.F.; Kapetsky, J.M.; Stewart, D.J. The Fisheries of the Kafue River Flats, Zambia, in Relation to the Kafue Gorge Dam; University of Michigan, School of Natural Resources: Ann Arbor, MI, USA, 1971.
- Duponchelle, F.; Isaac, V.J.; Doria, C.; Van Damme, P.A.; Herrera-R, G.A.; Anderson, E.P.; Cruz, R.E.A.; Hauser, M.; Hermann, T.W.; Agudelo, E.; et al. Conservation of Migratory Fishes in the Amazon Basin. Aquat. Conserv. 2021, 31, 1087–1105. https://doi.org/10.1002/aqc.3550.
- Agostinho, A.A.; Gomes, L.C. Dams and the Fish Fauna of the Neotropical Region: Impacts and Management Related to Diversity and Fisheries. Braz. J. Biol. 2008, 68, 1119–1132.
- Poff, L.N.; Hart, D.D. How Dams Vary and Why It Matters for the Emerging Science of Dam Removal. Bioscience 2002, 52, 659–668.
- Jackson, D.; Marmulla, G.; Dams, Fish and Fisheries. Opportunities, Challenges and Conflict Resolution. FAO Fish. Tech. Paper 2001, 419, 166.
- Carolsfeld, J.; Harvey, B.; Ross, C.; Baer, A. (Eds.) Migratory Fishes of South. America; World Fisheries Trust: Victoria, BC, USA, 2003; ISBN 9780444536433.
- Sato, Y. Reprodução de Peixes Da Bacia Do Rio São Francisco: Indução e Caracterização de Padrões; 1999. 179f. Tese (Doutorado). Centro de Ciências Biológicas e da Saúde, Universidade Federal de São Carlos, São Carlos, SP.
- Sato, Y.; Godinho, H.P. Migratory Fishes of the São Francisco River. In Migrotory Fish of South America; Carolsfeld, J., Harvey, B., Ross, C., Eds.; World Fisheries Trust: Victoria, BC, USA, 2003; pp. 195–232.
- Godinho, H.P.; Miranda, M.O.T.; Godinho, A.L.; Santos, J.E. Pesca e Biologia Do Surubim Pseudoplatystoma Coruscans No Rio São Francisco; Puc Minas: Belo Horizonte, Brazil, 1997.
- Godinho, A.L.; Godinho, H.P. Breve Visão Do São Francisco. Belo Horizonte: PUC Minas 468 (2003): 15-23. In Águas, Peixes e Pescadores do São Francisco das Minas Gerais; PUC Minas: Belo Horizonte, Brazil, 2003; Volume 468, pp. 15–23.
- Menezes, R.S. Pesca e Piscicultura No Vale Do São Francisco. Boletim da Secretaria da Agricultura Indústria e Comércio do Estado de Pernambuco 1956, 23, 43–105.
- Godinho, A.L.; Kynard, B.; Godinho, H.P. Migration and Spawning of Female Surubim (Pseudoplatystoma corruscans, Pimelodidae) in the São Francisco River, Brazil. Environ. Biol. Fish. 2007, 80, 421–433.
- Isaac, V.; Ruffino, M.; McGrath, D. In Search of a New Approach to Fisheries Management in the Middle Amazon Region. In Fishery Stock Assessment Models; Alaska Sea Grant, University of Alaska Fairbanks: Fairbanks, AK, USA, 1998; pp. 889–902.
- Castello, L.; McGrath, D.G.; Beck, P.S.A. Resource Sustainability in Small-Scale Fisheries in the Lower Amazon Floodplains. Fish. Res. 2011, 110, 356–364. https://doi.org/10.1016/j.fishres.2011.05.002.
- Agudelo, E.A.; Acosta-Santos, G.; Gómez, B.D.; Gil, R.E.; Ajiaco-Martínez; Ramírez-Gil, H. Pseudoplatystoma punctifer (Siluriformes, Pimelodidae). In: Lasso, C. A.; Agudelo Córdoba, E.; Jiménez-Segura, L. F.; H. Ramírez- Gil, H.; Morales-Betancourt, M.; Ajiaco-Martínez, R. E.; Gutiérrez,F. de P.; Usma Oviedo, J.S.; Muñoz Torres, S.E.; Sanabria Ochoa, A.I. (Eds.) I. Catálogo de los Recursos Pesqueros Continentales de Colombia. Serie Editorial Recursos Hidro- biológicos y Pesqueros continentales de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt (IAvH). Bogotá, D. C., Colombia, 715 pp. Capitulo 7; 2011; pp. 509–512.

