Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Luis Vitetta | -- | 3162 | 2023-06-02 03:16:49 | | | |
| 2 | Alfred Zheng | Meta information modification | 3162 | 2023-06-02 04:07:47 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Vitetta, L.; Bambling, M.; Strodl, E. Probiotics Influence the Gut–Brain Link. Encyclopedia. Available online: https://encyclopedia.pub/entry/45116 (accessed on 23 July 2026).
Vitetta L, Bambling M, Strodl E. Probiotics Influence the Gut–Brain Link. Encyclopedia. Available at: https://encyclopedia.pub/entry/45116. Accessed July 23, 2026.
Vitetta, Luis, Matthew Bambling, Esben Strodl. "Probiotics Influence the Gut–Brain Link" Encyclopedia, https://encyclopedia.pub/entry/45116 (accessed July 23, 2026).
Vitetta, L., Bambling, M., & Strodl, E. (2023, June 02). Probiotics Influence the Gut–Brain Link. In Encyclopedia. https://encyclopedia.pub/entry/45116
Vitetta, Luis, et al. "Probiotics Influence the Gut–Brain Link." Encyclopedia. Web. 02 June, 2023.
Copy Citation
The effect of the intestinal microbiome on the gut–brain axis has received considerable attention, strengthening the evidence that intestinal bacteria influence emotions and behavior. The colonic microbiome is important to health and the pattern of composition and concentration varies extensively in complexity from birth to adulthood. That is, host genetics and environmental factors are complicit in shaping the development of the intestinal microbiome to achieve immunological tolerance and metabolic homeostasis from birth. Given that the intestinal microbiome perseveres to maintain gut homeostasis throughout the life cycle, epigenetic actions may determine the effect on the gut–brain axis and the beneficial outcomes on mood.
probiotics
Lactobacillus
Bifidobacterium
major depression
gut dysbiosis
epigenetics
inflammation
1. Introduction
What has ensued from epidemiological studies is the observation of an existence of an association between psychiatric and gastrointestinal disorders. Patients diagnosed with anxiety disorders or major depressive disorders (MDD), often also have reported intestinal complaints [1]. In addition to this, patients diagnosed with intestinal disorders such as ulcerative colitis, Crohn’s disease, and irritable bowel syndrome (IBS), often present with a picture of co-morbidity with anxiety issues and/or depression [1]. Such epidemiological outcomes have introduced the concept of the gut–brain axis that informs researchers of the extensive bidirectional communication network that links the intestinal tract with central cognitive and emotional centers within the central nervous system (Figure 1).
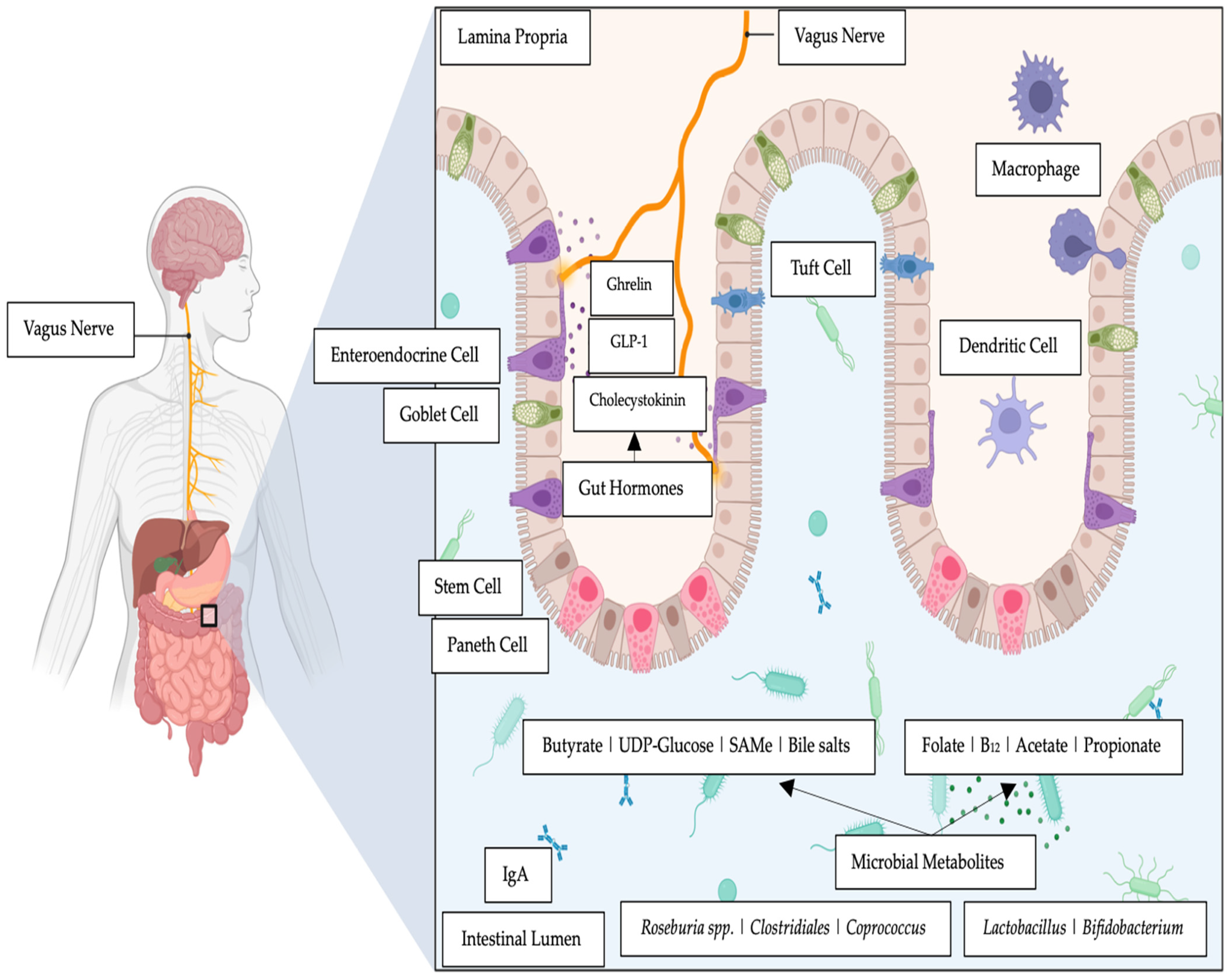
Figure 1. The gut–brain axis and examples of intestinal metabolites elaborated by relevant microbial species and gut hormones.
Reports on the gut–brain axis apprise the bidirectional molecular interactions that exist between the intestines, the resident intestinal microbiome, and the enteric and central nervous systems [2]. Investigations of the intestinal microbiome have progressed our understanding of the gut–brain axis’ role in mood disorders [3]. Numerous studies on laboratory animals and humans have reported that an imbalanced intestinal microbiome (i.e., dysbiosis) is linked to mood disorders such as anxiety and depression [4][5]. Moreover, intestinal dysbiosis-associated depression can present with gut inflammatory sequalae which can significantly exacerbate excessive adverse gut mucosal barrier integrity (i.e., a dysbiotic intestinal barrier), further disrupting homeostasis [5].
The dominant phyla in a typical intestinal microbiome include the Bacteroidetes and Firmicutes that account for approximately 90% of the intestinal bacterial community, and other subdominant phyla that include Actinobacteria, Proteobacteria, and Verucomicrobia [6]. Research activities are accumulating evidence that the intestinal microbiota may influence brain activity and behavior via neural and humoral pathways [7]. It is further reported that gut bacteria may have translational applications in the treatment of neuropsychiatric disorders such as MDD [8]. Support for these contentions comes from several animal studies that overall conclude that intestinal bacteria could have a significant impact on the neurobiological features of mood dispositions and depression [9][10][11].
The established consensus from clinical, epidemiological, and immunological evidence suggests that intestinal bacteria can extensively and profoundly influence how the gut and the brain respond and influence mental states, emotional regulation, neuromuscular function, and the regulation of the hypothalamus–pituitary–adrenal axis (HPA) [1]. The effect of probiotics on psychiatric symptoms and central nervous system function in human health remain contentious as reported in a recent meta-analysis [12]. Notwithstanding, the meta-analysis [12] concluded that developing next-generation probiotics to improve psychiatric conditions and other CNS functions is a worthwhile endeavor with early reports showing clinical efficacy. These promising signs of clinical effectiveness may be due to the epigenetic effects exerted on gut metabolites (e.g., B group vitamins, SCFAs, orotate metabolites) elaborated in the intestines by probiotic and commensal bacteria that can contribute to beneficial epigenetic changes [13].
Research continues to elucidate mechanisms of action to explain both the direct and indirect effects of intestinal bacteria on emotional and cognitive centers of the brain [7] that specifically link gut bacteria compositional changes to brain function and behavior. The intestines harbor a complex and diverse community of bacteria with a concentration gradient from the proximal to distal direction with most of the metabolic activity (i.e., host bacterial nutrient, xenobiotic and drug metabolism) restricted to the colon [14]. Reports show that the intestinal bacteria can elaborate a plethora of neurotransmitters including dopamine, gamma-amino butyrate, norepinephrine, acetylcholine, and serotonin that can affect mood [15] as well as short-chain fatty acids (SCFAs) (e.g., butyrate) and hormones such as cortisol and chemicals that modulate the immune system (e.g., quinolinic acid) [15]. For example, the Flemish Gut Flora Project observed a depletion of butyrate-producing bacteria (i.e., Coprococcus and Dialister) in individuals diagnosed with depression [15]. Whilst gut bacteria that were butyrate producers such as Faecalibacterium and Coprococcus bacteria were consistently associated with higher quality of life indicators [15].
Investigators agree that a healthy gut is related to a normal functioning central nervous system (CNS) [2][16][17][18]. It has been posited that gut dysbiosis exacerbates taxonomic changes, as observed in patients with MDD [18][19]. These changes have been associated with bacterial proinflammatory activity, impaired intestinal barrier integrity, and neurotransmitter production, with adverse effects on carbohydrates, tryptophane and glutamate metabolic pathways, and reduced levels of SCFAs [18]. An early report on gut dysbiosis from clinical sampling showed that the gut bacterial compositions of patients with major depressive disorders (MDD) versus those of healthy controls exhibited differences that were characterized by significant changes in the relative abundance of Firmicutes, Actinobacteria and Bacteroidetes phyla [20].
An intestinal microbiota–inflammasome hypothesis of MDD has been advanced that proposes the occurrence of pathological shifts in the composition of the intestinal microbiota (i.e., dysbiosis) that are exacerbated by stress and gut conditions that result in the upregulation of pro-inflammatory pathways [21]. Early studies support such links between gut resident bacteria and mood disorders. A study has reported an inflammasome-signaling pathway that affects anxiety and depressive mood disorders [22]. In a murine model study that induced gut inflammation with the administration of caspace-1 (an evolutionarily conserved enzyme that influences pro-inflammatory cytokines), the authors showed that when the animals were stressed and treated with an antibiotic (i.e., minocycline) the intestinal microbiota displayed alterations with increases in relative abundances of Akkermansia spp. and Blautia spp. These relative changes were compatible with the beneficial effects of attenuated inflammation and re-equilibrized gut microbiota. Moreover, the increase in Lachnospiracea abundance was consistent with changes in the microbiota in terms of caspace-1 deficiency [22]. Along the similar mechanistic studies, others have suggested that the emerging evidence further supports the presence of a microbiota–intestinal–inflammasome–brain axis, in which enteric bacteria modulate, via NLRP3 signaling, inflammatory pathways that, in turn, contribute and influence brain homeostasis [23].
2. Molecules That Influence the Gut–Brain Link
It is known that the intestinal microbiota affects many physiological processes, such as cell proliferation, epithelial barrier function, and immune responses, and these processes have a direct link to stress and mood dispositions [24]. The role of probiotics in the management of mood appears complex, with the evidence being stronger in depressed populations than in healthy populations [25]. However, this complexity might be explained by the molecules displaying epigenetic influence that have been posited to play important roles in mediating the health-promoting attributes of commensal and probiotic bacteria [13].
2.1. Reactive Oxygen Species
Reactive oxygen species (ROS) such as superoxide and hydrogen peroxide [26] are the active intermediates and regulators of major epigenetic processes such as DNA methylation and histone acetylation reactions [27]. Early reports show that specific taxa of intestinal bacteria can induce the rapid and transient enzymatic production of reactive oxygen species (ROS) within enterocytes [28]. This report showed that redox signaling can be triggered by microbially generated ROS. Mechanistical formation was reported to be a response to microbial signals activated via formyl peptide receptors and the gut epithelial NADPH oxidase1 (Nox1) [28]. There are different strains of commensal gut bacteria that can elicit markedly different levels of ROS from cells that they adhere to. Therefore, the idea has been presented that there is favorable redox potential that the gut microbiota (i.e., the tendency and capacity of the microbiota to gain electrons) and the host have which influences the homeostasis of the intestinal barrier [29]. Studies that administered antibiotics demonstrated how altering the diversity of the gut microbiome through antibiotic-induced changes can disrupt redox dynamics in the intestines [29]. It has also been posited that the redox potential in the intestines can also be modulated by the brain and the CNS via the vagal cholinergic anti-inflammatory pathway [30][31]. For example, Lactobacillus species from the gut have been reported to specifically generate potent inducers of ROS generation in cultured cells as well as in vivo. Moreover, researchers have reported that numerous bacteria exhibit some ability to alter the intracellular oxido-reductase environment [32].
Both DNA methylation and histone acetylation are nucleophilic processes and therefore ROS signaling through typical free-radical processes can take part in epigenetic processes via the reactions of nucleophilic substitution, and these reactions represent the first explanation of their role in epigenetic processes [33][34].
Yan et al., (2007) reported that there are soluble factors that are produced by different strains of Lactobacillus that can have beneficial effects in in vivo inflammatory models [35], thereby presenting data that expand our understanding of the intestinal microbiome’s activity. These results indicate that there are ROS-stimulating bacteria that possess effective specific membrane components and or secreted factors that activate cellular ROS production to maintain homeostasis [36]. Furthermore, it has been reported that enteric commensal bacteria produced the rapid generation of ROS thereby negotiating acceptance from the intestinal epithelia [37].
2.2. B Group Vitamins
The metabolic pathways of B vitamins (i.e., vitamin B6 and B9 and B12) have continuously been implicated in DNA methylations [27], where deficits of these vitamins have been thought to contribute to cognitive decline through increased homocysteine levels with subsequent adverse effects on oxido-reductase redox signaling [38]. In a recent longitudinal study with 2533 participants [39], it was concluded that higher levels of dietary folate at the baseline predicted a better cognitive reserve. Alternatively, decreased serum levels of B vitamins were posited to contribute to cognitive impairment, and this was in turn reported to affect the level of methylation of specific redox-related genes [39]. In a recent systematic review, it was concluded that in at-risk populations with poor mood dispositions, vitamin B group supplementations may provide a significant health benefit [40].
Furthermore, it has been reported that Streptococcus thermophiles, Lactobacillus and Bifidobacterium genera synthesize folic acid to increase DNA methylation and mRNA N6-methyladenosine in the gut, to thereby maintain normal intestinal homeostasis [41][42]. These data support the position that epigenetics presents a new frontier for the beneficial health effects that probiotics portray beyond the data reported by interventional studies [13]. The underlying molecular mechanisms associated with the health-promoting effects of probiotics are linked to effector molecules produced by probiotics that influence specific genes and even individual nucleotides [43].
2.3. Short-Chain Fatty Acids (SCFAs)
Although the underlying mechanisms remain unclear, SCFAs, the chief colon metabolites produced via commensal bacterial fermentation of dietary fibers and resistant starch, are postulated to have key roles in mediating the relationship between the intestines and DNA methylation [44][45]. For example, the SCFA butyrate has been reported to improve adverse mood dispositions [46].
The genome of some species of the Bifidobacterium genera from the human gut and those also of animal origin demonstrate a high presence of genes involved in the metabolism of complex oligosaccharides [47]. Other resident bacteria of the colon are also able to degrade inulin-type fructans, as is the case for Lactobacilli, Bacteroides, certain enterobacteria, and butyrate producers. Bacterial cross-feeding mechanisms in the colon form the basis of overall butyrate production, a functional characteristic of several colon bacteria. In addition, the specificity of polysaccharide use by the colon microbiota may determine diet-induced alterations in the microbiota and consequent metabolic effects [48]. Certainly, supplementation with undigested polysaccharides of plant origin is important for the enrichment of the intestinal microbiota with Lactobacillus and Bifidobacterium species, which ferment these compounds into SCFAs [49][50]. Early research has reported that SCFAs, such as butyrate, can have epigenetic effects on the gut [51]. Although the mechanism of action of butyrate is complex, numerous actions involve an epigenetic regulation of gene expression through the inhibition of histone deacetylase [51].
A mechanistic review reported that butyrate [52] and beta-hydroxy-butyrate (b-OHB) [53] are inhibitors of the catalytic activity of Zn2+-dependent histone deacetylases (HDACs). Such inhibition was robustly shown to elicit anti-inflammatory effects in cell culture and rodent model studies. Specifically, SCFA inhibition of HDACs was beneficial in improving neurocognitive and mood disorders. The class I HDAC inhibitor MS-275 prevented depression-like behaviors in mice when subjected to a social stress model [53].
There has been a stream of continuous research that has focused on the role fatty acids have in the neurometabolic pathophysiology of psychiatric disorders [54]. Furthermore, brain phospholipid metabolism and membrane fluidity have been posited to be involved in mood disorders [55]. In a murine model using the forced swim test reported that dietary supplementation with omega-3 fatty acids reduced immobility when given for 30 days [55]. This research also investigated and postulated the antidepressant effect of uridine. Moreover, when both agents were administered together, less of each agent was required for an effective antidepressant outcome.
2.4. Bile Salts, Mood Dispositions and Intestinal Bacteria
People previously reported that the intestinal microbiome can significantly affect and dysregulate the biochemistry of bile salts [56]. A systematic review of gut microbiome variations involved an investigation on patients diagnosed with MDD [57]. From the 17 included studies in the systematic review, 4 found that there was reduced alpha diversity in studies of MDD. Further, intestinal microbiota compositions were clustered separately according to the β diversity between patients and controls in twelve other studies. When comparing patients with MDD versus controls, it was concluded that there was an increase in the relative abundance of the bacterial genera Eggerthella, Atopobium, and Bifidobacterium, and that there was a decrease in the relative abundance of Faecalibacterium in MDD patients compared to the controls [57].
A recent study with a chronic unpredictable mild-stress-induced mouse model showed that increased secondary bile acid levels in the feces positively correlated with Ruminococcaceae, Ruminococcus, and Clostridia abundances [58]. The authors concluded that an increased abundance of bacterial species from the family Ruminococcaceae responded to chronic stress with an increased level of biosynthesis of deoxycholic acid, an unconjugated secondary bile acid in the intestines.
Moreover, a recent study on patients diagnosed with MDD compared to healthy controls reported that there exists a disturbance in the intestinal microbiome of patients diagnosed with MDD [59]. It was also reported that gut dysbiosis correlated with a disturbance in bile salt metabolism [59]. Specifically, bile salt analysis showed that the amount of 23-nordeoxycholic acid in patients with MDD was significantly higher than that in healthy controls, whereas the amounts of 9 taurolithocholic acid, glycolithocholic acid, and lithocholic acid 3-sulfate were significantly lower. Moreover, the intestinal microbiome showed positive associations between Turicibacteraceae, Turicibacterales, and Turicibacter with taurolithocholic acid, glycolithocholic acid and glycodeoxycholic acid [59].
2.5. Choline, Trimethylamine-N-Oxide and Bacteria
Dietary choline is an important nutrient for the production of acetylcholine, a neurotransmitter that plays an important role in regulating memory, mood, and intelligence [60] as well as in the synthesis of methionine, a methyl donor of s-adenosyl methionine (SAMe) [61]. Hence, choline is a metabolite that has been reported to be a methyl donor that influences epigenetic regulation [61].
There appears to be a paradoxical view in terms of the levels of choline relative to mood that are found in different body compartments. In a review analysis of clinical studies, it was reported that high levels of choline concentration in the frontal lobe were associated with depression both in those who responded to treatment and those who did not, following treatment with psychiatric medications, repetitive transcranial magnetic stimulation, or electroconvulsive therapy [62]. In contrast, in a large observational study, it was concluded that the lowest choline quintile level in blood was significantly associated with high anxiety levels but not depression [63]. Furthermore, Romano and colleagues (2017) reported that a gut bacteria-induced reduction in choline reduced methyl-donor availability and influenced global DNA methylation patterns with alterations in behavior.
SAMe is involved in one-carbon metabolism and epigenetic modifications of DNA [64]. The administration of SAMe has given positive data in the management of MDD when administered as a monotherapy or as an adjunct to antidepressant pharmacotherapy [65]. In a recent review, the therapeutic roles of SAMe and probiotics in depression was highlighted [66]. SAMe and probiotics may have synergistic effects in terms of positively influencing mood dispositions.
Intestinal bacterial choline metabolism generates trimethylamine oxide (TMAO) and regulates epigenetic mechanisms [67][68]. A recent study reported that elevated TMAO levels immediately after an AMI could reflect severe stress in PTSD-vulnerable patients [69], making it a plausible biological correlate for severe stress that is associated with vulnerability to PTSD.
2.6. Orotate, Uridine and Bacteria
Orotate is an organic compound present in dairy products as well as in the milk of ruminants whereby it has critical value in basal processes of the organism. It is primarily an intermediate for pyrimidine metabolism, a precursor of uridine-monophosphate, with important roles in DNA and RNA synthesis and anti-inflammatory activity through uridine-monophosphate formation [70]. The group conducted two pilot studies that investigated the administration of magnesium orotate in combination with SSRIs and of probiotics and SSRIs, respectively [71][72], to patients diagnosed with major depression not responsive to the SSRIs.
The first pilot study [71] investigated the plausible beneficial effects of S-adenosylmethionine (SAMe) and magnesium orotate in patients with major depression. The study was conducted with patients during a 15-week period beginning with SAMe administration. At the end of week 15, non-responsive patients with depression were subjected to an 8-week-long administration of a combined intervention of SSRIs and magnesium orotate. The results showed that there was good compliance with no participant drop-outs and a lack of adverse effects. People posited at the time that the intestinal microbiome was complicit in both the suboptimal response to psychotropic therapy and in improving the SAMe and magnesium orotate response via metabolism and absorption functions.
The second pilot study [72] recruited patients with treatment-resistant symptoms of depression. The study recruited 12 patients who were administered, over an 8-week period, a combination of magnesium orotate and probiotics as adjuncts to SSRIs. At the end of the 8-week treatment period, all participants had improved depression and anxiety scores, with self-assessment scores revealing increased energy and higher levels of well-being. These results suggest the bidirectional synergic mechanism of the combination of a SSRI, magnesium orotate and probiotics on the gut–brain axis. At the 16-week follow-up after the cessation of the administration of magnesium orotate and probiotics, there was an observed relapse of depression symptoms while the SSRI medications were still being administered [72].
Early in vitro studies demonstrated how in yeast and in extracts from livers of several species, there was an observed conversion of orotate into pyrimidine nucleotides [73]. Previous studies showed that a Lactobacillus species (Lactobacillus bulgaricus 09) for which orotate is a specific growth factor was found to have an enzyme converting orotate to uridine-5-phosphate [74], while an additional Lactobacillus species (Lactobacillus arabinosus) required uracil specifically at a certain phase of its growth cycle [74][75]. It was concluded that these bacterial species which responded nutritionally to either pyrimidine were found to have both the orotate- and uracil-utilizing enzymes [73]. Given the role of uridine derivates such as that of uridine diphosphate (UDP)-glucose 6-dehydrogenase (UGDH) in DNA methylation [76], the findings from our two pilot studies point to specific probiotic bacteria and other intestinal commensals utilizing orotate to elaborate uridine metabolites that link uridine’s role in DNA methylation.
References
- Mayer, E.A. Gut feelings: The emerging biology of gut-brain communication. Nat. Rev. Neurosci. 2011, 12, 453–466.
- Carabotti, M.; Scirocco, A.; Maselli, M.A.; Severi, C. The gut-brain axis: Interactions between enteric microbiota, central and enteric nervous systems. Ann. Gastroenterol. 2015, 28, 203–209.
- Oroojzadeh, P.; Bostanabad, S.Y.; Lotfi, H. Psychobiotics: The Influence of Gut Microbiota on the Gut-Brain Axis in Neurological Disorders. J. Mol. Neurosci. 2022, 72, 1952–1964.
- Belizário, J.E.; Faintuch, J. Microbiome and Gut Dysbiosis. Exp. Suppl. 2018, 109, 459–476.
- Vitetta, L.; Bambling, M.; Alford, H. The gastrointestinal tract microbiome, probiotics, and mood. Inflammopharmacology 2014, 22, 333–339.
- Vaga, S.; Lee, S.; Ji, B.; Andreasson, A.; Talley, N.J.; Agréus, L.; Bidkhori, G.; Kovatcheva-Datchary, P.; Park, J.; Lee, D.; et al. Compositional and functional differences of the mucosal microbiota along the intestine of healthy individuals. Sci. Rep. 2020, 10, 14977.
- Cryan, J.F.; Dinan, T.G. Mind-altering microorganisms: The impact of the gut microbiota on brain and behaviour. Nat. Rev. Neurosci. 2012, 13, 701–712.
- Hsiao, E.Y.; McBride, S.W.; Hsien, S.; Sharon, G.; Hyde, E.R.; McCue, T.; Codelli, J.A.; Chow, J.; Reisman, S.E.; Petrosino, J.F.; et al. Microbiota modulate behavioral and physiological abnormalities associated with neurodevelopmental disorders. Cell 2013, 155, 1451–1463.
- Bailey, M.T.; Dowd, S.E.; Galley, J.D.; Hufnagle, A.R.; Allen, R.G.; Lyte, M. Exposure to a social stressor alters the structure of the intestinal microbiota: Implications for stressor-induced immunomodulation. Brain Behav. Immun. 2011, 25, 397–407.
- Clarke, G.; Grenham, S.; Scully, P.; Fitzgerald, P.; Moloney, R.D.; Shanahan, F.; Dinan, T.G.; Cryan, J.F. The microbiome-gut-brain axis during early life regulates the hippocampal serotonergic system in a sex-dependent manner. Mol. Psychiatry 2013, 18, 666–673.
- El Aidy, S.; Kunze, W.; Bienenstock, J.; Kleerebezem, M. The microbiota and the gut-brain axis: Insights from the temporal and spatial mucosal alterations during colonisation of the germfree mouse intestine. Benef. Microbes 2012, 3, 251–259.
- Le Morvan de Sequeira, C.; Hengstberger, C.; Enck, P.; Mack, I. Effect of Probiotics on Psychiatric Symptoms and Central Nervous System Functions in Human Health and Disease: A Systematic Review and Meta-Analysis. Nutrients 2022, 14, 621.
- Morovic, W.; Budinoff, C.R. Epigenetics: A New Frontier in Probiotic Research. Trends Microbiol. 2021, 29, 117–126.
- Olofsson, L.E.; Bäckhed, F. The Metabolic Role and Therapeutic Potential of the Microbiome. Endocr. Rev. 2022, 43, 907–926.
- Valles-Colomer, M.; Falony, G.; Darzi, Y.; Tigchelaar, E.F.; Wang, J.; Tito, R.Y.; Schiweck, C.; Kurilshikov, A.; Joossens, M.; Wijmenga, C.; et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat. Microbiol. 2019, 4, 623–632.
- Zhou, L.; Foster, J.A. Psychobiotics and the gut-brain axis: In the pursuit of happiness. Neuropsychiatr. Dis. Treat. 2015, 11, 715–723.
- Góralczyk-Bińkowska, A.; Szmajda-Krygier, D.; Kozłowska, E. The Microbiota-Gut-Brain Axis in Psychiatric Disorders. Int. J. Mol. Sci. 2022, 23, 11245.
- Łoniewski, I.; Misera, A.; Skonieczna-Żydecka, K.; Kaczmarczyk, M.; Kaźmierczak-Siedlecka, K.; Misiak, B.; Marlicz, W.; Samochowiec, J. Major Depressive Disorder and gut microbiota—Association not causation. A scoping review. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110111.
- Sherwin, E.; Rea, K.; Dinan, T.G.; Cryan, J.F. A gut (microbiome) feeling about the brain. Curr. Opin. Gastroenterol. 2016, 32, 96–102.
- Zheng, P.; Zeng, B.; Zhou, C.; Liu, M.; Fang, Z.; Xu, X.; Zeng, L.; Chen, J.; Fan, S.; Du, X.; et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol. Psychiatry 2016, 21, 786–796.
- Inserra, A.; Rogers, G.B.; Licinio, J.; Wong, M.L. The Microbiota-Inflammasome Hypothesis of Major Depression. Bioessays 2018, 40, e1800027.
- Wong, M.L.; Inserra, A.; Lewis, M.D.; Mastronardi, C.A.; Leong, L.; Choo, J.; Kentish, S.; Xie, P.; Morrison, M.; Wesselingh, S.L.; et al. Inflammasome signaling affects anxiety- and depressive-like behavior and gut microbiome composition. Mol. Psychiatry 2016, 21, 797–805.
- Pellegrini, C.; Antonioli, L.; Calderone, V.; Colucci, R.; Fornai, M.; Blandizzi, C. Microbiota-gut-brain axis in health and disease: Is NLRP3 inflammasome at the crossroads of microbiota-gut-brain communications? Prog. Neurobiol. 2020, 191, 101806.
- Madison, A.; Kiecolt-Glaser, J.K. Stress, depression, diet, and the gut microbiota: Human-bacteria interactions at the core of psychoneuroimmunology and nutrition. Curr. Opin. Behav. Sci. 2019, 28, 105–110.
- Ng, Q.X.; Peters, C.; Ho, C.Y.X.; Lim, D.Y.; Yeo, W.S. A meta-analysis of the use of probiotics to alleviate depressive symptoms. J. Affect. Disord. 2018, 228, 13–19.
- Linnane, A.W.; Kios, M.; Vitetta, L. Healthy aging: Regulation of the metabolome by cellular redox modulation and prooxidant signaling systems: The essential roles of superoxide anion and hydrogen peroxide. Biogerontology 2007, 8, 445–467.
- Coppedè, F. One-carbon epigenetics and redox biology of neurodegeneration. Free Radic. Biol. Med. 2021, 170, 19–33.
- Neish, A.S. Redox signaling mediated by the gut microbiota. Free Radic. Res. 2013, 47, 950–957.
- Reese, A.T.; Cho, E.H.; Klitzman, B.; Nichols, S.P.; Wisniewski, N.A.; Villa, M.M.; Durand, H.K.; Jiang, S.; Midani, F.S.; Nimmagadda, S.N.; et al. Antibiotic-induced changes in the microbiota disrupt redox dynamics in the gut. Elife 2018, 7, e35987.
- Jones, R.M.; Mercante, J.W.; Neish, A.S. Reactive oxygen production induced by the gut microbiota: Pharmacotherapeutic implications. Curr. Med. Chem. 2012, 19, 1519–1529.
- Bonaz, B.; Bazin, T.; Pellissier, S. The Vagus Nerve at the Interface of the Microbiota-Gut-Brain Axis. Front. Neurosci. 2018, 12, 49.
- Lin, P.W.; Myers, L.E.; Ray, L.; Song, S.C.; Nasr, T.R.; Berardinelli, A.J.; Kundu, K.; Murthy, N.; Hansen, J.M.; Neish, A.S. Lactobacillus rhamnosus blocks inflammatory signaling in vivo via reactive oxygen species generation. Free Radic. Biol. Med. 2009, 47, 1205–1211.
- Afanas’ev, I. New nucleophilic mechanisms of ros-dependent epigenetic modifications: Comparison of aging and cancer. Aging Dis. 2014, 5, 52–62.
- Li, J.; Wang, Z.; Li, C.; Song, Y.; Wang, Y.; Bo, H.; Zhang, Y. Impact of Exercise and Aging on Mitochondrial Homeostasis in Skeletal Muscle: Roles of ROS and Epigenetics. Cells 2022, 11, 2086.
- Yan, F.; Cao, H.; Cover, T.L.; Whitehead, R.; Washington, M.K.; Polk, D.B. Soluble proteins produced by probiotic bacteria regulate intestinal epithelial cell survival and growth. Gastroenterology 2007, 132, 562–575.
- Neish, A.S.; Jones, R.M. Redox signaling mediates symbiosis between the gut microbiota and the intestine. Gut Microbes 2014, 5, 250–253.
- Lee, W.J. Bacterial-modulated signaling pathways in gut homeostasis. Sci. Signal. 2008, 1, pe24.
- Gil Martínez, V.; Avedillo Salas, A.; Santander Ballestín, S. Vitamin Supplementation and Dementia: A Systematic Review. Nutrients 2022, 14, 1033.
- An, Y.; Feng, L.; Zhang, X.; Wang, Y.; Wang, Y.; Tao, L.; Qin, Z.; Xiao, R. Dietary intakes and biomarker patterns of folate, vitamin B(6), and vitamin B(12) can be associated with cognitive impairment by hypermethylation of redox-related genes NUDT15 and TXNRD1. Clin. Epigenetics 2019, 11, 139.
- Young, L.M.; Pipingas, A.; White, D.J.; Gauci, S.; Scholey, A. A Systematic Review and Meta-Analysis of B Vitamin Supplementation on Depressive Symptoms, Anxiety, and Stress: Effects on Healthy and ‘At-Risk’ Individuals. Nutrients 2019, 11, 2232.
- Wu, J.; Zhao, Y.; Wang, X.; Kong, L.; Johnston, L.J.; Lu, L.; Ma, X. Dietary nutrients shape gut microbes and intestinal mucosa via epigenetic modifications. Crit. Rev. Food Sci. Nutr. 2022, 62, 783–797.
- Rossi, M.; Amaretti, A.; Raimondi, S. Folate production by probiotic bacteria. Nutrients 2011, 3, 118–134.
- Delgado, S.; Sánchez, B.; Margolles, A.; Ruas-Madiedo, P.; Ruiz, L. Molecules Produced by Probiotics and Intestinal Microorganisms with Immunomodulatory Activity. Nutrients 2020, 12, 391.
- Guo, W.; Zhang, Z.; Li, L.; Liang, X.; Wu, Y.; Wang, X.; Ma, H.; Cheng, J.; Zhang, A.; Tang, P.; et al. Gut microbiota induces DNA methylation via SCFAs predisposing obesity-prone individuals to diabetes. Pharmacol. Res. 2022, 182, 106355.
- Silva, Y.P.; Bernardi, A.; Frozza, R.L. The Role of Short-Chain Fatty Acids From Gut Microbiota in Gut-Brain Communication. Front. Endocrinol. 2020, 11, 25.
- Caspani, G.; Kennedy, S.; Foster, J.A.; Swann, J. Gut microbial metabolites in depression: Understanding the biochemical mechanisms. Microb. Cell 2019, 6, 454–481.
- Bondue, P.; Delcenserie, V. Genome of Bifidobacteria and Carbohydrate Metabolism. Korean J. Food Sci. Anim. Resour. 2015, 35, 1–9.
- De Vuyst, L.; Leroy, F. Cross-feeding between bifidobacteria and butyrate-producing colon bacteria explains bifdobacterial competitiveness, butyrate production, and gas production. Int. J. Food Microbiol. 2011, 149, 73–80.
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107.
- Horiuchi, H.; Kamikado, K.; Aoki, R.; Suganuma, N.; Nishijima, T.; Nakatani, A.; Kimura, I. Bifidobacterium animalis subsp. lactis GCL2505 modulates host energy metabolism via the short-chain fatty acid receptor GPR43. Sci. Rep. 2020, 10, 4158.
- Berni Canani, R.; Di Costanzo, M.; Leone, L. The epigenetic effects of butyrate: Potential therapeutic implications for clinical practice. Clin. Epigenetics 2012, 4, 4.
- Davie, J.R. Inhibition of histone deacetylase activity by butyrate. J. Nutr. 2003, 133, 2485s–2493s.
- Shimazu, T.; Hirschey, M.D.; Newman, J.; He, W.; Shirakawa, K.; Le Moan, N.; Grueter, C.A.; Lim, H.; Saunders, L.R.; Stevens, R.D.; et al. Suppression of oxidative stress by β-hydroxybutyrate, an endogenous histone deacetylase inhibitor. Science 2013, 339, 211–214.
- Mocking, R.J.T.; Assies, J.; Ruhé, H.G.; Schene, A.H. Focus on fatty acids in the neurometabolic pathophysiology of psychiatric disorders. J. Inherit. Metab. Dis. 2018, 41, 597–611.
- Carlezon, W.A., Jr.; Mague, S.D.; Parow, A.M.; Stoll, A.L.; Cohen, B.M.; Renshaw, P.F. Antidepressant-like effects of uridine and omega-3 fatty acids are potentiated by combined treatment in rats. Biol. Psychiatry 2005, 57, 343–350.
- Chen, J.; Thomsen, M.; Vitetta, L. Interaction of gut microbiota with dysregulation of bile acids in the pathogenesis of nonalcoholic fatty liver disease and potential therapeutic implications of probiotics. J. Cell. Biochem. 2019, 120, 2713–2720.
- Knudsen, J.K.; Bundgaard-Nielsen, C.; Hjerrild, S.; Nielsen, R.E.; Leutscher, P.; Sørensen, S. Gut microbiota variations in patients diagnosed with major depressive disorder-A systematic review. Brain Behav. 2021, 11, e02177.
- Qu, Y.; Su, C.; Zhao, Q.; Shi, A.; Zhao, F.; Tang, L.; Xu, D.; Xiang, Z.; Wang, Y.; Wang, Y.; et al. Gut Microbiota-Mediated Elevated Production of Secondary Bile Acids in Chronic Unpredictable Mild Stress. Front. Pharm. 2022, 13, 837543.
- Sun, N.; Zhang, J.; Wang, J.; Liu, Z.; Wang, X.; Kang, P.; Yang, C.; Liu, P.; Zhang, K. Abnormal gut microbiota and bile acids in patients with first-episode major depressive disorder and correlation analysis. Psychiatry Clin. Neurosci. 2022, 76, 321–328.
- Dave, N.; Judd, J.M.; Decker, A.; Winslow, W.; Sarette, P.; Villarreal Espinosa, O.; Tallino, S.; Bartholomew, S.K.; Bilal, A.; Sandler, J.; et al. Dietary choline intake is necessary to prevent systems-wide organ pathology and reduce Alzheimer’s disease hallmarks. Aging Cell 2023, 22, e13775.
- Romano, K.A.; Martinez-Del Campo, A.; Kasahara, K.; Chittim, C.L.; Vivas, E.I.; Amador-Noguez, D.; Balskus, E.P.; Rey, F.E. Metabolic, Epigenetic, and Transgenerational Effects of Gut Bacterial Choline Consumption. Cell Host Microbe 2017, 22, 279–290.e277.
- Riley, C.A.; Renshaw, P.F. Brain choline in major depression: A review of the literature. Psychiatry Res. Neuroimaging 2018, 271, 142–153.
- Bjelland, I.; Tell, G.S.; Vollset, S.E.; Konstantinova, S.; Ueland, P.M. Choline in anxiety and depression: The Hordaland Health Study. Am. J. Clin. Nutr. 2009, 90, 1056–1060.
- Abdul, Q.A.; Yu, B.P.; Chung, H.Y.; Jung, H.A.; Choi, J.S. Epigenetic modifications of gene expression by lifestyle and environment. Arch. Pharm. Res. 2017, 40, 1219–1237.
- Cuomo, A.; Beccarini Crescenzi, B.; Bolognesi, S.; Goracci, A.; Koukouna, D.; Rossi, R.; Fagiolini, A. S-Adenosylmethionine (SAMe) in major depressive disorder (MDD): A clinician-oriented systematic review. Ann. Gen. Psychiatry 2020, 19, 50.
- Ullah, H.; Khan, A.; Rengasamy, K.R.R.; Di Minno, A.; Sacchi, R.; Daglia, M. The Efficacy of S-Adenosyl Methionine and Probiotic Supplementation on Depression: A Synergistic Approach. Nutrients 2022, 14, 2751.
- Begum, N.; Mandhare, A.; Tryphena, K.P.; Srivastava, S.; Shaikh, M.F.; Singh, S.B.; Khatri, D.K. Epigenetics in depression and gut-brain axis: A molecular crosstalk. Front. Aging Neurosci. 2022, 14, 1048333.
- Craciun, S.; Balskus, E.P. Microbial conversion of choline to trimethylamine requires a glycyl radical enzyme. Proc. Natl. Acad. Sci. USA 2012, 109, 21307–21312.
- Baranyi, A.; Enko, D.; von Lewinski, D.; Rothenhäusler, H.B.; Amouzadeh-Ghadikolai, O.; Harpf, H.; Harpf, L.; Traninger, H.; Obermayer-Pietsch, B.; Schweinzer, M.; et al. Assessment of trimethylamine N-oxide (TMAO) as a potential biomarker of severe stress in patients vulnerable to posttraumatic stress disorder (PTSD) after acute myocardial infarction. Eur. J. Psychotraumatol. 2021, 12, 1920201.
- Löffler, M.; Carrey, E.A.; Zameitat, E. Orotate (orotic acid): An essential and versatile molecule. Nucl. Nucl. Nucleic Acids 2016, 35, 566–577.
- Bambling, M.; Parham, S.; Coulson, S.; Vitetta, L. S-adenosylmethionine (SAMe) and Magnesium Orotate as adjunctives to SSRIs in sub-optimal treatment response of depression in adults: A. pilot study. Adv. Integr. Med. 2015, 2, 56–62.
- Bambling, M.; Edwards, S.C.; Hall, S.; Vitetta, L. A combination of probiotics and magnesium orotate attenuate depression in a small SSRI resistant cohort: An intestinal anti-inflammatory response is suggested. Inflammopharmacology 2017, 25, 271–274.
- Arvidson, H.; Eliasson, N.A. Orotic acid as a precusor of pyrimidines in the rat. J. Biol. Chem. 1949, 179, 169–173.
- Wright, L.D.; Miller, C.S.; Skeggs, H.; Huff, J.; Weed, L.L.; Wilson, D. Biological Precursors of the Pyrimidines. J. Am. Chem. Soc. 1951, 73, 1898–1899.
- Wright, L.D.; Miller, C.S. Uracil in growth and pyrimidine nucleotide synthesis of Lactobacillus bulgaricus 09. Proc. Soc. Exp. Biol. Med. 1952, 81, 131–134.
- Oyinlade, O.; Wei, S.; Lal, B.; Laterra, J.; Zhu, H.; Goodwin, C.R.; Wang, S.; Ma, D.; Wan, J.; Xia, S. Targeting UDP-α-D-glucose 6-dehydrogenase inhibits glioblastoma growth and migration. Oncogene 2018, 37, 2615–2629.
More
Information
Subjects:
Microbiology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.3K
Entry Collection:
Gastrointestinal Disease
Revisions:
2 times
(View History)
Update Date:
02 Jun 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No