Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Eric TOUSSIROT | -- | 2515 | 2023-03-14 11:26:52 | | | |
| 2 | Peter Tang | Meta information modification | 2515 | 2023-03-15 02:22:48 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Toussirot, E. Adipokines in Inflammatory Rheumatic Diseases. Encyclopedia. Available online: https://encyclopedia.pub/entry/42170 (accessed on 06 June 2026).
Toussirot E. Adipokines in Inflammatory Rheumatic Diseases. Encyclopedia. Available at: https://encyclopedia.pub/entry/42170. Accessed June 06, 2026.
Toussirot, Eric. "Adipokines in Inflammatory Rheumatic Diseases" Encyclopedia, https://encyclopedia.pub/entry/42170 (accessed June 06, 2026).
Toussirot, E. (2023, March 14). Adipokines in Inflammatory Rheumatic Diseases. In Encyclopedia. https://encyclopedia.pub/entry/42170
Toussirot, Eric. "Adipokines in Inflammatory Rheumatic Diseases." Encyclopedia. Web. 14 March, 2023.
Copy Citation
Inflammatory rheumatic diseases (IRDs) are complex immune-mediated diseases that are characterized by chronic inflammation of the joints. Rheumatoid arthritis (RA) and spondyloarthritis (SpA), including axial SpA (ax SpA) and psoriatic arthritis (PsA), are the most common forms of IRD. Both RA and ax SpA are characterized by a chronic course with progressive structural modifications, namely, cartilage damage and bone erosions in RA and osteoproliferative changes with spinal ossifications in ax SpA. The adipose tissue is involved in the pathophysiology of IRDs via the release of several proteins, namely, adipokines. Several adipokines with pro-inflammatory effects have been identified, such as leptin, adiponectin, visfatin and resistin.
adipokines
rheumatoid arthritis
axial spondyloarthritis
bone erosion
syndesmophyte
1. Adipokines: Main Biological and Immunological Properties
Adipokines are mainly produced by adipocytes but also by other cellular sources, including immune cells, synoviocytes and chondrocytes [1][2][3]. The contribution of adipokines in IRDs has been extensively described, with high levels of adipokines reported in both the blood and the synovial compartment [2][3][4][5][6][7]. The expression of certain adipokines has also been reported in the synovial tissue [4]. The interplay between adipokines and immune cells from both the innate and adaptive systems has been well described, highlighting the role these mediators play in IRDs [2][3][5].
1.1. Leptin
Leptin is a 16 kDa hormone mainly produced by adipose tissue. Leptin is primarily involved in appetite control and the energetic balance. However, leptin has a wide range of physiological functions [8]. It strongly correlates with fat mass and body mass index (BMI) and is considered to be a surrogate marker of adiposity. Leptin production is dependent on energetic factors, including insulin and sex hormones, but also pro-inflammatory mediators such as TNF alpha, IL-6 and IL-1 beta, which may also stimulate its release [9][10]. Leptin synthesis is stimulated by ovarian sex hormones and inhibited by testosterone, and thus, circulating leptin levels are more elevated in women than in men, even after adjustment for BMI [8]. The immunomodulatory effects of leptin are well described, and in general, it is considered to be a pro-inflammatory adipokine because of its connection with the innate and adaptive immune systems [9]. Indeed, leptin stimulates the production of pro-inflammatory cytokines such as TNF alpha and IL-6, and alternatively, TNF alpha and IL-1 beta increase the expression of leptin in adipose tissue [10]. Leptin may stimulate monocytes, macrophages, dendritic cells, neutrophils and NK cells [9][10]. Leptin increases the phagocytic activity of monocytes/macrophages, inducing the production of nitric oxide and several cytokines [10]. Leptin is able to induce T-cell activation and differentiation toward a Th1 subset, leading to the increased production of IFN-gamma and IL-2 and the decreased production of IL-4 [9]. Leptin is also associated with Th17 cell proliferation and stimulation [11]. Collectively, leptin is considered to have marked pro-inflammatory properties [6][9].
1.2. Adiponectin
Adipose tissue is the main site of adiponectin production, and adiponectin is expressed in different molecular isoforms: globular adiponectin, full-length adiponectin, and low- (LMW), middle- and high-molecular-weight (HMW) adiponectin [2]. Adiponectin has a predominant metabolic function by increasing insulin sensitivity. In fact, patients with insulin resistance and type 2 diabetes (T2D) have low levels of circulating adiponectin. Contrary to leptin, adiponectin levels are decreased in overweight subjects [1]. The influence of adiponectin on the immune system is complex and depends on its different isoforms [2][3][5][12]. Indeed, it is considered that adiponectin has predominant anti-inflammatory effects on atherosclerosis, metabolic syndrome and T2D [13][14]. A favorable effect on cardiovascular (CV) risk has been reported for the specific HMW isoform of adiponectin. In parallel, contradictory effects have been reported in inflammatory disorders such as IRDs [2][5]. Adiponectin may induce the production of anti-inflammatory cytokines such as IL-10 and IL-1 receptor antagonists [15]. TNF alpha and IL-6 may inhibit adiponectin gene expression and protein release. The blockade of the adiponectin receptor AdipoR1 inhibited synovial inflammation in collagen-induced arthritis [16]. In parallel, the different isoforms of adiponectin may exert distinct and sometimes opposing biological functions: LMW adiponectin inhibits LPS-mediated IL-6 release and stimulates IL-10 secretion, while HMW adiponectin induces the secretion of IL-6 by monocytes [17]. In cultured synovial fibroblasts from patients with RA, adiponectin may induce the production of pro-inflammatory mediators [2]. Taken together, it is considered that HMW adiponectin has pro-inflammatory effects on the joint, whereas the LMW isoform is instead associated with anti-inflammatory properties. All in all, adiponectin is considered to have anti-inflammatory effects [13]. However, the opposing effects of adiponectin depend on the relative ratio of its different isoforms, the cytokine environment and the target cell or tissue that is analyzed [5]. In addition, metabolic syndrome (MS) includes different metabolic disorders that increase the risk of diabetes and CV diseases. Visceral fat mass and its derived adipokines may contribute to the development of MS. In this sense, leptin and adiponectin or the leptin/adiponectin ratio may be considered surrogate markers of insulin resistance and MS [18][19].
1.3. Resistin
Resistin is a 12.5 kDa cysteine-rich protein that circulates in the blood as a homodimer. It is mainly produced by mononuclear cells and may induce immune cell activation [2][12][20]. Adipose tissue is also a source of resistin production, but to a lesser extent. Resistin is involved in metabolic functions such as insulin resistance [21]. Resistin is found in areas of inflammation and is capable of inducing the production of IL-6, TNF alpha and IL-1 beta by peripheral blood mononuclear cells [22][23].
1.4. Visfatin
Visfatin (also known as pre-B-cell colony-enhancing factor or nicotinamide phosphoribosyltransferase) is mainly produced by visceral adipose tissue, a fat tissue localization that strongly correlates with increased CV risk. Visfatin regulates insulin secretion and has insulin-like effects [12][20]. Visfatin has pro-inflammatory effects in various cells via the release of TNF alpha, IL-1 beta, IL-6 and chemokines [24].
2. Adipokines in IRD and Their Effects on Joint Components
Adipokines have been extensively studied in patients with RA, but information is less abundant for ax SpA or PsA. The adipokines studied were predominantly leptin and adiponectin and, to a lesser extent, visfatin and resistin [2][3][4][5][12][20][25] (Figure 1).
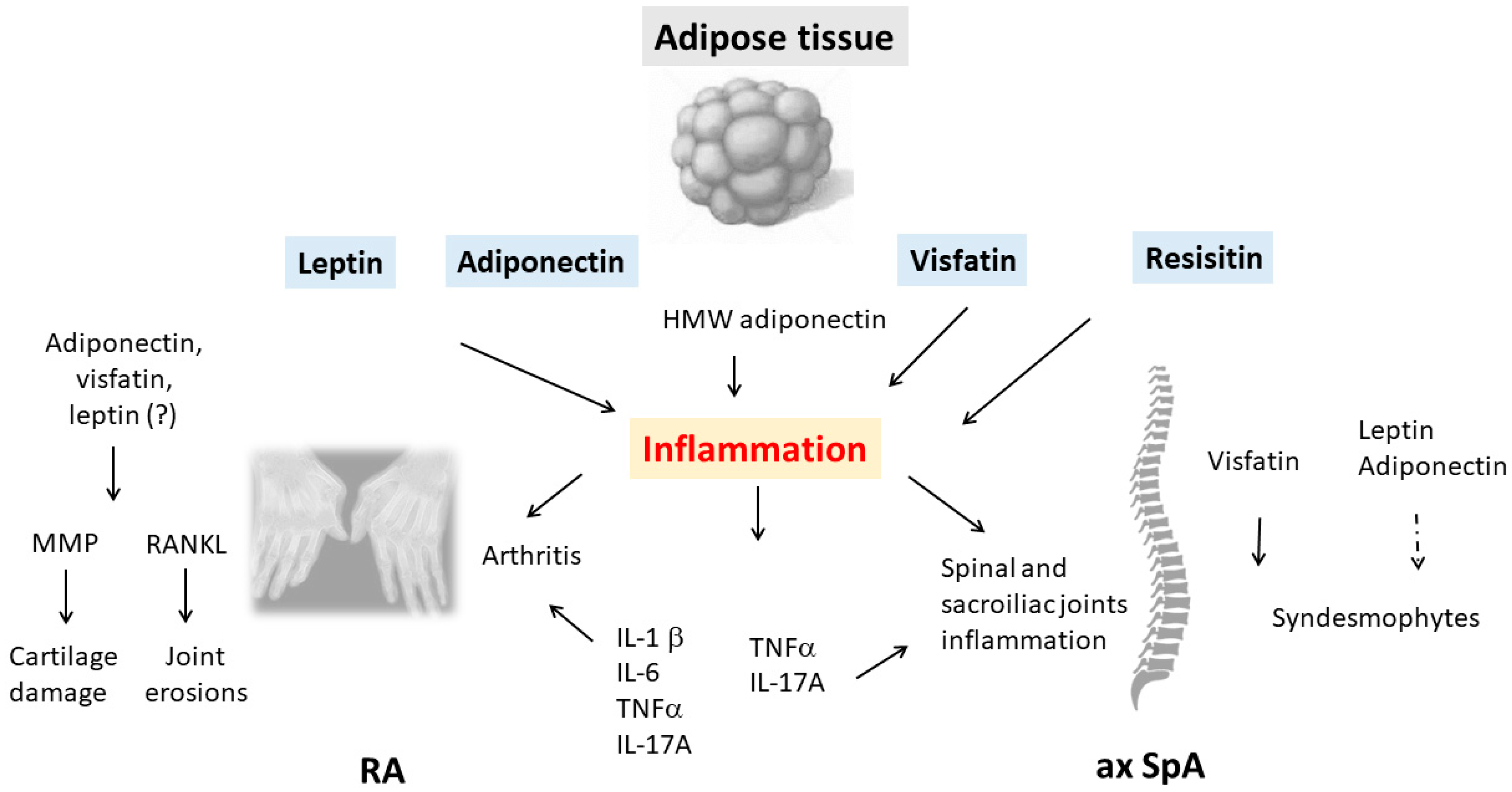
Figure 1. The contribution of adipokines to joint modification in inflammatory rheumatic diseases (IRDs). Several factors are produced by the adipose tissue, collectively called adipokines. Leptin, adiponectin, visfatin and resistin are all involved in inflammation of the joints/axial skeleton in rheumatoid arthritis and axial spondyloarthritis. By stimulating pro-inflammatory cytokine release from innate and adaptive immune cells, adipokines induce an inflammatory milieu. In turn, pro-inflammatory cytokines may promote the release of adipokines. Leptin, visfatin and resistin all have pro-inflammatory effects on the joints and/or axial skeleton, while the effects of adiponectin on the inflammatory process are variable and depend on its isoform, with the high-molecular-weight isoform driving a pro-inflammatory reaction. Due to the interplay that adipokines have with specific factors involved in cartilage and bone metabolism, they may contribute to cartilage degradation and bone erosions in RA or spinal ligamentous ossifications in ax SpA (RA: rheumatoid arthritis; ax SpA: axial spondyloarthritis; HMW: high molecular weight; MMP: metalloproteinases; RANKL: receptor activator of nuclear factor κB ligand; dashed arrow: inhibitory effect; lined arrow: stimulating effect).
2.1. Rheumatoid Arthritis
Collectively, studies have shown higher circulating levels of leptin, visfatin and resistin in patients with RA and the presence of leptin and other adipokines in the synovial fluid of these patients [26][27][28][29][30][31]. A relationship between disease activity and acute-phase reactants such as CRP and leptin has been described, but with contradictory results [32].
Leptin has pro-inflammatory activity and may potentially influence the process of bone erosion in RA by favoring the release of pro-inflammatory mediators, which in turn may activate resident synovial cells to produce metalloproteinases (MPPs) and receptor antagonist of nuclear factor κB ligand (RANKL) for cartilage and bone degradation, respectively [6]. Indeed, in RA synoviocyte-like fibroblasts, leptin may induce the expression of IL-6 and IL-8 with the involvement of the JAK2/STAT3 pathway [33]. In leptin-deficient mice, the severity of arthritis was reduced, along with TNF alpha and IL-1 beta levels [5]. The relationship between periodontal diseases and RA is well established. In this regard, high levels of circulating leptin were reported in patients with early RA and periodontitis and in those with the presence of Porphyromonas gingvalis (P. gingivalis) in the oral cavity [34]. In first-degree relatives of patients with RA, serum leptin levels were associated with tender joints, radiographic changes and also the presence of P. gingivalis [35]. In addition, leptin may induce increased levels of IL-8 and vascular cell adhesion molecule-1 (VCAM-1) in human chondrocytes [36]. Leptin may induce the gene expression of pro-degradative proteases known as ADAMTSs (ADAMTS4, ADAMTS5, ADAMTS9) in human chondrocytes by activating mitogen-activated protein (MAP) kinases and NFκB [37]. Alternatively, leptin may induce the expression of oncostatin M in osteoblasts from healthy human donors [38], as well as RANKL, bone morphogenic protein (BMP) 4 and alkaline phosphatase in mouse vascular smooth muscle cells, promoting osteoblastic differentiation [39]. Leptin may also reduce the differentiation of bone marrow stem cells into osteoclasts and stimulate the production of osteoprotegerin by mononuclear precursors, favoring an osteoblast lineage and thus osteogenesis [40].
A number of studies have reported that adiponectin levels are increased in the serum and in the synovial fluid of patients with RA compared to healthy controls or patients with OA [27][41][42][43][44][45][46][47][48]. Adiponectin levels were found to correlate with the disease activity of RA in certain studies. The synovial tissues of patients with RA, especially fibroblast-like synoviocytes, expressed adiponectin and its receptors, AdipoR1 and AdipoR2 [49]. In addition, in an experimental model, adiponectin alone or in combination with IL-1 beta may induce the release of IL-6, IL-8 and PGE2 by fibroblast synoviocytes [50]. The metalloproteinases MMP-1 and MMP-3 are also produced by fibroblast-like synoviocytes when stimulated by adiponectin [15]. Adiponectin may activate osteoclastic activation through the stimulation of RANKL and the inhibition of osteoprotegerin production by osteoblasts, thus favoring joint bone erosion [6][12]. In human chondrocytes, adiponectin stimulated the production of IL-6, IL-8, MMP-3, MMP-9 and monocyte chemoattractant protein-1 (MCP-1) [51]. It has been reported that HMW adiponectin is likely to induce a stronger pro-inflammatory reaction in RA synovial fibroblasts compared to the other isoforms [5]. Indeed, HMW adiponectin induced the secretion of IL-6 in human monocytes, while LMW adiponectin reduced IL-6 release and stimulated IL-10 secretion by LPS-activated monocytes [52]. Adiponectin is able to promote the differentiation of naïve CD4+ T lymphocytes toward a Th17 phenotype, thus contributing to joint inflammation and bone erosion. This differentiation was dependent on the AdipoR1 receptor, as demonstrated by an AdipoR1 knockout model [53]. The intra-articular injection of adiponectin in the joints of mice with collagen-induced arthritis resulted in severe disease with marked inflammation and synovial hyperplasia, RANKL expression and bone erosions [54]. Finally, it was demonstrated that the stimulation of RA synovial fibroblasts by adiponectin induced the production of osteopontin, which in turn recruited osteoclasts to the bone surface to initiate bone erosions [55]. Collectively, these data effectively demonstrate the degradative properties of adiponectin on the joint components.
Serum concentrations of visfatin were found to be higher in patients with RA, compared to healthy controls or patients with OA [27][56]. Visfatin is positively correlated with measurements of disease activity of RA [27]. Visfatin is able to induce a pro-inflammatory response in various cells, including RA fibroblast-like synoviocytes, monocytes, chondrocytes and bone cells [5]. For instance, visfatin activated human chondrocytes to produce pro-inflammatory cytokines, MMP-3 and PGE2 [3][6]. Osteoblasts may respond to visfatin by secreting IL-6 and chemokines, while conflicting results have been observed for osteoclasts, with both inhibition and activation of their differentiation [57][58]. Visfatin may potentially promote angiogenesis by increasing RA synovial fibroblast adhesion to endothelial cells under static and flow conditions [59]. In an animal model of arthritis, osteoclastogenesis required the presence of visfatin, suggesting a direct role in joint damage [60].
Resistin has been found to be moderately elevated in the sera of patients with RA, while its concentration is markedly increased in the synovial fluid compared to results obtained from subjects with OA [23][61]. A meta-analysis of eight studies of RA reported that serum resistin was higher in patients with RA compared to normal controls [62]. Resistin is expressed in the synovial fluid and the synovial membrane of patients with RA. Local cells in the joint cavity, including RA synovial fibroblasts, macrophages, B cells and osteoblasts, expressed resistin [63]. Serum resistin levels correlated with systemic biomarkers of inflammation, such as CRP or the erythrocyte sedimentation rate [64]. When stimulated by resistin, RA synovial fibroblasts produced pro-inflammatory factors and chemokines [65]. Resistin may also promote angiogenesis among endothelial progenitor cells by increasing vascular endothelial growth factor (VEGF) production [5].
2.2. Axial Spondyloarthritis
More limited information is available on adipokines in ax SpA. Conflicting results have been reported for circulating leptin levels in ankylosing spondylitis (AS) (also called radiographic ax SpA), with decreased levels in some studies [66][67] and increased levels in another [68]. Serum leptin correlated with the Bath Ankylosing Spondylitis Disease Activity Index (BASDAI), a validated patient-reported index of disease activity, and with acute-phase reactants (CRP, IL-6) [69]. The relationships between adipokines and the ankylosing spondylitis disease activity score (ASDAS), a more recent index of disease activity including acute-phase reactants, have never been examined. In a recent meta-analysis, it was reported that there was no difference in serum leptin or serum adiponectin between AS and controls, while patients with AS had higher serum resistin levels [70]. These results for serum leptin were confirmed by a second meta-analysis, indicating that there is no significant difference in plasma/serum leptin between patients and controls [71]. Certain adipokines have the potential to stimulate bone cells toward an osteogenic profile and/or to activate osteoblasts to induce bone formation, a relevant question for ax SpA. In this sense, visfatin is able to bind to osteoblasts, stimulate their proliferation and induce chemokine production and type X collagen expression [72][73]. Leptin has previously been linked to bone remodeling and thus has been implicated in the pathophysiology of osteoporosis [74]. However, the effects of leptin on bone formation remain debated, with both positive and negative consequences [75]. Indeed, leptin has effects on the central nervous system and the peripheral regulation of bone cells. On the one hand, at the central level, leptin has complex hormonal, autonomic nervous system and hypothalamic interactions, resulting in bone loss [76]. On the other hand, the peripheral effects of leptin result from direct interactions with bone cells: leptin may activate osteoblasts and inhibit osteoclasts [77][78]. Adiponectin and its receptors are expressed in osteoblasts and can stimulate their activation and differentiation. Thus, adiponectin may promote osteoblastogenesis and suppress osteoclastogenesis [79]. Conversely, adiponectin may activate osteoclasts by producing RANKL and inhibiting osteoprotegerin in osteoblasts [80]. The influence of resistin on bone remodeling is less well established. Indeed, resistin is expressed in human osteoblasts and can increase the osteoclast number and activity, but it also has a weak effect on osteoblasts by enhancing their proliferation. These data indicate a global impact of resistin on both osteoclastogenesis and osteoblastogenesis [81].
2.3. Psoriatic Arthritis
There are limited data on adipokines in patients with PsA [82][83]. High serum levels of leptin and low levels of adiponectin in patients with PsA compared to healthy controls have been reported [84]. In a large Canadian series, adipokines were compared between patients with psoriasis alone and patients with PsA. The results showed higher adiponectin levels in patients with PsA and higher levels of leptin in PsA, but only among women [85]. Finally, circulating leptin, adiponectin and resistin levels have been found to be higher in PsA than in healthy subjects [86]. The relationship between these adipokines and the levels of disease activity of PsA was not clear, either with laboratory parameters of inflammation or with clinical measurements of disease activity [84].
References
- Fantuzzi, G. Adipose tissue, adipokines, and inflammation. J. Allergy Clin. Immunol. 2005, 115, 911–919.
- Neumann, E.; Frommer, K.W.; Vasile, M.; Muller-Ladner, U. Adipocytokines as driving forces in rheumatoid arthritis and related inflammatory diseases? Arthritis. Rheum. 2011, 63, 1159–1169.
- Laurindo, L.F.; de Maio, M.C.; Barbalho, S.M.; Guiguer, E.L.; Araujo, A.C.; de Alvares Goulart, R.; Flato, U.A.P.; Junior, E.B.; Detregiachi, C.R.P.; Dos Santos Haber, J.F.; et al. Organokines in Rheumatoid Arthritis: A Critical Review. Int. J. Mol. Sci. 2022, 23, 6193.
- Toussirot, E.; Streit, G.; Wendling, D. The contribution of adipose tissue and adipokines to inflammation in joint diseases. Curr. Med. Chem. 2007, 14, 1095–1100.
- Neumann, E.; Hasseli, R.; Ohl, S.; Lange, U.; Frommer, K.W.; Muller-Ladner, U. Adipokines and Autoimmunity in Inflammatory Arthritis. Cells 2021, 10, 216.
- Neumann, E.; Junker, S.; Schett, G.; Frommer, K.; Muller-Ladner, U. Adipokines in bone disease. Nat. Rev. Rheumatol. 2016, 12, 296–302.
- Toussirot, E. Mini-Review: The Contribution of Adipokines to Joint Inflammation in Inflammatory Rheumatic Diseases. Front. Endocrinol. 2020, 11, 606560.
- Houseknecht, K.L.; Baile, C.A.; Matteri, R.L.; Spurlock, M.E. The biology of leptin: A review. J. Anim. Sci. 1998, 76, 1405–1420.
- Otero, M.; Lago, R.; Lago, F.; Casanueva, F.F.; Dieguez, C.; Gomez-Reino, J.J.; Gualillo, O. Leptin, from fat to inflammation: Old questions and new insights. FEBS Lett. 2005, 579, 295–301.
- Iikuni, N.; Lam, Q.L.; Lu, L.; Matarese, G.; La Cava, A. Leptin and Inflammation. Curr. Immunol. Rev. 2008, 4, 70–79.
- Deng, J.; Liu, Y.; Yang, M.; Wang, S.; Zhang, M.; Wang, X.; Ko, K.H.; Hua, Z.; Sun, L.; Cao, X.; et al. Leptin exacerbates collagen-induced arthritis via enhancement of Th17 cell response. Arthritis Rheum. 2012, 64, 3564–3573.
- Carrion, M.; Frommer, K.W.; Perez-Garcia, S.; Muller-Ladner, U.; Gomariz, R.P.; Neumann, E. The Adipokine Network in Rheumatic Joint Diseases. Int. J. Mol. Sci. 2019, 20, 4091.
- Ouchi, N.; Walsh, K. Adiponectin as an anti-inflammatory factor. Clin. Chim. Acta 2007, 380, 24–30.
- Okamoto, Y.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Libby, P. Adiponectin: A key adipocytokine in metabolic syndrome. Clin. Sci. 2006, 110, 267–278.
- Fatel, E.C.S.; Rosa, F.T.; Simao, A.N.C.; Dichi, I. Adipokines in rheumatoid arthritis. Adv. Rheumatol. 2018, 58, 25.
- Wang, Y.; Liu, R.; Zhao, P.; Zhang, Q.; Huang, Y.; Wang, L.; Lv, C.; Che, N.; Tan, W.; Zhang, M. Blockade of adiponectin receptor 1 signaling inhibits synovial inflammation and alleviates joint damage in collagen-induced arthritis. Clin. Rheumatol. 2022, 41, 255–264.
- Song, H.; Chan, J.; Rovin, B.H. Induction of chemokine expression by adiponectin in vitro is isoform dependent. Transl. Res. 2009, 154, 18–26.
- Bungau, S.; Behl, T.; Tit, D.M.; Banica, F.; Bratu, O.G.; Diaconu, C.C.; Nistor-Cseppento, C.D.; Bustea, C.; Aron, R.A.C.; Vesa, C.M. Interactions between leptin and insulin resistance in patients with prediabetes, with and without NAFLD. Exp. Ther. Med. 2020, 20, 197.
- Carmen Zaha, D.; Vesa, C.; Uivarosan, D.; Bratu, O.; Fratila, O.; Mirela Tit, D.; Pantis, C.; Diaconu, C.; Bungau, S. Influence of inflammation and adipocyte biochemical markers on the components of metabolic syndrome. Exp. Ther. Med. 2020, 20, 121–128.
- Krysiak, R.; Handzlik-Orlik, G.; Okopien, B. The role of adipokines in connective tissue diseases. Eur. J. Nutr. 2012, 51, 513–528.
- Norata, G.D.; Ongari, M.; Garlaschelli, K.; Raselli, S.; Grigore, L.; Catapano, A.L. Plasma resistin levels correlate with determinants of the metabolic syndrome. Eur. J. Endocrinol. 2007, 156, 279–284.
- Nagaev, I.; Bokarewa, M.; Tarkowski, A.; Smith, U. Human resistin is a systemic immune-derived proinflammatory cytokine targeting both leukocytes and adipocytes. PLoS ONE 2006, 1, e31.
- Bokarewa, M.; Nagaev, I.; Dahlberg, L.; Smith, U.; Tarkowski, A. Resistin, an adipokine with potent proinflammatory properties. J. Immunol. 2005, 174, 5789–5795.
- Meier, F.M.; Frommer, K.W.; Peters, M.A.; Brentano, F.; Lefevre, S.; Schroder, D.; Kyburz, D.; Steinmeyer, J.; Rehart, S.; Gay, S.; et al. Visfatin/pre-B-cell colony-enhancing factor (PBEF), a proinflammatory and cell motility-changing factor in rheumatoid arthritis. J. Biol. Chem. 2012, 287, 28378–28385.
- Francisco, V.; Pino, J.; Gonzalez-Gay, M.A.; Mera, A.; Lago, F.; Gomez, R.; Mobasheri, A.; Gualillo, O. Adipokines and inflammation: Is it a question of weight? Br. J. Pharmacol. 2018, 175, 1569–1579.
- Toussirot, E.; Michel, F.; Binda, D.; Dumoulin, G. The role of leptin in the pathophysiology of rheumatoid arthritis. Life Sci. 2015, 140, 29–36.
- Otero, M.; Lago, R.; Gomez, R.; Lago, F.; Dieguez, C.; Gomez-Reino, J.J.; Gualillo, O. Changes in plasma levels of fat-derived hormones adiponectin, leptin, resistin and visfatin in patients with rheumatoid arthritis. Ann. Rheum. Dis. 2006, 65, 1198–1201.
- Harle, P.; Pongratz, G.; Weidler, C.; Buttner, R.; Scholmerich, J.; Straub, R.H. Possible role of leptin in hypoandrogenicity in patients with systemic lupus erythematosus and rheumatoid arthritis. Ann. Rheum. Dis. 2004, 63, 809–816.
- Toussirot, E.; Nguyen, N.U.; Dumoulin, G.; Aubin, F.; Cedoz, J.P.; Wendling, D. Relationship between growth hormone-IGF-I-IGFBP-3 axis and serum leptin levels with bone mass and body composition in patients with rheumatoid arthritis. Rheumatology 2005, 44, 120–125.
- Tian, G.; Liang, J.N.; Pan, H.F.; Zhou, D. Increased leptin levels in patients with rheumatoid arthritis: A meta-analysis. Ir. J. Med. Sci. 2014, 183, 659–666.
- Lee, Y.H.; Bae, S.C. Circulating leptin level in rheumatoid arthritis and its correlation with disease activity: A meta-analysis. Z. Rheumatol. 2016, 75, 1021–1027.
- Anders, H.J.; Rihl, M.; Heufelder, A.; Loch, O.; Schattenkirchner, M. Leptin serum levels are not correlated with disease activity in patients with rheumatoid arthritis. Metabolism 1999, 48, 745–748.
- Muraoka, S.; Kusunoki, N.; Takahashi, H.; Tsuchiya, K.; Kawai, S. Leptin stimulates interleukin-6 production via janus kinase 2/signal transducer and activator of transcription 3 in rheumatoid synovial fibroblasts. Clin. Exp. Rheumatol. 2013, 31, 589–595.
- Rodriguez, J.; Lafaurie, G.I.; Bautista-Molano, W.; Chila-Moreno, L.; Bello-Gualtero, J.M.; Romero-Sanchez, C. Adipokines and periodontal markers as risk indicators of early rheumatoid arthritis: A cross-sectional study. Clin. Oral. Investig. 2021, 25, 1685–1695.
- Ramos-Casallas, A.; De Avila, J.; Chaparro-Sanabria, A.; Chalem-Choueka, P.; Bello-Gualtero, J.M.; Chila-Moreno, L.; Pacheco-Tena, C.; Bautista-Molano, W.; Romero-Sanchez, C. Adipokine profile on joint and periodontal conditions in first-degree relatives of patients with rheumatoid arthritis. Curr. Rheumatol. Rev. 2022.
- Gomez, R.; Scotece, M.; Conde, J.; Gomez-Reino, J.J.; Lago, F.; Gualillo, O. Adiponectin and leptin increase IL-8 production in human chondrocytes. Ann. Rheum. Dis. 2011, 70, 2052–2054.
- Yaykasli, K.O.; Hatipoglu, O.F.; Yaykasli, E.; Yildirim, K.; Kaya, E.; Ozsahin, M.; Uslu, M.; Gunduz, E. Leptin induces ADAMTS-4, ADAMTS-5, and ADAMTS-9 genes expression by mitogen-activated protein kinases and NF-kB signaling pathways in human chondrocytes. Cell. Biol. Int. 2015, 39, 104–112.
- Yang, W.H.; Tsai, C.H.; Fong, Y.C.; Huang, Y.L.; Wang, S.J.; Chang, Y.S.; Tang, C.H. Leptin induces oncostatin M production in osteoblasts by downregulating miR-93 through the Akt signaling pathway. Int. J. Mol. Sci. 2014, 15, 15778–15790.
- Liu, G.Y.; Liang, Q.H.; Cui, R.R.; Liu, Y.; Wu, S.S.; Shan, P.F.; Yuan, L.Q.; Liao, E.Y. Leptin promotes the osteoblastic differentiation of vascular smooth muscle cells from female mice by increasing RANKL expression. Endocrinology 2014, 155, 558–567.
- Cirmanova, V.; Bayer, M.; Starka, L.; Zajickova, K. The effect of leptin on bone: An evolving concept of action. Physiol. Res. 2008, 57, S143–S151.
- Toussirot, E.; Grandclement, E.; Gaugler, B.; Michel, F.; Wendling, D.; Saas, P.; Dumoulin, G.; Cbt. Serum adipokines and adipose tissue distribution in rheumatoid arthritis and ankylosing spondylitis. A comparative study. Front. Immunol. 2013, 4, 453.
- Rho, Y.H.; Solus, J.; Sokka, T.; Oeser, A.; Chung, C.P.; Gebretsadik, T.; Shintani, A.; Pincus, T.; Stein, C.M. Adipocytokines are associated with radiographic joint damage in rheumatoid arthritis. Arthritis Rheum. 2009, 60, 1906–1914.
- Giles, J.T.; Allison, M.; Bingham, C.O., 3rd; Scott, W.M., Jr.; Bathon, J.M. Adiponectin is a mediator of the inverse association of adiposity with radiographic damage in rheumatoid arthritis. Arthritis Rheum. 2009, 61, 1248–1256.
- Giles, J.T.; van der Heijde, D.M.; Bathon, J.M. Association of circulating adiponectin levels with progression of radiographic joint destruction in rheumatoid arthritis. Ann. Rheum. Dis. 2011, 70, 1562–1568.
- Klein-Wieringa, I.R.; van der Linden, M.P.; Knevel, R.; Kwekkeboom, J.C.; van Beelen, E.; Huizinga, T.W.; van der Helm-van Mil, A.; Kloppenburg, M.; Toes, R.E.; Ioan-Facsinay, A. Baseline serum adipokine levels predict radiographic progression in early rheumatoid arthritis. Arthritis Rheum. 2011, 63, 2567–2574.
- Senolt, L.; Pavelka, K.; Housa, D.; Haluzik, M. Increased adiponectin is negatively linked to the local inflammatory process in patients with rheumatoid arthritis. Cytokine 2006, 35, 247–252.
- Ebina, K.; Fukuhara, A.; Ando, W.; Hirao, M.; Koga, T.; Oshima, K.; Matsuda, M.; Maeda, K.; Nakamura, T.; Ochi, T.; et al. Serum adiponectin concentrations correlate with severity of rheumatoid arthritis evaluated by extent of joint destruction. Clin. Rheumatol. 2009, 28, 445–451.
- Ozgen, M.; Koca, S.S.; Dagli, N.; Balin, M.; Ustundag, B.; Isik, A. Serum adiponectin and vaspin levels in rheumatoid arthritis. Arch. Med. Res. 2010, 41, 457–463.
- Tan, W.; Wang, F.; Zhang, M.; Guo, D.; Zhang, Q.; He, S. High adiponectin and adiponectin receptor 1 expression in synovial fluids and synovial tissues of patients with rheumatoid arthritis. Semin. Arthritis Rheum. 2009, 38, 420–427.
- Chen, X.; Lu, J.; Bao, J.; Guo, J.; Shi, J.; Wang, Y. Adiponectin: A biomarker for rheumatoid arthritis? Cytokine Growth Factor Rev. 2013, 24, 83–89.
- Szumilas, K.; Szumilas, P.; Sluczanowska-Glabowska, S.; Zgutka, K.; Pawlik, A. Role of Adiponectin in the Pathogenesis of Rheumatoid Arthritis. Int. J. Mol. Sci. 2020, 21, 8265.
- Neumeier, M.; Weigert, J.; Schaffler, A.; Wehrwein, G.; Muller-Ladner, U.; Scholmerich, J.; Wrede, C.; Buechler, C. Different effects of adiponectin isoforms in human monocytic cells. J. Leukoc. Biol. 2006, 79, 803–808.
- Zhang, Q.; Wang, L.; Jiang, J.; Lin, S.; Luo, A.; Zhao, P.; Tan, W.; Zhang, M. Critical Role of AdipoR1 in Regulating Th17 Cell Differentiation Through Modulation of HIF-1alpha-Dependent Glycolysis. Front. Immunol. 2020, 11, 2040.
- Sun, X.; Feng, X.; Tan, W.; Lin, N.; Hua, M.; Wei, Y.; Wang, F.; Li, N.; Zhang, M. Adiponectin exacerbates collagen-induced arthritis via enhancing Th17 response and prompting RANKL expression. Sci. Rep. 2015, 5, 11296.
- Qian, J.; Xu, L.; Sun, X.; Wang, Y.; Xuan, W.; Zhang, Q.; Zhao, P.; Wu, Q.; Liu, R.; Che, N.; et al. Adiponectin aggravates bone erosion by promoting osteopontin production in synovial tissue of rheumatoid arthritis. Arthritis Res. Ther. 2018, 20, 26.
- Lee, Y.H.; Bae, S.C. Circulating adiponectin and visfatin levels in rheumatoid arthritis and their correlation with disease activity: A meta-analysis. Int. J. Rheum. Dis. 2018, 21, 664–672.
- Moschen, A.R.; Geiger, S.; Gerner, R.; Tilg, H. Pre-B cell colony enhancing factor/NAMPT/visfatin and its role in inflammation-related bone disease. Mutat. Res. 2010, 690, 95–101.
- Venkateshaiah, S.U.; Khan, S.; Ling, W.; Bam, R.; Li, X.; van Rhee, F.; Usmani, S.; Barlogie, B.; Epstein, J.; Yaccoby, S. NAMPT/PBEF1 enzymatic activity is indispensable for myeloma cell growth and osteoclast activity. Exp. Hematol. 2013, 41, 547–557.e2.
- Hasseli, R.; Frommer, K.W.; Schwarz, M.; Hulser, M.L.; Schreiyack, C.; Arnold, M.; Diller, M.; Tarner, I.H.; Lange, U.; Pons-Kuhnemann, J.; et al. Adipokines and Inflammation Alter the Interaction Between Rheumatoid Arthritis Synovial Fibroblasts and Endothelial Cells. Front. Immunol. 2020, 11, 925.
- Li, X.; Islam, S.; Xiong, M.; Nsumu, N.N.; Lee, M.W., Jr.; Zhang, L.Q.; Ueki, Y.; Heruth, D.P.; Lei, G.; Ye, S.Q. Epigenetic regulation of NfatC1 transcription and osteoclastogenesis by nicotinamide phosphoribosyl transferase in the pathogenesis of arthritis. Cell Death Discov. 2019, 5, 62.
- Schaffler, A.; Ehling, A.; Neumann, E.; Herfarth, H.; Tarner, I.; Scholmerich, J.; Muller-Ladner, U.; Gay, S. Adipocytokines in synovial fluid. JAMA 2003, 290, 1709–1710.
- Huang, Q.; Tao, S.S.; Zhang, Y.J.; Zhang, C.; Li, L.J.; Zhao, W.; Zhao, M.Q.; Li, P.; Pan, H.F.; Mao, C.; et al. Serum resistin levels in patients with rheumatoid arthritis and systemic lupus erythematosus: A meta-analysis. Clin. Rheumatol. 2015, 34, 1713–1720.
- Senolt, L.; Housa, D.; Vernerova, Z.; Jirasek, T.; Svobodova, R.; Veigl, D.; Anderlova, K.; Muller-Ladner, U.; Pavelka, K.; Haluzik, M. Resistin in rheumatoid arthritis synovial tissue, synovial fluid and serum. Ann. Rheum. Dis. 2007, 66, 458–463.
- Yoshino, T.; Kusunoki, N.; Tanaka, N.; Kaneko, K.; Kusunoki, Y.; Endo, H.; Hasunuma, T.; Kawai, S. Elevated serum levels of resistin, leptin, and adiponectin are associated with C-reactive protein and also other clinical conditions in rheumatoid arthritis. Intern. Med. 2011, 50, 269–275.
- Sato, H.; Muraoka, S.; Kusunoki, N.; Masuoka, S.; Yamada, S.; Ogasawara, H.; Imai, T.; Akasaka, Y.; Tochigi, N.; Takahashi, H.; et al. Resistin upregulates chemokine production by fibroblast-like synoviocytes from patients with rheumatoid arthritis. Arthritis Res. Ther. 2017, 19, 263.
- Sari, I.; Demir, T.; Kozaci, L.D.; Akar, S.; Kavak, T.; Birlik, M.; Onen, F.; Akkoc, N. Body composition, insulin, and leptin levels in patients with ankylosing spondylitis. Clin. Rheumatol. 2007, 26, 1427–1432.
- Toussirot, E.; Streit, G.; Nguyen, N.U.; Dumoulin, G.; Le Huede, G.; Saas, P.; Wendling, D. Adipose tissue, serum adipokines, and ghrelin in patients with ankylosing spondylitis. Metabolism 2007, 56, 1383–1389.
- Park, M.C.; Chung, S.J.; Park, Y.B.; Lee, S.K. Pro-inflammatory effect of leptin on peripheral blood mononuclear cells of patients with ankylosing spondylitis. Jt. Bone Spine 2009, 76, 170–175.
- Park, M.C.; Lee, S.W.; Choi, S.T.; Park, Y.B.; Lee, S.K. Serum leptin levels correlate with interleukin-6 levels and disease activity in patients with ankylosing spondylitis. Scand. J. Rheumatol. 2007, 36, 101–106.
- Yang, J.; Zhang, X.; Ma, Y.; Wu, M.; Hu, X.; Han, R.; Yuan, Y.; Wang, M.; Chen, M.; Jiang, S.; et al. Serum levels of leptin, adiponectin and resistin in patients with ankylosing spondylitis: A systematic review and meta-analysis. Int. Immunopharmacol. 2017, 52, 310–317.
- Mei, Y.J.; Wang, P.; Chen, L.J.; Li, Z.J. Plasma/Serum Leptin Levels in Patients with Ankylosing Spondylitis: A Systematic Review and Meta-analysis. Arch. Med. Res. 2016, 47, 111–117.
- Xie, H.; Tang, S.Y.; Luo, X.H.; Huang, J.; Cui, R.R.; Yuan, L.Q.; Zhou, H.D.; Wu, X.P.; Liao, E.Y. Insulin-like effects of visfatin on human osteoblasts. Calcif. Tissue Int. 2007, 80, 201–210.
- Laiguillon, M.C.; Houard, X.; Bougault, C.; Gosset, M.; Nourissat, G.; Sautet, A.; Jacques, C.; Berenbaum, F.; Sellam, J. Expression and function of visfatin (Nampt), an adipokine-enzyme involved in inflammatory pathways of osteoarthritis. Arthritis Res. Ther. 2014, 16, R38.
- Karsenty, G.; Khosla, S. The crosstalk between bone remodeling and energy metabolism: A translational perspective. Cell. Metab. 2022, 34, 805–817.
- Chen, X.X.; Yang, T. Roles of leptin in bone metabolism and bone diseases. J. Bone. Miner. Metab. 2015, 33, 474–485.
- Ducy, P.; Amling, M.; Takeda, S.; Priemel, M.; Schilling, A.F.; Beil, F.T.; Shen, J.; Vinson, C.; Rueger, J.M.; Karsenty, G. Leptin inhibits bone formation through a hypothalamic relay: A central control of bone mass. Cell 2000, 100, 197–207.
- Reseland, J.E.; Syversen, U.; Bakke, I.; Qvigstad, G.; Eide, L.G.; Hjertner, O.; Gordeladze, J.O.; Drevon, C.A. Leptin is expressed in and secreted from primary cultures of human osteoblasts and promotes bone mineralization. J. Bone Miner. Res. 2001, 16, 1426–1433.
- Holloway, W.R.; Collier, F.M.; Aitken, C.J.; Myers, D.E.; Hodge, J.M.; Malakellis, M.; Gough, T.J.; Collier, G.R.; Nicholson, G.C. Leptin inhibits osteoclast generation. J. Bone Miner. Res. 2002, 17, 200–209.
- Luo, X.H.; Guo, L.J.; Yuan, L.Q.; Xie, H.; Zhou, H.D.; Wu, X.P.; Liao, E.Y. Adiponectin stimulates human osteoblasts proliferation and differentiation via the MAPK signaling pathway. Exp. Cell. Res. 2005, 309, 99–109.
- Luo, X.H.; Guo, L.J.; Xie, H.; Yuan, L.Q.; Wu, X.P.; Zhou, H.D.; Liao, E.Y. Adiponectin stimulates RANKL and inhibits OPG expression in human osteoblasts through the MAPK signaling pathway. J. Bone Miner. Res. 2006, 21, 1648–1656.
- Thommesen, L.; Stunes, A.K.; Monjo, M.; Grosvik, K.; Tamburstuen, M.V.; Kjobli, E.; Lyngstadaas, S.P.; Reseland, J.E.; Syversen, U. Expression and regulation of resistin in osteoblasts and osteoclasts indicate a role in bone metabolism. J. Cell. Biochem. 2006, 99, 824–834.
- Russolillo, A.; Iervolino, S.; Peluso, R.; Lupoli, R.; Di Minno, A.; Pappone, N.; Di Minno, M.N. Obesity and psoriatic arthritis: From pathogenesis to clinical outcome and management. Rheumatology 2013, 52, 62–67.
- Toussirot, E.; Aubin, F.; Desmarets, M.; Wendling, D.; Auge, B.; Gillard, J.; Messica, O.; Guillot, X.; Laheurte, C.; Monnet, E.; et al. Visceral adiposity in patients with psoriatic arthritis and psoriasis alone and its relationship with metabolic and cardiovascular risk. Rheumatology 2021, 60, 2816–2825.
- Xue, Y.; Jiang, L.; Cheng, Q.; Chen, H.; Yu, Y.; Lin, Y.; Yang, X.; Kong, N.; Zhu, X.; Xu, X.; et al. Adipokines in psoriatic arthritis patients: The correlations with osteoclast precursors and bone erosions. PLoS ONE 2012, 7, e46740.
- Eder, L.; Jayakar, J.; Pollock, R.; Pellett, F.; Thavaneswaran, A.; Chandran, V.; Rosen, C.F.; Gladman, D.D. Serum adipokines in patients with psoriatic arthritis and psoriasis alone and their correlation with disease activity. Ann. Rheum. Dis. 2013, 72, 1956–1961.
- Dikbas, O.; Tosun, M.; Bes, C.; Tonuk, S.B.; Aksehirli, O.Y.; Soy, M. Serum levels of visfatin, resistin and adiponectin in patients with psoriatic arthritis and associations with disease severity. Int. J. Rheum. Dis. 2016, 19, 672–677.
More
Information
Subjects:
Rheumatology
Contributor
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
783
Revisions:
2 times
(View History)
Update Date:
15 Mar 2023
Table of Contents




Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No