 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Norizah Mhd Sarbon | -- | 3447 | 2023-03-23 20:37:49 | | | |
| 2 | Rita Xu | -4 word(s) | 3443 | 2023-03-24 02:38:19 | | |
Video Upload Options
Consumption of legumes has been shown to enhance health and lower the risk of cardiovascular disease and specific types of cancer. ACE inhibitors, antioxidants, and synthetic anti-inflammatories are widely used today. In vitro and in vivo research has shown the bioactive peptides generated from legume protein hydrolysates, such as antioxidant, anti-hypertensive, anticancer, anti-proliferative, anti-inflammatory, etc., in the context of different disease mitigation. Therefore, researchers describe the recent advances in in vitro and in vivo studies of antioxidant, anti-hypertensive and anti-inflammatory peptides isolated from legume-derived protein hydrolysates. The results indicated that antioxidant legumes peptides are characterized by shortchain sequence amino acids and possess anti-hypertensive properties by reducing systolic blood pressure (SBP) in spontaneously hypertensive rats (SHR).
1. Introduction
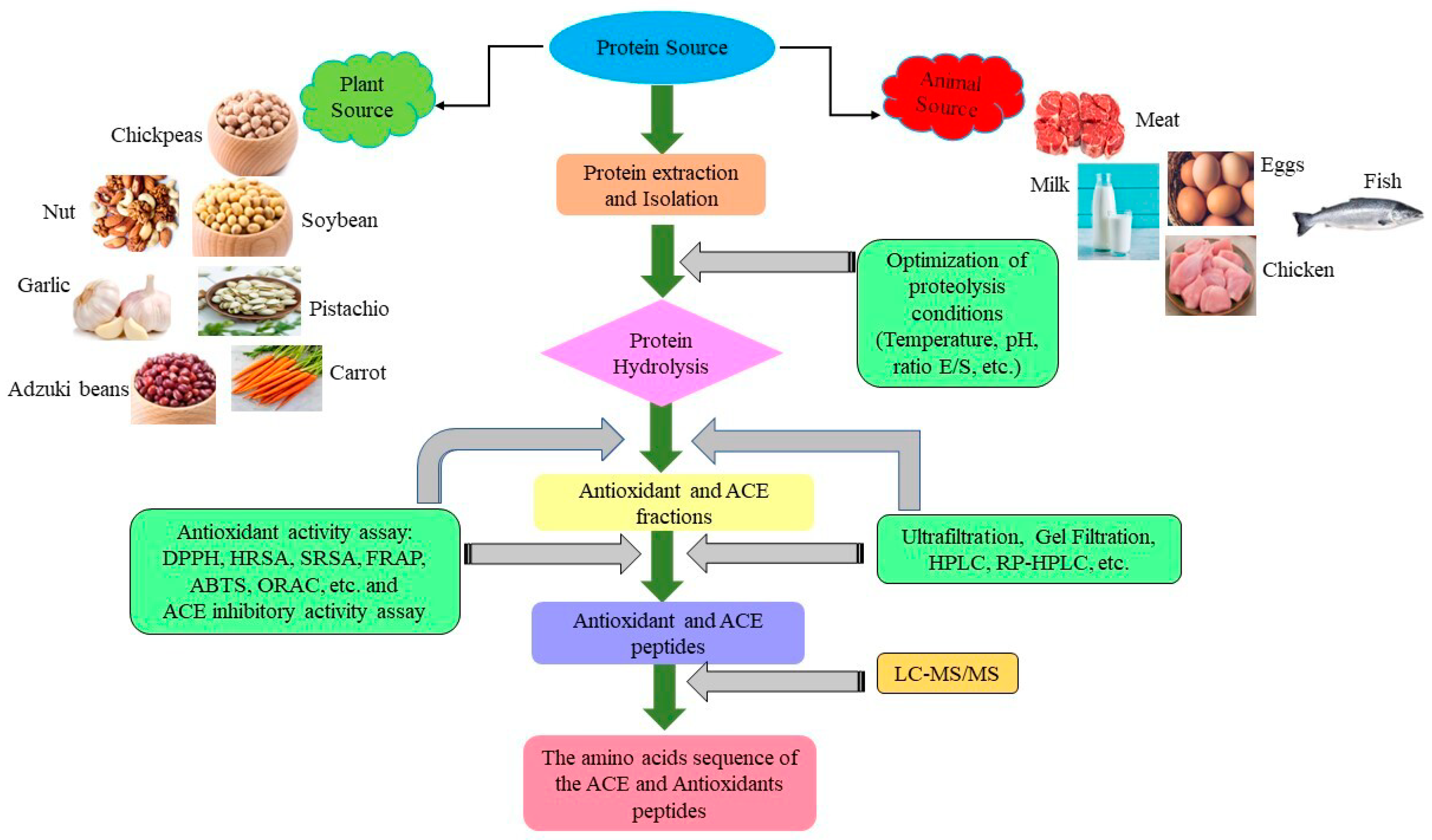
2. Legume Protein Isolate and Hydrolysate
3. In Vitro and In Vivo Studies on the Antioxidant, ACE-Inhibitory, and Anti-Inflammatory Peptides from Legume Protein Hydrolysate
3.1. In-Vitro Study of Antioxidant, ACE-Inhibitory, and Anti-Inflammatory Peptides
3.1.1. Antioxidant Peptide
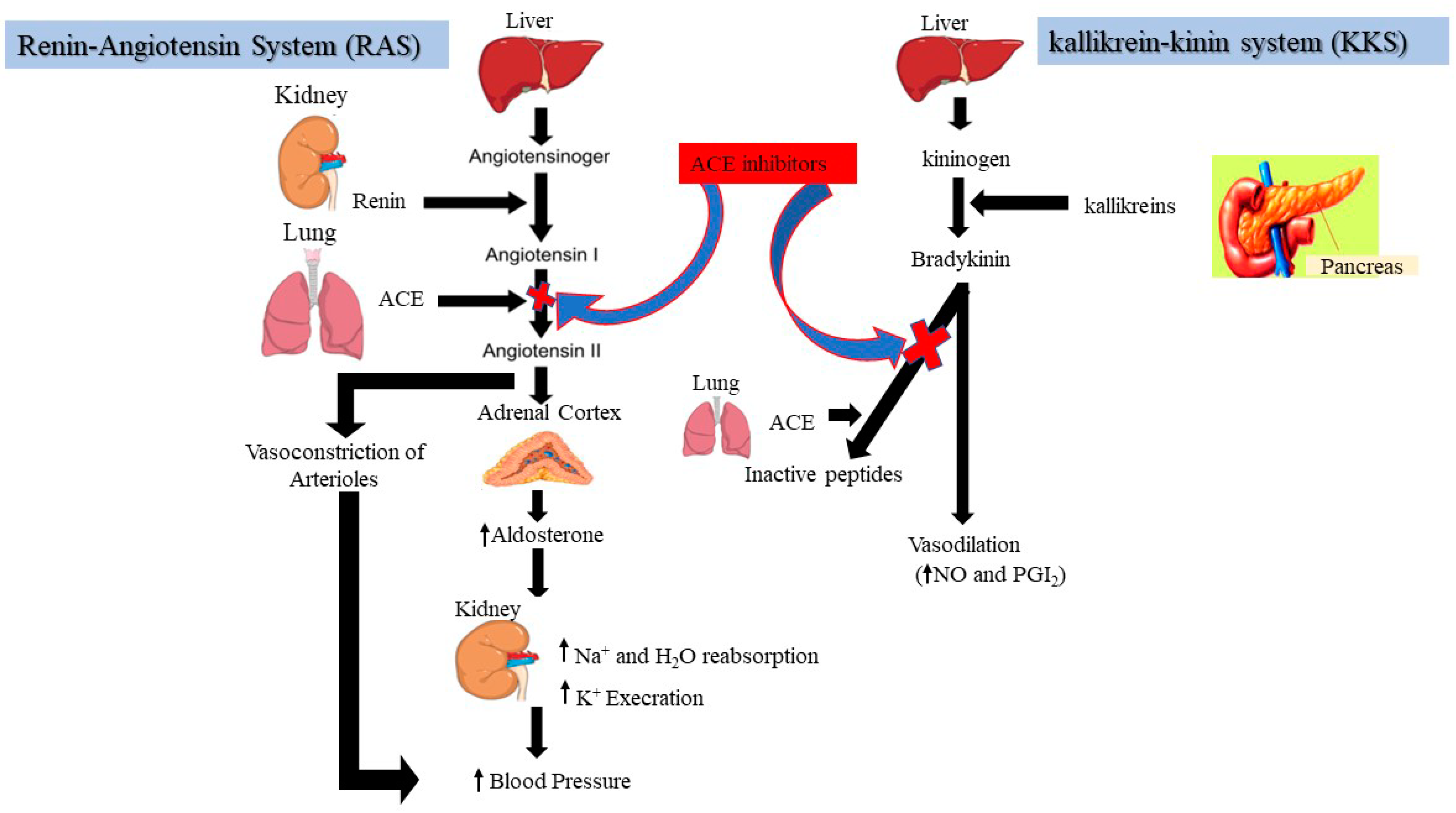
3.1.2. ACE-Inhibitory Peptide
3.1.3. Anti-Inflammatory Peptide
References
- Tripathi, K.; Gore, P.G.; Singh, M.; Pamarthi, R.K.; Mehra, R.; VanGayacharan, C. Legume Genetic Resources: Status and Opportunities for Sustainability. In Legume Crops-Prospects, Production and Uses; Hasanuzzaman, M., Ed.; BoD–Books on Demand: London, UK, 2020.
- Carbonaro, M.; Nucara, A. Legume Proteins and Peptides as Compounds in Nutraceuticals: A Structural Basis for Dietary Health Effects. Nutrients 2022, 14, 1188.
- Olagunju, A.I.; Omoba, O.S.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Antioxidant properties, ACE/renin inhibitory activities of pigeon pea hydrolysates and effects on systolic blood pressure of spontaneously hypertensive rats. Food Sci. Technol. 2018, 6, 1879–1889.
- Hanafi, M.A.; Hashim, S.N.; Chay, S.Y.; Ebrahimpour, A.; Zarei, M.; Muhammad, K.; Saari, N. High angiotensin-I converting enzyme (ACE) inhibitory activity of Alcalase-digested green soybean (Glycine max) hydrolysates. Food Res. Int. 2018, 106, 589–597.
- Sánchez-Chino, X.M.; Jiménez Martínez, C.; León-Espinosa, E.B.; Garduño-Siciliano, L.; Álvarez-González, I.; Madrigal-Bujaidar, E.; Dávila-Ortiz, G. Protective effect of chickpea protein hydrolysates on colon carcinogenesis associated with a hypercaloric diet. J. Am. Coll. Nutr. 2019, 38, 162–170.
- Matemu, A.; Nakamura, S.; Katayama, S. Health benefits of antioxidative peptides derived from legume proteins with a high amino acid score. Antioxidants 2021, 10, 316.
- Wen, C.; Liu, G.; Ren, J.; Deng, Q.; Xu, X.; Zhang, J. Current Progress in the Extraction, Functional Properties, Interaction with Polyphenols, and Application of Legume Protein. J. Agric. Food Chem. 2022, 70, 992–1002.
- Oo, Z.Z.; Ko, T.L.; Than, S.S. Chemical and functional characterizations of chickpea protein concentrate. ASRJETS 2017, 38, 272–280.
- Rababah, T.; Albiss, B.A.; Al-U’datt, M.; Akkam, Y.; Abu Kayed, A. Effect of Ultrasound Treatment on the Physicochemical, Nutraceutical, and Functional Properties of Lupine Flour. J. Agric. Sci. Technol. 2021, 23, 825–838.
- Boukid, F. Chickpea (Cicer arietinum L.) protein as a prospective plant-based ingredient: A review. Int. J. Food Sci. 2021, 56, 5435–5444.
- Chen, Y.; Zheng, Z.; Ai, Z.; Zhang, Y.; Tan, C.P.; Liu, Y. Exploring the Antioxidant and Structural Properties of Black Bean Protein Hydrolysate and Its Peptide Fractions. Front. Nutr. 2022, 9, 884537.
- Tong, X.; Cao, J.; Tian, T.; Lyu, B.; Miao, L.; Lian, Z.; Jiang, L. Changes in structure, rheological property and antioxidant activity of soy protein isolate fibrils by ultrasound pretreatment and EGCG. Food Hydrocoll. 2022, 122, 107084.
- Putra, I.D.; Marsono, Y.; Indrati, R. Effect of simulated gastrointestinal digestion of bioactive peptide from pigeon pea (Cajanus cajan) tempe on angiotensin-I converting enzyme inhibitory activity. Nutr. Food Sci. 2020, 51, 244–254.
- Langton, M.; Ehsanzamir, S.; Karkehabadi, S.; Feng, X.; Johansson, M.; Johansson, D.P. Gelation of faba bean proteins-Effect of extraction method, pH and NaCl. Food Hydrocoll. 2020, 103, 105622.
- Sonklin, C.; Alashi, M.A.; Laohakunjit, N.; Kerdchoechuen, O.; Aluko, R.E. Identification of antihypertensive peptides from mung bean protein hydrolysate and their effects in spontaneously hypertensive rats. J. Funct. Foods 2020, 64, 103635.
- Karimi, A.; Gavlighi, H.A.; Sarteshnizi, R.A.; Udenigwe, C.C. Effect of maize germ protein hydrolysate addition on digestion, in vitro antioxidant activity and quality characteristics of bread. J. Cereal Sci. 2021, 97, 103148.
- Wang, R.; Zhao, H.; Pan, X.; Orfila, C.; Lu, W.; Ma, Y. Preparation of bioactive peptides with antidiabetic, antihypertensive, and antioxidant activities and identification of α-glucosidase inhibitory peptides from soy protein. Food Sci. Nutr. 2019, 7, 1848–1856.
- Olagunju, A.I.; Omoba, O.S.; Enujiugha, V.N.; Alashi, A.M.; Aluko, R.E. Pigeon pea enzymatic protein hydrolysates and ultrafiltration peptide fractions as potential sources of antioxidant peptides: An in vitro study. Food Sci. Technol. 2018, 97, 269–278.
- Ahmed, J.; Mulla, M.; Al-Ruwaih, N.; Arfat, Y.A. Effect of high-pressure treatment prior to enzymatic hydrolysis on rheological, thermal, and antioxidant properties of lentil protein isolate. Legum. Sci. 2019, 1, e10.
- Aguilar, J.G.D.S.; Granato Cason, V.; de Castro, R.J.S. Improving antioxidant activity of black bean protein by hydrolysis with protease combinations. Int. J. Food Sci. 2019, 54, 34–41.
- Chunkao, S.; Youravong, W.; Yupanqui, C.T.; Alashi, A.M.; Aluko, R.E. Structure and Function of Mung Bean Protein-Derived Iron-Binding Antioxidant Peptides. Foods. 2020, 9, 1406.
- Gupta, N.; Bhagyawant, S.S. Angiotensin-I converting enzyme (ACE-I) inhibitory and anti-proliferative potential of chickpea seed protein hydrolysate. Ann. Plant Prot. Sci. 2018, 7, 2149–2153.
- Xu, Z.; Wu, C.; Sun-Waterhouse, D.; Zhao, T.; Waterhouse, G.I.; Zhao, M.; Su, G. Identification of post-digestion angiotensin-I converting enzyme (ACE) inhibitory peptides from soybean protein Isolate: Their production conditions and in silico molecular docking with ACE. Food Chem. 2021, 345, 128855.
- Ratnayani, K.; Suter, I.K.; Antara, N.S.; Putra, I.N.K. Effect of in vitro gastrointestinal digestion on the Angiotensin Converting Enzyme (ACE) inhibitory activity of pigeon pea protein isolate. Int. Food Res. J. 2019, 26, 1397–1404.
- Dang, Y.; Zhou, T.; Hao, L.; Cao, J.; Sun, Y.; Pan, D. In vitro and in vivo studies on the angiotensin-converting enzyme inhibitory activity peptides isolated from broccoli protein hydrolysate. Food Chem. 2019, 67, 6757–6764.
- Ijarotimi, O.S.; Malomo, S.A.; Alashi, A.M.; Nwachukwu, I.D.; Fagbemi, T.N.; Osundahunsi, O.F.; Aluko, R.E. Antioxidant and antihypertensive activities of wonderful cola (Buchholzia coriacea) seed protein and enzymatic protein hydrolysates. J. Food Bioact. 2018, 3, 133–143.
- Wang, X.; Chen, H.; Fu, X.; Li, S.; Wei, J. A novel antioxidant and ACE inhibitory peptide from rice bran protein: Biochemical characterization and molecular docking study. Food Sci. Technol. 2017, 75, 93–99.
- Tawalbeh, D.; Ahmad, W.W.; Sarbon, N.M. Effect of ultrasound pretreatment on the functional and bioactive properties of legumes protein hydrolysates and peptides: A comprehensive review. Food Rev. Int. 2022, 1–23.
- Wang, R.; Lu, X.; Sun, Q.; Gao, J.; Ma, L.; Huang, J. Novel ACE inhibitory peptides derived from simulated gastrointestinal digestion in vitro of sesame (Sesamum indicum L.) protein and molecular docking study. Int. J. Mol. Sci. 2020, 21, 1059.
- Jebitta, S.R.; Durga Devi, P.R.; Deva Dharshini, L.; Theerdham, N.S.H.; Vignesh, K. A Comprehensive Review on Protein Isolates from Legumes. Int. J. Eng. Technol. 2021, 9, 2277–3878.
- Nadzri, F.A.; Tawalbeh, D.; Sarbon, N.M. Physicochemical properties and antioxidant activity of enzymatic hydrolysed chickpea (Cicer arietinum L.) protein as influence by alcalase and papain enzyme. Biocatal. Agric. Biotechnol. 2021, 36, 102131.
- Wani, I.A.; Sogi, D.S.; Shivhare, U.S.; Gill, B.S. Physico-chemical and functional properties of native and hydrolyzed kidney bean (Phaseolus vulgaris L.) protein isolates. Int. Food Res. J. 2015, 76, 11–18.
- Jakubczyk, A.; Baraniak, B. Angiotensin I converting enzyme inhibitory peptides obtained after in vitro hydrolysis of pea (Pisum sativum var. Bajka) globulins. Biomed Res. Int. 2014, 2014, 438459.
- Ngoh, Y.Y.; Gan, C.Y. Enzyme-assisted extraction and identification of antioxidative and α-amylase inhibitory peptides from Pinto beans (Phaseolus vulgaris cv. Pinto). Food Chem. 2016, 190, 331–337.
- Karaś, M.; Jakubczyk, A.; Szymanowska, U.; Materska, M.; Zielińska, E. Antioxidant activity of protein hydrolysates from raw and heat-treated yellow string beans (Phaseolus vulgaris L.). Acta Sci. Pol. Technol. Aliment. 2014, 13, 385–391.
- Schlegel, K.; Lidzba, N.; Ueberham, E.; Eisner, P.; Schweiggert-Weisz, U. Fermentation of Lupin Protein Hydrolysates—Effects on Their Functional Properties, Sensory Profile and the Allergenic Potential of the Major Lupin Allergen Lup an 1. Foods 2021, 10, 281.
- Jarpa-Parra, M. Lentil protein: A review of functional properties and food application. An overview of lentil protein functionality. Int. J. Food Sci. 2018, 53, 892–903.
- Roy, M.; Sarker, A.; Azad, M.A.K.; Shaheb, M.R.; Hoque, M.M. Evaluation of antioxidant and antimicrobial properties of dark red kidney bean (Phaseolus vulgaris) protein hydrolysates. J. Food Meas. Charact. 2020, 14, 303–313.
- Los, F.G.B.; Demiate, I.M.; Dornelles, R.C.P.; Lamsal, B. Enzymatic hydrolysis of Carioca bean (Phaseolus vulgaris L.) protein as an alternative to commercially rejected grains. Food Sci. Technol. 2020, 125, 109191.
- Rasli, H.; Sarbon, N.M. Optimization of enzymatic hydrolysis conditions and characterization of Shortfin scad (Decapterus macrosoma) skin gelatin hydrolysate using response surface methodology. Int. Food Res. J. 2018, 25, 1541–1549.
- El Hajj, S.; Irankunda, R.; Echavarría, J.A.C.; Arnoux, P.; Paris, C.; Stefan, L.; Canabady-Rochelle, L. Metal-chelating activity of soy and pea protein hydrolysates obtained after different enzymatic treatments from protein isolates. Food Chem. 2023, 405, 134788.
- Márquez, O.G.M.; Juárez-Chairez, M.F.; Márquez-Flores, Y.K.; Jiménez-Martínez, C.; Osorio-Revilla, G. In vitro anti-inflammatory and antioxidant activity of chickpea (Cicer arietinum L.) proteins hydrolysate fractions. Biotecnia 2022, 24, 59–68.
- Karimi, A.; Azizi, M.H.; Ahmadi Gavlighi, H. Fractionation of hydrolysate from corn germ protein by ultrafiltration: In vitro antidiabetic and antioxidant activity. Food Sci. Nutr. 2020, 8, 2395–2405.
- Arámburo-Gálvez, J.G.; Arvizu-Flores, A.A.; Cárdenas-Torres, F.I.; Cabrera-Chávez, F.; Ramírez-Torres, G.I.; Flores-Mendoza, L.K.; Ontiveros, N. Prediction of ACE-I Inhibitory Peptides Derived from Chickpea (Cicer arietinum L.): In Silico Assessments Using Simulated Enzymatic Hydrolysis, Molecular Docking and ADMET Evaluation. Foods 2022, 11, 1576.
- Garcés-Rimón, M.; Morales, D.; Miguel-Castro, M. Potential Role of Bioactive Proteins and Peptides Derived from Legumes towards Metabolic Syndrome. Nutrients 2022, 14, 5271.
- Tacias-Pascacio, V.G.; Morellon-Sterling, R.; Siar, E.H.; Tavano, O.; Berenguer-Murcia, A.; Fernandez-Lafuente, R. Use of Alcalase in the production of bioactive peptides: A review. Int. J. Biol. Macromol. 2020, 165, 2143–2196.
- Yu, J.; Mikiashvili, N.; Bonku, R.; Smith, I.N. Allergenicity, antioxidant activity and ACE-inhibitory activity of protease hydrolyzed peanut flour. Food Chem. 2021, 360, 129992.
- Chen, Z.; Li, W.; Santhanam, R.K.; Wang, C.; Gao, X.; Chen, Y.; Chen, H. Bioactive peptide with antioxidant and anticancer activities from black soybean byproduct: Isolation, identification and molecular docking study. Eur. Food Res. Technol. 2019, 245, 677–689.
- Pan, M.; Liu, K.; Yang, J.; Liu, S.; Wang, S.; Wang, S. Advances on food-derived peptidic antioxidants—A review. Antioxidants 2020, 9, 799.
- Ji, N.; Sun, C.; Zhao, Y.; Xiong, L.; Sun, Q. Purification and identification of antioxidant peptides from peanut protein isolate hydrolysates using UHR-Q-TOF mass spectrometer. Food Chem. 2014, 161, 148–154.
- Soladoye, O.P.; Saldo, J.; Peiro, L.; Rovira, A.; Mor-Mur, M. Antioxidant and angiotensin I converting enzyme inhibitory functions from chicken collagen hydrolysates. J. Nutr. Sci. 2015, 5, 1–10.
- Dabbour, M.; He, R.; Mintah, B.; Ma, H. Antioxidant activities of sunflower protein hydrolysates treated with dual-frequency ultrasonic: Optimization study. J. Food Process Eng. 2019, 42, e13084.
- Noman, A.; Qixing, J.; Xu, Y.; Ali, A.H.; Al-Bukhaiti, W.Q.; Abed, S.M.; Xia, W. Influence of degree of hydrolysis on chemical composition, functional properties, and antioxidant activities of chinese sturgeon (Acipenser sinensis) hydrolysates obtained by using alcalase 2.4 L. J. Aquat. Food Prod. Technol. 2019, 28, 583–597.
- Soleimani, M.; Dehabadi, L.; Wilson, L.D.; Tabil, L.G. Antioxidants classification and applications in lubricants. In Lubrication Tribology, Lubricants, and Additives; Johnson, D.W., Ed.; BoD–Books on Demand: London, UK, 2018; pp. 23–42.
- Phongthai, S.; Rawdkuen, S. Fractionation and characterization of antioxidant peptides from rice bran protein hydrolysates stimulated by in vitro gastrointestinal digestion. Cereal Chem. 2020, 97, 316–325.
- Lourenço, S.C.; Moldão-Martins, M.; Alves, V.D. Antioxidants of natural plant origins: From sources to food industry applications. Molecules 2019, 24, 4132.
- Khammuang, S.; Sarnthima, R.; Sanachai, K. Purification and identification of novel antioxidant peptides from silkworm pupae (Bombyx mori) protein hydrolysate and molecular docking study. Biocatal. Agric. Biotechnol. 2022, 42, 102367.
- Zhang, Q.; Tong, X.; Li, Y.; Wang, H.; Wang, Z.; Qi, B.; Jiang, L. Purification, and characterization of antioxidant peptides from alcalase-hydrolyzed soybean (Glycine max L.) hydrolysate and their cytoprotective effects in human intestinal Caco-2 cells. J. Agric. Food Chem. 2019, 67, 5772–5781.
- Tadesse, S.A.; Emire, S.A. Production and processing of antioxidant bioactive peptides: A driving force for the functional food market. Heliyon 2020, 6, e04765.
- Atta, E.M.; Mohamed, N.H.; Silaev, A.A.A. Antioxidants: An overview on the natural and synthetic types. Eur. Chem. Bull. 2017, 6, 365–375.
- Harvian, Z.A.; Ningrum, A.; Anggrahini, S.; Setyaningsih, W. In silico approach in evaluation of Jack Bean (Canavalia ensiformis) Canavalin protein as precursors of bioactive peptides with dual antioxidant and angiotensin I-converting enzyme inhibitor. Mater. Sci. Forum. 2019, 948, 85–94.
- Zehadi, M.J.A.; Masamba, K.; Li, Y.; Chen, M.; Chen, X.; Sharif, H.R.; Zhong, F. Identification and purification of antioxidant peptides from lentils (Lens Culinaris) hydrolysates. J. Plant Sci. 2015, 3, 123–132.
- Intiquilla, A.; Jiménez-Aliaga, K.; Zavaleta, A.I.; Hernández-Ledesma, B. Production of antioxidant hydrolyzates from a Lupinus mutabilis (Tarwi) protein concentrate with alcalase: Optimization by response surface methodology. Nat. Prod. Commun. 2018, 13, 1934578.
- Wali, A.; Mijiti, Y.; Yanhua, G.; Yili, A.; Aisa, H.A.; Kawuli, A. Isolation and Identification of a Novel Antioxidant Peptide from Chickpea (Cicer arietinum L.) Sprout Protein Hydrolysates. Int. J. Pept. Res. Ther. 2021, 27, 219–227.
- Kou, X.; Gao, J.; Xue, Z.; Zhang, Z.; Wang, H.; Wang, X. Purification and identification of antioxidant peptides from chickpea (Cicer arietinum L.) albumin hydrolysates. LWT 2013, 50, 591–598.
- Malomo, S.A.; Niwachukwu, I.D.; Girgih, A.T.; Idowu, A.O.; Aluka, R.E.; Fagbemi, T.N. Antioxidant and renin-angiotensin system inhibitory properties of cashew nut and fluted-pumpkin protein hydrolysates. Pol. J. Food Nutr. Sci. 2020, 70, 275–289.
- Li, T.; Shi, C.; Zhou, C.; Sun, X.; Ang, Y.; Dong, X.; Zhou, G. Purification and characterization of novel antioxidant peptides from duck breast protein hydrolysates. LWT 2020, 1259, 109215.
- Zheng, Y.; Li, Y.; Zhang, Y.; Ruan, X.; Zhang, R. Purification, characterization, synthesis, in vitro ACE inhibition and in vivo antihypertensive activity of bioactive peptides derived from oil palm kernel glutelin-2 hydrolysates. J. Funct. Foods 2017, 28, 48–58.
- Garcia-Mora, P.; Frias, J.; Peñas, E.; Zieliński, H.; Giménez-Bastida, J.A.; Wiczkowski, W.; Martinez-Villaluenga, C. Simultaneous release of peptides and phenolics with antioxidant, ACE-inhibitory and anti-inflammatory activities from pinto bean (Phaseolus vulgaris L. var. pinto) proteins by subtilisins. J. Funct. Foods. 2015, 18, 319–332.
- Daroit, D.J.; Brandelli, A. In vivo bioactivities of food protein-derived peptides–a current review. Curr. Opin. Food. 2021, 39, 120–129.
- Valenzuela-García, P.; Bobadilla, N.A.; Ramírez-González, V.; León-Villanueva, A.; Lares-Asseff, I.A.; Valdez-Ortiz, A.; Medina-Godoy, S. Antihypertensive effect of protein hydrolysate from azufrado beans in spontaneously hypertensive rats. Cereal Chem. 2017, 94, 117–123.
- Zhang, T.; Li, Y.; Miao, M.; Jiang, B. Purification and characterisation of a new antioxidant peptide from chickpea (Cicer arietium L.) protein hydrolysates. Food Chem. 2011, 128, 28–33.
- Ghribi, A.M.; Sila, A.; Przybylski, R.; Nedjar-Arroume, N.; Makhlouf, I.; Blecker, C.; Besbes, S. Purification and identification of novel antioxidant peptides from enzymatic hydrolysate of chickpea (Cicer arietinum L.) protein concentrate. J. Funct. Foods 2015, 12, 516–525.
- Tang, L.; Sun, J.; Zhang, H.C.; Zhang, C.S.; Yu, L.N.; Bi, J.; Yang, Q.L. Evaluation of physicochemical and antioxidant properties of peanut protein hydrolysate. PLoS ONE 2012, 7, e37863.
- Ding, J.; Liang, R.; Yang, Y.; Sun, N.; Lin, S. Optimization of pea protein hydrolysate preparation and purification of antioxidant peptides based on an in silico analytical approach. LWT 2020, 123, 109126.
- Evangelho, J.A.D.; Berrios, J.D.J.; Pinto, V.Z.; Antunes, M.D.; Vanier, N.L.; Zavareze, E.D.R. Antioxidant activity of black bean (Phaseolus vulgaris L.) protein hydrolysates. Food Sci. Technol. 2016, 36, 23–27.
- Ishak, N.H.; Sarbon, N.M. Optimization of the enzymatic hydrolysis conditions of waste from shortfin scad (Decapterus Macrosoma) for the production of angiotensin I-converting enzyme (ACE) inhibitory peptide using response surface methodology. Int. Food Res. J. 2017, 24, 1735.
- Ghanbari, R.; Zarei, M.; Ebrahimpour, A.; Abdul-Hamid, A.; Ismail, A.; Saari, N. Angiotensin-I converting enzyme (ACE) inhibitory and antioxidant activities of sea cucumber (Actinopyga lecanora) hydrolysates. Int. J. Mol. Sci. 2015, 16, 28870–28885.
- Boschin, G.; Scigliuolo, G.M.; Resta, D.; Arnoldi, A. ACE-inhibitory activity of enzymatic protein hydrolysates from lupin and other legumes. Food Chem. 2014, 145, 34–40.
- Jakubczyk, A.; Karaś, M.; Złotek, U.; Szymanowska, U. Identification of potential inhibitory peptides of enzymes involved in the metabolic syndrome obtained by simulated gastrointestinal digestion of fermented bean (Phaseolus vulgaris L.) seeds. Food Res. Int. 2017, 100, 489–496.
- Nawaz, K.A.; David, S.M.; Murugesh, E.; Thandeeswaran, M.; Kiran, K.G.; Mahendran, R.; Angayarkanni, J. Identification and in silico characterization of a novel peptide inhibitor of angiotensin converting enzyme from pigeon pea (Cajanus cajan). Phytomedicine. 2017, 36, 1–7.
- Ciau-Solís, N.A.; Acevedo-Fernández, J.J.; Betancur-Ancona, D. In vitro renin–angiotensin system inhibition and in vivo antihypertensive activity of peptide fractions from lima bean (Phaseolus lunatus L.). J. Sci. Food Agric. 2018, 98, 781–786.
- Shi, A.; Liu, H.; Liu, L.; Hu, H.; Wang, Q.; Adhikari, B. Isolation, purification and molecular mechanism of a peanut protein-derived ACE-inhibitory peptide. PLoS ONE 2014, 9, e111188.
- Puspitojati, E.; Indrati, R.; Cahyanto, M.N.; Marsono, Y. Formation of ACE-Inhibitory Peptides during Fermentation of Jack Bean Tempe Inoculated by Usar Hibiscus Tiliaceus Leaves Starter. In IOP Conference Series: Environmental Earth Science; IOP Publishing: Bangkok, Thailand, 2019; p. 012022.
- Gupta, N.; Bhagyawant, S.S. Impact of hydrolysis on functional properties, antioxidant, ACE-I inhibitory and anti-proliferative activity of Cicer arietinum and Cicer reticulatum hydrolysates. Nutrire 2019, 44, 5.
- Rivera-Jiménez, J.; Berraquero-García, C.; Pérez-Gálvez, R.; García-Moreno, P.J.; Espejo-Carpio, F.J.; Guadix, A.; Guadix, E.M. Peptides and protein hydrolysates exhibiting anti-inflammatory activity: Sources, structural features and modulation mechanisms. Food Funct. 2022, 13, 12510–12540.
- Dilshara, M.G.; Lee, K.T.; Jayasooriya, R.G.P.T.; Kang, C.H.; Park, S.R.; Choi, Y.H.; Kim, G.Y. Downregulation of NO and PGE2 in LPS-stimulated BV2 microglial cells by trans-isoferulic acid via suppression of PI3K/Akt-dependent NF-κB and activation of Nrf2-mediated HO-1. Int. Immunopharmacol. 2014, 18, 203–211.
- Weissman, S.; Sinh, P.; Mehta, T.I.; Thaker, R.K.; Derman, A.; Heiberger, C.; Tabibian, J.H. Atherosclerotic cardiovascular disease in inflammatory bowel disease: The role of chronic inflammation. WJGP 2020, 11, 104.
- del Carmen Millán-Linares, M.; Millán, F.; Pedroche, J.; del Mar Yust, M. GPETAFLR: A new anti-inflammatory peptide from Lupinus angustifolius L. protein hydrolysate. JFF 2015, 18, 358–367.
- de Medeiros, A.F.; de Queiroz, J.L.C.; Maciel, B.L.L.; de Araújo Morais, A.H. Hydrolyzed Proteins and Vegetable Pep-tides: Anti-Inflammatory Mechanisms in Obesity and Potential Therapeutic Targets. Nutrients 2022, 14, 690.
- Indrati, R. Bioactive Peptides from Legumes and Their Bioavailability. In Legumes Research; Jose, C., Jimenez-Lopez, A.C., Eds.; IntechOpen: London, UK, 2021; Volume 2.
- Montserrat-de la Paz, S.; Villanueva, A.; Pedroche, J.; Millan, F.; Martin, M.E.; Millan-Linares, M.C. Antioxidant and an-ti-inflammatory properties of bioavailable protein hydrolysates from lupin-derived agri-waste. Biomolecules 2021, 11, 1458.
- López-Barrios, L.; Antunes-Ricardo, M.; Gutiérrez-Uribe, J.A. Changes in antioxidant and antiinflammatory activity of black bean (Phaseolus vulgaris L.) protein isolates due to germination and enzymatic digestion. Food Chem. 2016, 203, 417–424.
- Cruz-Chamorro, I.; Álvarez-Sánchez, N.; del Carmen Millán-Linares, M.; del Mar Yust, M.; Pedroche, J.; Millán, F.; Car-rillo-Vico, A. Lupine protein hydrolysates decrease the inflammatory response and improve the oxidative status in human peripheral lymphocytes. Food Res. Int. 2019, 126, 108585.
- González-Montoya, M.; Hernández-Ledesma, B.; Silván, J.M.; Mora-Escobedo, R.; Martínez-Villaluenga, C. Peptides de-rived from in vitro gastrointestinal digestion of germinated soybean proteins inhibit human colon cancer cells prolifera-tion and inflammation. Food Chem. 2018, 242, 75–82.



