Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Lorentz Jäntschi | -- | 2596 | 2023-04-11 08:46:32 | | | |
| 2 | Camila Xu | -3 word(s) | 2593 | 2023-04-11 10:14:15 | | | | |
| 3 | Camila Xu | Meta information modification | 2593 | 2023-04-11 10:14:43 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Pruteanu, L.L.; Bailey, D.S.; Grădinaru, A.C.; Jäntschi, L. Biochemistry of Antioxidants and Their Mode of Action. Encyclopedia. Available online: https://encyclopedia.pub/entry/42918 (accessed on 25 July 2026).
Pruteanu LL, Bailey DS, Grădinaru AC, Jäntschi L. Biochemistry of Antioxidants and Their Mode of Action. Encyclopedia. Available at: https://encyclopedia.pub/entry/42918. Accessed July 25, 2026.
Pruteanu, Lavinia Lorena, David Stanley Bailey, Andrei Cristian Grădinaru, Lorentz Jäntschi. "Biochemistry of Antioxidants and Their Mode of Action" Encyclopedia, https://encyclopedia.pub/entry/42918 (accessed July 25, 2026).
Pruteanu, L.L., Bailey, D.S., Grădinaru, A.C., & Jäntschi, L. (2023, April 11). Biochemistry of Antioxidants and Their Mode of Action. In Encyclopedia. https://encyclopedia.pub/entry/42918
Pruteanu, Lavinia Lorena, et al. "Biochemistry of Antioxidants and Their Mode of Action." Encyclopedia. Web. 11 April, 2023.
Copy Citation
Antioxidants are a class of a multitude of chemical substances clearly associated with large health benefits and lower risks of various age-related diseases.
biochemistry
health
antioxidants
free radicals
1. Introduction
Antioxidants are a class of a multitude of chemical substances clearly associated with large health benefits and lower risks of various age-related diseases.
They also can stop the damaging actions of reactive oxygen species (ROS) [1][2], which include partially reduced or “energized” forms of oxygen, some of them as “free radicals”, with an unpaired electron included in an orbital, while others as “nonradical species”, such as hydrogen peroxide and singlet oxygen, whose reactivity is even greater than that of the ground state of molecular oxygen [1][3].
2. Biochemistry of Antioxidants and Their Mode of Action
Endogenous antioxidants are body products. In contrast with the exogenous antioxidants, the body possesses enzyme systems with an antioxidant action (superoxide dismutase, glutathione peroxidase, and catalysis), co-participating in the deactivation of some free radicals that are formed in the body [4][5]. As a defense against oxidative stress imbalances, the body has produced so-called endogenous antioxidants, enzyme systems capable of annihilating free oxygen molecules, preventing the production of negative effects in the body [6][7]. Among the endogenous antioxidants, superoxide-dismutase, catalase, glutathione peroxidase, and hydropersulfides are mentioned [8][9]. Some subtypes of glutathione peroxidase are selenium (Se)-dependent, and recent studies [10][11][12][13][14] show that an increased intake of Se is associated with protection against the development of cancer and other chronic diseases.
Antioxidants as food additives are referring to some natural or synthetic (established) antioxidants which are also widely used in the food industry to prevent a reduction in the oxidation of fats or other components present in food, during the preservation period [15][16].
Exogenous antioxidants are introduced with food and are referred to the established or natural antioxidants. Because it is much more effective, and cheaper, to maintain good health rather than to regain it, the best protection against free radicals is to build and maintain “antioxidant shields”, through a regimen of adequate food with little fat, rich in digestive fibers, and in antioxidant substances, such as vitamin E, vitamin C, and beta-carotene, combined with regular exercise [17][18], and through a life program [19] aimed at avoiding, as much as possible, the situations in which researchers are exposed to the attack of free radicals. Foods with a high content of antioxidants constitute the basis of nutritional strategies that external sources can be taken from [20][21]. Food of plant origin was associated with a high content of antioxidants [21][22]. Importantly, exogenous antioxidants that can be taken in the diet have the same role in reducing the excessive number of free radicals. The most important of these external (or exogenous) antioxidants are vitamin C, vitamin E, and beta-carotene [23].
Taking the example of vitamin C’s different pathways to biosynthesis in marine algae or plants, animals, and the human bodies, the importance of the exogenous addition of antioxidants such as vitamin C in the human body can be emphasized. The Smirnoff–Wheeler pathway, in which vitamin C is synthesized from D-mannose and L-galactose (D-mannose/L-galactose pathway) [24], represents the major route of vitamin C biosynthesis in marine algae and plants, at the cellular level (Figure 1), the other three involved routes being the glucose, myoinositol, and the galacturonate pathways [25][26][27][28]. Most animals produce relatively high levels of ascorbic acid from glucose in the liver via the glucuronic acid pathway (Figure 1) [29][30][31][32]. Humans are unable to synthesize vitamin C and must ingest this vitamin [33][34].
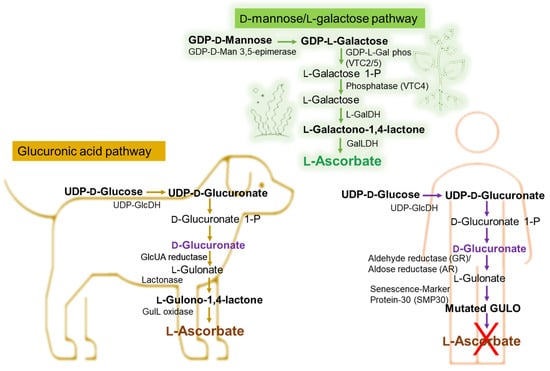
Figure 1. Biochemical pathway of vitamin C synthesis in animals vs. plants/green algae and humans. GDP-L-Gal phos, guanosine diphosphate-L-galactose phosphorylase; GalDH, galactose dehydrogenase; UDP-GlcDH, uridine diphosphate glucose dehydrogenase; GlcUA reductase, glucuronic acid reductase; GulL oxidase (GULO), L-gulonolactone oxidase. This figure is based on information from references [35][36][37][38][39].
As shown in Figure 1, in certain vertebrates (i.e., dogs), L-Ascorbate synthesis involves three enzymatic steps starting from the conversion of D-Glucuronate, with L-Gulonate and L-Gulono-γ-lactone (L-Gulono-1,4-lactone) as intermediate metabolites [35][36][37][38][39][40]. If in this case, the final enzymatic step is catalyzed by L-Gulono-γ-lactone oxidase (GULO), converting L-Gulonate to L-Ascorbate, in humans, the GULO enzyme is mutated (Figure 1) and not functional in primates also including guinea pigs and some spontaneous mutant mouse and rat models [39][41]. Instead, the conversion of L-Gulonate to L-Gulono-1,4-lactone occurs via Senescence-Marker Protein-30 (SMP30) (Figure 1) also known as regucalcin [36][39]. Likewise, in the humans’ case, the conversion of D-Glucuronate to L-Gulonate occurs mainly through aldehyde reductase (GR) and to a smaller extent with aldose reductase’ (AR) contribution (Figure 1) [25][39][40]. Considering that the common molecular mechanism of the body’s limited ability to synthesize vitamin C is the absence of GULO [42][43][44][45], genetically, it is considered that the loss of synthesizing the ascorbic acid is likely due to the complete loss of the L-gulono-γ-lactone oxidase (GULO) gene.
Combinations of all these facts bring us to the conclusion of an important balanced diet associated with antioxidant supplements, potentiating each other’s effects and influencing the prevention of diseases, such as heart disease, arthritis, visual impairment, stroke, and premature aging of the skin, enhancing well-being.
Membrane lipids represent a major target of the radical attack, due to the presence of double bonds in the structures of the polyunsaturated fatty acids which comprise them. Membrane phospholipids most frequently contain unsaturated fatty acids, i.e., linoleic acid, linolenic acid, and arachidonic acid [46][47]. Membrane lipid peroxidation affects the structure and functions of the plasma membrane and the membranes of intracytoplasmic organelles so that transmembrane potentials, ion fluxes, and transmembrane transport are disturbed, and membrane receptors are inactivated and signaling pathways are deregulated [45][48].
The process of lipid peroxidation changes not only the lipid components of membranes but also the proteins, following the reaction of some amino acids with the aldehyde products of peroxidation [45]. Oxidative changes in proteins under the action of reactive oxygen species can also cause the inactivation of enzymes and membrane proteins [49], thereby producing structural changes that lead to the destabilization of cell morphology. The products generated because of lipid peroxidation are involved in inflammatory diseases [50], aging [51], hepatotoxicity [52], hemolysis [53], and all phases of carcinogenesis during the appearance of malignant tumors and metastases [54].
The effect of reactive oxygen species on enzymes includes, for the most part, decreased catalytic capacity, often caused by the oxidation of sulfhydryl groups and the modification of amino groups [55][56]. Some free radicals result from normal cellular processes, for example, when cells use oxygen as fuel for energy production, free radicals appear as secondary products of this metabolic process necessary to sustain life [57]. On the other hand, both the environment, in which we live, and the living environment are other main factors causing reactive oxygen species [57][58]. Antioxidants can interrupt the sequence of oxidation reactions before it is initiated. In general, antioxidants have a high reduction potential, releasing hydrogen ions, with the inhibition process proceeding as shown in the following representation (Equation (1)) [59]:
InH + RO2− → RO2H + In− (1)
where InH is an antioxidant, RO−2 is a free hydroperoxide radical ion, RO2H is hydroperoxide of, e.g., a fatty acid, and In− is an inactive or weakly active radical ion.
But in all cases, with the increase in the inactivation duration, there is a decrease in the number of antioxidants—the increase in the peroxide index is found only after there has been a significant decrease in the added antioxidant [2][60][61]. Taking here the α-tocopherol (Vitamin E) as an example of lipid-soluble antioxidant, which acts as a “chain breakerf” to intercept lipid peroxyl radicals (LOO˙) and to terminate the lipid peroxidation chain reactions (Equation (2)) [55], it can be seen that the mechanism of action is much more complex, as the antioxidants can act at successive steps of initiation, propagation, and chain termination of the oxidative radical process [56].
OO˙ + α-tocopherol–OH → LOOH + α-tocopherol–O˙ (2)
It can be explained that there is a close correlation between the structure of antioxidants and their mode of action, determined by factors as follows [62][63]:
-
The presence of the aromatic nucleus of phenol or naphthol, of a secondary or tertiary hydroxyl group which increases the effectiveness (most antioxidants have a phenolic structure).
-
The presence of allylic groups in the ortho or para position compared to the hydroxyl groups which have a favorable effect.
-
The antioxidant effect increases proportionally with the length of the chain.
-
Alkylation in the meta position is less effective.
-
The esterification of the hydroxyl groups which causes a total disappearance of the antioxidant activity.
A classification of antioxidants according to their mode of action is presented in Table 1 [61][64].
Table 1. Inhibitors of lipid oxidation reactions.
| Type | Mode of Action | Examples |
|---|---|---|
| “Metal scavenger” | Chelates metal ions such as copper and iron, forming inactive complexes |
Chelating agents such as EDTA, citric acid, phospholipids, polyphosphates |
| “Oxygen scavenger” | Reacts with oxygen; reduces oxygen | Ascorbic acid, ascorbyl-palmitate |
| Antioxidant (AH) | Interrupts propagation stages in the case of oxidation reactions; donates a hydrogen atom | Phenolic compounds such as BHA, BHT, TBHQ, PG, tocopherols, hydroxytyrosol, caffeic acid, carnosol, etc. |
| Reducing agents (RSH) | Regenerates phenols (synergism) | Ascorbic acid |
| Enzymatic antioxidant | Removes dissolved oxygen or oxidative species | Superoxide dismutase, glutathione peroxidase, glucose-oxidase-catalase |
| Antioxidants with multiple functions | Regenerates primary antioxidants chelated with metals; reduces hydroperoxides | Phospholipids (phosphatidyl-ethanol amine- fish oil), products of the Maillard reaction |
| Methyl-silicone and ethylidene phytosterols | They prevent oxidative polymerization in heated oils |
Polydimethylsiloxane, citrostadienol |
Glutathione peroxidase (GSHPx), catalase (CAT), and superoxide dismutase (SOD) act as the first-line defense antioxidants, as their importance is especially related to superoxide anion radical (*O2) which is perpetually generated in normal body metabolism, particularly through the mitochondrial energy production pathway (MEPP) and their fundamental role in preventing oxidative stress and the cellular damage [5][55][60].
Glutathione is a nonenzymatic antioxidant that is found in most cells, and tissues of plants and animals, and in humans, the highest levels are in the liver, lens, pancreas, spleen, and kidney [65][66]. It is mainly synthesized by the body [66], and it can increase the level of cytotoxic T cells in lymphocytes and neutralize free radicals [67][68]. Given that glutathione has a tripeptide composition of cysteine, glutamate, and glycine, it has an active thiol (SH−) within the cysteine structure [6][8]. In the cell, >98% of glutathione is found in the reduced thiol form (GSH) [5][66], but due to the cysteine residues that can be easily oxidized nonenzymatically by various electrophilic substances (free radicals, reactive oxygen, and nitrogen species), it is also present in the oxidized form as glutathione disulfide (GSSG) or glutathione peroxidase [5][66]. After synthesis, it is distributed to intracellular compartments and the extracellular space for use by other cells and tissues [66][68]. The rate of GSH synthesis is largely controlled by the degree of expression and catalytic activity of the enzyme γ-glutamyl-cysteine synthetase (GCS) and the cellular availability of cysteine [5][60][66][67][68]. Oxidative stress, inflammatory cytokines, cancer, chemotherapy, ionizing radiation, heat shock, inhibition of GCS activity, GSH depletion, GSH conjugation, heavy metals, antioxidants, and insulin increase γ-glutamyl-cysteine synthase transcription or activity in a wide variety of cells [69][70]. In contrast, protein deficiency, dexamethasone, erythropoietin, TNF-β (tumor necrosis factor), hyperglycemia, and GCS phosphorylation decrease GCS transcription or activity [69][71][72]. The glutathione system also represents a “capture system” for peroxides from water metabolism and lipid peroxides permanently formed in the cell, metabolizing them with the formation of water and oxygen [5][66][67][68]. It provides important protection for the mitochondrial and cell membrane against the harmful effects of reactive oxygen species (oxidative stress) [5], protects the tertiary structure of proteins, and activates the transport of amino acids through the cell membrane [5][73]. The cellular level of glutathione is stimulated by alpha lipoic acid, glutamine, colostrum, selenium, and vitamins C, B6, and B2, and the effectiveness of vitamins C, E, and coenzyme Q10 depends on the level of glutathione in the body [74]. Food sources rich in GSH are generally green leafy vegetables, such as spinach, parsley, and broccoli. However, glutathione from food is only partially absorbed, being mostly hydrolyzed by peptidases [74][75][76]. However, the diet plays an important role in the exogenous intake of glutathione by providing important cofactors, such as Se, Mn, Zn, and S-containing amino acids. GSH has a dual role in our health and pathology as an antioxidant and in the detoxification of certain xenobiotics [74][75][76].
Catalases. While GSHPxs are cytosolic residents, catalases are mainly found in peroxisomes, in the liver, and erythrocytes, but some catalases are found in all tissues [60][77], being the first characterized antioxidant enzymes [66] and being one of the crucial antioxidant enzymes that mitigate oxidative stress to a considerable extent by destroying cellular hydrogen peroxide to produce water and oxygen by using either iron or manganese as a cofactor [5][78]. Basically, they are present in almost all living tissues that utilize oxygen [5]. Due to its chemical structure of four subunits, each containing a heme group and a molecule of NADPH, catalase basically catalyzes the conversion of hydrogen peroxide to water and oxygen [60], while superoxide dismutase (one of the most potent intracellular enzymatic antioxidants) catalyzes the conversion of superoxide anions to dioxygen and hydrogen peroxide [60]. Hence, all three, catalase, glutathione peroxidase, and superoxide dismutase, are functionally interconnected due to the hydrogen peroxide (H2O2), which is produced as a result of the reaction catalyzed by SOD, H2O2 being the substrate of both CAT and GSHPx [65]. Deficiency or malfunction of catalase causes aging disorders and pathogenesis of degenerative diseases, such as diabetes mellitus, hypertension, anemia, vitiligo, Alzheimer’s disease, Parkinson’s disease, bipolar disorder, cancer, schizophrenia, or even male infertility [78][79].
Superoxide dismutase. Depending on its expressed activity, superoxide dismutase may act either as an antioxidant or as a prooxidant [66] as exists in several isoforms, differing in the active metal center, amino acid composition, cofactors, and other properties [60], and neutralizes superoxide ions by going through successive oxidative and reductive cycles of transition metal ions at its active site [60]. In humans, three forms of SOD are present: cytosolic Cu, Zn-SOD (consisting of a dinuclear metal cluster with copper and zinc ions, which catalyzes the dismutation of the superoxide anion to oxygen and water), the mitochondrial Mn-SOD (a homotetramer that includes one manganese atom per subunit, which partitions the superoxide anion), and the extracellular superoxide dismutase containing copper and zinc (a tetrameric secretary glycoprotein having a high affinity for certain glycosaminoglycans) [60][80][81]. In contrast with the fact that superoxide dismutase is indispensable to cellular health, that is protecting body cells from oxidative stress, and that helps in the process of aging or cell death, superoxide dismutase enzyme deficiency is quite common, and more than this, levels of superoxide dismutase decline with age, whereas free radical formation increases [5]. As a result, plant sources of SOD and SOD supplementation became of interest for health enhancement [81][82]. It has been reported that a considerable and adequate daily SOD supplementation protects the immune system and significantly reduces the chances of degenerative diseases and aging pathogenesis, and there are several natural resources that can assure the daily intake of SOD, such as cabbage, Brussels sprouts, wheat grass, barley grass, or broccoli [5].
If the first-line antioxidants act to suppress or prevent the formation of free radicals or reactive species in cells being very fast in neutralizing molecules with the potential of developing into a free radical or neutralizing any free radical with the ability to induce the production of other radicals [5], the second-line defense antioxidants (scavenging antioxidants) are neutralizing or scavenging free radicals by donating an electron to them, becoming free radicals themselves but of lesser damaging effects [5][60]. These are mainly represented by hydrophilic antioxidants, such as ascorbic acid, uric acid, and glutathione, and by lipophilic antioxidants, such as alpha-tocopherol (vitamin E) and ubiquinol [5][60].
After free radical damage has occurred, a third category of antioxidants (de novo enzymes), such as polymerases, glycosylases, nucleases, proteinases, proteases, and peptidases, are acting towards repairing the damage caused to biomolecules and reconstitute the damaged cell membrane [5], while a fourth-line defense antioxidants can prevent the formation or reaction of free radicals [5][60].
References
- Benzie, I.F.F.; Choi, S.W. Antioxidants in food: Content measurement, significance, action, cautions, caveats, and research needs (Chapter 1). Adv. Food Nutr. 2014, 71, 1–53.
- Lü, J.M.; Lin, P.H.; Yao, Q.; Chen, C. Chemical and Molecular Mechanisms of Antioxidants: Experimental Approaches and Model Systems. J. Cell. Mol. Med. 2010, 14, 840–860.
- Halliwell, B. Reactive oxygen species (ROS), oxygen radicals and antioxidants: Where are we now, where is the field going and where should we go? Biochem. Biophys. Res. Commun. 2022, 633, 17–19.
- Yu, S.; Huang, X.; Miao, L.; Zhu, J.; Yin, Y.; Luo, Q.; Xu, J.; Shen, J.; Liu, J. A supramolecular bifunctional artificial enzyme with superoxid dismutase and glutathione peroxidase activities. Bioorg. Chem. 2010, 38, 159–164.
- Ighodaro, O.M.; Akinloye, O.A. First time defense antioxidants—Superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPx): Their functional role in the entire antioxidant defense grid. Alex. J. Med. 2018, 54, 287–293.
- Mirończuk-Chodakowska, I.; Witkowska, A.M.; Zujko, M.E. Endogenous non-enzymatic antioxidants in the human body. Adv. Med. Sci. 2018, 63, 68–78.
- Lange, M.; Olzmann, J.A. Hydropersulfides are endogenous antioxidants that inhibit ferroptosis. Cell Chem. Biol. 2022, 29, 1661–1663.
- Wu, Z.; Khodade, V.S.; Chauvin, J.R.; Rodriguez, D.; Toscano, J.P.; Pratt, D.A. Hydropersulfides Inhibit Lipid Peroxidation and Protect Cells from Ferroptosis. J. Am. Chem. Soc. 2022, 144, 15825–15837.
- Barayeu, U.; Schilling, D.; Eid, M.; da Silva, T.N.X.; Schlicker, L.; Mitreska, N.; Zapp, C.; Gräter, F.; Miller, A.K.; Kappl, R.; et al. Hydropersulfides inhibit lipid peroxidation and ferroptosis by scavenging radicals. Nat. Chem. Biol. 2023, 19, 28–37.
- Razaghi, A.; Poorebrahim, M.; Sarhan, D.; Björnstedt, M. Selenium stimulates the antitumour immunity: Insights to future research. Eur. J. Cancer 2021, 155, 256–267.
- Hu, W.; Zhao, C.; Hu, H.; Yin, S. Food Sources of Selenium and Its Relationship with Chronic Diseases. Nutrients 2021, 13, 1739.
- Wang, N.; Tan, H.Y.; Li, S.; Xu, Y.; Guo, W.; Feng, Y. Supplementation of Micronutrient Selenium in Metabolic Diseases: Its Role as an Antioxidant. Oxidative Med. Cell. Longev. 2017, 2017, 7478523.
- Cardoso, B.R.; Cominetti, C.; Seale, L.A. Editorial: Selenium, Human Health and Chronic Disease. Front. Nutr. 2022, 8, 827759.
- Kuršvietienė, L.; Mongirdienė, A.; Bernatonienė, J.; Šulinskienė, J.; Stanevičienė, I. Selenium Anticancer Properties and Impact on Cellular Redox Status. Antioxidants 2020, 9, 80.
- Silva, M.M.; Lidon, F.C. An overview on applications and side effects of antioxidant food additives. Emir. J. Food Agric. 2016, 28, 823–832.
- Franco, R.; Navarro, G.; Martínez-Pinilla, E. Antioxidants versus Food Antioxidant Additives and Food Preservatives. Antioxidants 2019, 8, 542.
- Adams, A.K.; Best, T.M. The role of antioxidants in exercise and disease prevention. Physician Sportsmed. 2015, 30, 37–44.
- Baranowski, B.J.; Marko, D.M.; Fenech, R.K.; Yang, A.J.T.; MacPherson, R.E.K. Healthy brain, healthy life: A review of diet and exercise interventions to promote brain health and reduce Alzheimer’s disease risk. Appl. Physiol. Nutr. Metab. 2020, 45, 1055–1065.
- Sharifi-Rad, M.; Anil Kumar, N.V.; Zucca, P.; Varoni, E.M.; Dini, L.; Panzarini, E.; Rajkovic, J.; Tsouh Fokou, P.V.; Azzini, E.; Peluso, I.; et al. Lifestyle, Oxidative Stress, and Antioxidants: Back and Forth in the Pathophysiology of Chronic Diseases. Front. Physiol. 2020, 11, 694.
- Wahlqvist, M.L. Antioxidant relevance to human health. Asia Pac. J. Clin. Nutr. 2013, 22, 171–176.
- Zeb, A. Concept, mechanism, and applications of phenolic antioxidants in foods. J. Food Biochem. 2020, 44, e13394.
- Shaw, K.A.; Zello, G.A.; Rodgers, C.D.; Warkentin, T.D.; Baerwald, A.R.; Chilibeck, P.D. Benefits of a plant-based diet and considerations for the athlete. Eur. J. Appl. Physiol. 2022, 122, 1163–1178.
- Serafini, M.; Peluso, I. Functional Foods for Health: The Interrelated Antioxidant and Anti-Inflammatory Role of Fruits, Vegetables, Herbs, Spices and Cocoa in Humans. Curr. Pharm. Des. 2016, 22, 6701–6715.
- Paciolla, C.; Fortunato, S.; Dipierro, N.; Paradiso, A.; De Leonardis, S.; Mastropasqua, L.; de Pinto, M.C. Vitamin C in Plants: From Functions to Biofortification. Antioxidants 2019, 8, 519.
- Smirnoff, N. Ascorbic acid metabolism and functions: A comparison of plants and mammals. Free Radic. Biol. Med. 2018, 122, 116–129.
- Duan, W.; Ren, J.; Li, Y.; Liu, T.; Song, X.; Chen, Z.; Huang, Z.; Hou, X.; Li, Y. Conservation and Expression Patterns Divergence of Ascorbic Acid d-mannose/l-galactose Pathway Genes in Brassica rapa. Front. Plant Sci. 2016, 7, 778.
- Zheng, X.; Gong, M.; Zhang, Q.; Tan, H.; Li, L.; Tang, Y.; Li, Z.; Peng, M.; Deng, W. Metabolism and Regulation of Ascorbic Acid in Fruits. Plants 2022, 11, 1602.
- Broad, R.C.; Bonneau, J.P.; Hellens, R.P.; Johnson, A.A.T. Manipulation of Ascorbate Biosynthetic, Recycling, and Regulatory Pathways for Improved Abiotic Stress Tolerance in Plants. Int. J. Mol. Sci. 2020, 21, 1790.
- Fujii, J. Ascorbate is a multifunctional micronutrient whose synthesis is lacking in primates. J. Clin. Biochem. Nutr. 2021, 69, 1–15.
- Hishiyama, N.; Kayanuma, H.; Matsui, T.; Yano, H.; Suganuma, T.; Funaba, M.; Fujise, H. Plasma concentration of vitamin C in dogs with a portosystemic shunt. Can. J. Vet. Res. 2006, 70, 305–307.
- Gordon, D.S.; Rudinsky, A.J.; Guillaumin, J.; Parker, V.J.; Creighton, K.J. Vitamin C in Health and Disease: A Companion Animal Focus. Top. Companion Anim. Med. 2020, 39, 100432.
- Drouin, G.; Godin, J.R.; Pagé, B. The genetics of vitamin C loss in vertebrates. Curr. Genom. 2011, 12, 371–378.
- Sharma, Y.; Popescu, A.; Horwood, C.; Hakendorf, P.; Thompson, C. Relationship between vitamin C deficiency and cognitive impairment in older hospitalised patients: A cross—Sectional study. Antioxidants 2022, 11, 463.
- DePhillipo, N.N.; Aman, Z.S.; Kennedy, M.I.; Begley, J.P.; Moatshe, G.; LaPrade, R.F. Efficacy of vitamin c supplementation on collagen synthesis and oxidative stress after musculoskeletal injuries: A systematic review. Orthop. J. Sports Med. 2018, 6, 2325967118804544.
- Linster, C.L.; Van Schaftingen, E. Vitamin C biosynthesis, recycling and degradation in mammals. FEBS J. 2007, 274, 1–22.
- Baek, S.-M.; Lee, S.-W.; Kim, T.-U.; Choi, S.-K.; Yun, S.; Lee, W.-J.; Han, S.-H.; Hong, I.-H.; Park, S.-J.; Kim, T.-H.; et al. Senescence Marker Protein 30 (SMP30): A Novel Pan-Species Diagnostic Marker for the Histopathological Diagnosis of Breast Cancer in Humans and Animals. Int. J. Mol. Sci. 2021, 22, 2340.
- Tao, J.; Hao, Z.; Huang, C. Molecular evolution of GDP-L-galactose phosphorylase, a key regulatory gene in plant ascorbate biosynthesis. AoB Plants 2020, 12, plaa055.
- Gabbay, K.H.; Bohren, K.M.; Morello, R.; Bertin, T.; Liu, J.; Vogel, P. Ascorbate synthesis pathway: Dual role of ascorbate in bone homeostasis. J. Biol. Chem. 2010, 285, 19510–19520.
- Yang, H. Conserved or lost: Molecular evolution of the key gene GULO in vertebrate vitamin C biosynthesis. Biochem. Genet. 2013, 51, 413–425.
- Meščić Macan, A.; Gazivoda Kraljević, T.; Raić-Malić, S. Therapeutic Perspective of Vitamin C and Its Derivatives. Antioxidants 2019, 8, 247.
- Nandi, A.; Mukhopadhyay, C.K.; Ghosh, M.K.; Chattopadhyay, D.J.; Chatterjee, I.B. Evolutionary significance of vitamin C biosynthesis in terrestrial vertebrates. Free Radic. Biol. Med. 1997, 22, 1047–1054.
- Schröder, G.C.; Meilleur, F. Metalloprotein catalysis: Structural and mechanistic insights into oxidoreductases from neutron protein crystallography. Acta Crystallogr. Sect. D Struct. Biol. 2021, 77, 1251–1269.
- Fain, O. Musculoskeletal manifestations of scurvy. Jt. Bone Spine 2005, 72, 124–128.
- El Basuini, M.F.; Shahin, S.A.; Teiba, I.I.; Zaki, M.A.A.; El Hais, A.M.; Sewilam, H.; Almeer, R.; Abdelkhalek, N.; Dawood, M.A.O. The influence of dietary coenzyme Q10 and vitamin C on the growth rate, immunity, oxidative-related genes, and the resistance against Streptococcus agalactiae of Nile tilapia (Oreochromis niloticus). Aquaculture 2021, 531, 735862.
- Li, Y.; Schellhorn, H.E. New developments and novel therapeutic perspectives for vitamin C. J. Nutr. 2007, 137, 2171–2184.
- Juan, C.A.; Pérez de la Lastra, J.M.; Plou, F.J.; Pérez-Lebeña, E. The Chemistry of Reactive Oxygen Species (ROS) Revisited: Outlining Their Role in Biological Macromolecules (DNA, Lipids and Proteins) and Induced Pathologies. Int. J. Mol. Sci. 2021, 22, 4642.
- Adwas, A.A.; Elsayed, A.S.I.; Azab, A.E.; Quwaydir, F.A. Oxidative stress and antioxidant mechanisms in human body. J. Appl. Biotechnol. Bioeng. 2019, 6, 43–47.
- Chen, X.; Li, J.; Kang, R.; Klionsky, D.J.; Tang, D. Ferroptosis: Machinery and regulation. Autophagy 2021, 17, 2054–2081.
- Bhattacharjee, S. Membrane lipid peroxidation and its conflict of interest: The two faces of oxidative stress. Curr. Sci. 2014, 107, 1811–1823.
- Hawkins, C.L.; Davies, M.J. Detection, identification, and quantification of oxidative protein modifications. J. Biol. Chem. 2019, 20, 19683–19708.
- Xu, C.; Liu, Z.; Xiao, J. Ferroptosis: A Double-Edged Sword in Gastrointestinal Disease. Int. J. Mol. Sci. 2021, 22, 12403.
- Nguyen-Powanda, P.; Robaire, B. Aging and oxidative stress alter DNA repair mechanisms in male germ cells of superoxide dismutase-1 null mice. Biol. Reprod. 2021, 105, 944–957.
- Bárcena, B.; Salamanca, A.; Pintado, C.; Mazuecos, L.; Villar, M.; Moltó, E.; Bonzón-Kulichenko, E.; Vázquez, J.; Andrés, A.; Gallardo, N. Aging Induces Hepatic Oxidative Stress and Nuclear Proteomic Remodeling in Liver from Wistar Rats. Antioxidants 2021, 10, 1535.
- Menon, A.V.; Liu, J.; Tsai, H.P.; Zeng, L.; Yang, S.; Asnani, A.; Kim, J. Excess heme upregulates heme oxygenase 1 and promotes cardiac ferroptosis in mice with sickle cell disease. Blood 2022, 139, 936–941.
- Jaganjac, M.; Cindric, M.; Jakovcevic, A.; Žarkovic, K.; Žarkovic, N. Lipid peroxidation in brain tumors. Neurochem. Int. 2021, 149, 105–118.
- Satish, B.N.; Dilipkumar, P. Free radicals, natural antioxidants, and their reaction mechanisms. RSC Adv. 2015, 5, 27986–28006.
- Jakubczyk, K.; Dec, K.; Kałduńska, J.; Kawczuga, D.; Kochman, J.; Janda, K. Reactive oxygen species—Sources, functions, oxidative damage. Pol. Merkur. Lekarski. 2020, 48, 124–127.
- Phaniendra, A.; Jestadi, D.B.; Periyasamy, L. Free radicals: Properties, sources, targets, and their implication in various diseases. Indian J. Clin. Biochem. 2015, 30, 11–26.
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74.
- Prenzler, P.D.; Ryan, D.; Robards, K. Introduction to Basic Principles of Antioxidant Activity. In Handbook of Antioxidant Methodology: Approaches to Activity Determination; Royal Society of Chemistry: London, UK, 2021; pp. 1–62.
- Kurutas, E.B. The importance of antioxidants which play the role in cellular response against oxidative/nitrosative stress: Current state. Nutr. J. 2016, 15, 71.
- Musakhanian, J.; Rodier, J.D.; Dave, M. Oxidative Stability in Lipid Formulations: A Review of the Mechanisms, Drivers, and Inhibitors of Oxidation. AAPS PharmSciTech 2022, 23, 151.
- Parcheta, M.; Świsłocka, R.; Orzechowska, S.; Akimowicz, M.; Choińska, R.; Lewandowski, W. Recent Developments in Effective Antioxidants: The Structure and Antioxidant Properties. Materials 2021, 14, 1984.
- Losada-Barreiro, S.; Sezgin-Bayindir, Z.; Paiva-Martins, F.; Bravo-Díaz, C. Biochemistry of Antioxidants: Mechanisms and Pharmaceutical Applications. Biomedicines 2022, 10, 3051.
- Flieger, J.; Flieger, W.; Baj, J.; Maciejewski, R. Antioxidants: Classification, Natural Sources, Activity/Capacity Measurements, and Usefulness for the Synthesis of Nanoparticles. Materials 2021, 14, 4135.
- Irato, P.; Santovito, G. Enzymatic and Non-Enzymatic Molecules with Antioxidant Function. Antioxidants 2021, 10, 579.
- Lushchak, V.I. Glutathione homeostasis and functions: Potential targets for medical interventions. J. Amino Acids 2012, 2012, 736837.
- Kesarwani, P.; Murali, A.K.; Al-Khami, A.A.; Mehrotra, S. Redox regulation of T-cell function: From molecular mechanisms to significance in human health and disease. Antioxid. Redox Signal. 2013, 18, 1497–1534.
- Galano, A.; Alvarez-Idaboy, J.R. Glutathione: Mechanism and kinetics of its non-enzymatic defense action against free radicals. RSC Adv. 2011, 1, 1763–1771.
- Du, X.; Yu, J.; Sun, X.; Qu, S.; Zhang, H.; Hu, M.; Yang, S.; Zhou, P. Impact of epigallocatechin-3-gallate on expression of nuclear factor erythroid 2-related factor 2 and γ-glutamyl cysteine synthetase genes in oxidative stress-induced mouse renal tubular epithelial cells. Mol. Med. Rep. 2018, 17, 7952–7958.
- Espinosa-Diez, C.; Fierro-Fernández, M.; Sánchez-Gómez, F.; Rodríguez-Pascual, F.; Alique, M.; Ruiz-Ortega, M.; Beraza, N.; Martínez-Chantar, M.L.; Fernández-Hernando, C.; Lamas, S. Targeting of Gamma-Glutamyl-Cysteine Ligase by miR-433 Reduces Glutathione Biosynthesis and Promotes TGF-β-Dependent Fibrogenesis. Antioxid. Redox Signal. 2015, 23, 1092–1105.
- Lu, S.C. Regulation of glutathione synthesis. Curr. Top. Cell. Regul. 2001, 36, 95–116.
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Asp. Med. 2009, 30, 1–12.
- Ballatori, N.; Krance, S.M.; Notenboom, S.; Shi, S.; Tieu, K.; Hammond, C.L. Glutathione dysregulation and the etiology and progression of human diseases. Biol. Chem. 2009, 390, 191–214.
- Sitohang, I.B.S.; Anwar, A.I.; Jusuf, N.K.; Arimuko, A.; Norawati, L.; Veronica, S. Evaluating Oral Glutathione Plus Ascorbic Acid, Alpha-lipoic Acid, and Zinc Aspartate as a Skin-lightening Agent: An Indonesian Multicenter, Randomized, Controlled Trial. J. Clin. Aesthetic Dermatol. 2021, 14, 53–58.
- Minich, D.M.; Brown, B.I. A Review of Dietary (Phyto)Nutrients for Glutathione Support. Nutrients 2019, 11, 2073.
- Hossain, A.; Khatun, M.A.; Islam, M.; Huque, R. Enhancement of Antioxidant Quality of Green Leafy Vegetables upon Different Cooking Method. Prev. Nutr. Food Sci. 2017, 22, 216–222.
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183.
- Nandi, A.; Yan, L.J.; Jana, C.K.; Das, N. Role of Catalase in Oxidative Stress- and Age-Associated Degenerative Diseases. Oxidative Med. Cell. Longev. 2019, 2019, 9613090.
- Rubio-Riquelme, N.; Huerta-Retamal, N.; Gómez-Torres, M.J.; Martínez-Espinosa, R.M. Catalase as a Molecular Target for Male Infertility Diagnosis and Monitoring: An Overview. Antioxidants 2020, 9, 78.
- Sheng, Y.; Abreu, I.A.; Cabelli, D.E.; Maroney, M.J.; Miller, A.; Teixeira, M.; Valentine, J.S. Superoxide Dismutases and Superoxide Reductases. Chem. Rev. 2014, 114, 3854–3918.
- Mondola, P.; Damiano, S.; Sasso, A.; Santillo, M. The Cu, Zn Superoxide Dismutase: Not Only a Dismutase Enzyme. Front. Physiol. 2016, 7, 594.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.4K
Revisions:
3 times
(View History)
Update Date:
11 Apr 2023
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No