Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Ion Udroiu | -- | 2706 | 2022-12-19 11:26:25 | | | |
| 2 | Conner Chen | Meta information modification | 2706 | 2022-12-20 11:22:48 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Udroiu, I.; Marinaccio, J.; Sgura, A. Function and Components of Telomerase. Encyclopedia. Available online: https://encyclopedia.pub/entry/38960 (accessed on 24 July 2026).
Udroiu I, Marinaccio J, Sgura A. Function and Components of Telomerase. Encyclopedia. Available at: https://encyclopedia.pub/entry/38960. Accessed July 24, 2026.
Udroiu, Ion, Jessica Marinaccio, Antonella Sgura. "Function and Components of Telomerase" Encyclopedia, https://encyclopedia.pub/entry/38960 (accessed July 24, 2026).
Udroiu, I., Marinaccio, J., & Sgura, A. (2022, December 19). Function and Components of Telomerase. In Encyclopedia. https://encyclopedia.pub/entry/38960
Udroiu, Ion, et al. "Function and Components of Telomerase." Encyclopedia. Web. 19 December, 2022.
Copy Citation
Telomerase is the only known eukaryotic-specific enzyme with reverse transcriptase activity, which adds telomeric repeats at the ends of linear chromosomes. In this way, it counteracts telomere shortening and cellular replicative senescence. Telomerase consists of a catalytic protein subunit with reverse transcriptase activity (TERT), and an essential RNA component known as telomerase RNA component (TERC) that contains a template for the synthesis of telomeric DNA, as well as additional proteins (dyskerin, NHP2, NOP10 and GAR1 in vertebrates) that play crucial roles in its biogenesis, localization, and regulation. Beside its telomere-elongating activity, a growing number of studies have evidenced non-telomeric functions.
telomerase
gene regulation
mitochondria
apoptosis
1. Introduction
Since its discovery by Greider and Blackburn [1] (preceded by the prediction of its existence by Olovnikov [2]), telomerase has attracted the attention of many researchers, probably because of its unique, distinctive features: it is the only known eukaryotic-specific enzyme with reverse transcriptase activity, and it counteracts replicative senescence, potentially allowing cells to be immortal. Moreover, the idea that “normal” cells do not express telomerase and tumor cells do prompted the idea that a specific target to kill only cancer cells (without harming the normal ones) had been found [3]. The real picture, however, is far more complex, since many somatic cells express telomerase: not only hematopoietic stem and progenitor cells [4], but also many types of epithelial cells, such as keratinocytes [5] and lens epithelial cells [6].
2. Telomerase: Function and Components
2.1. Telomerase Function
Telomerase is a reverse transcriptase that adds telomeric repeats at the 3′ ends of linear chromosomes [1]. These sequences are TTAGGG in most eukaryotes (and most probably represent the ancestral motif) but show many variations in several lineages [7]. Telomerase activity, through telomere elongation, helps to maintain genome stability, preventing chromosome ends from being recognized and processed as DNA double strand breaks [8].
Telomerase consists of a catalytic protein subunit with reverse transcriptase activity (TERT), and an essential RNA component known as telomerase RNA component (TERC) that contains a template for the synthesis of telomeric DNA [9]. The telomerase holoenzyme also contains additional proteins (dyskerin, NHP2, NOP10 and GAR1 in vertebrates) that play crucial roles in its biogenesis, localization, and regulation [10][11].
The telomerase catalytic cycle comprises two main phases: the synthesis of a single telomere repeat, and the repositioning of the template for the synthesis of additional repeats. More precisely, it includes four stages: primer binding, elongation, translocation and dissociation [8].
The reaction starts by binding the DNA primer at the 3′ end of the telomere with the 5′ region of the RNA template of telomerase, which results in the formation of a short hybrid DNA/RNA duplex and then proceeds with the synthesis of one telomeric repeat. Once the repeat is completed, nucleotide addition arrests, followed by translocation of the RNA template. The translocation is a complex multi-step process and it has been shown to occur outside the active site [12]. Through this process, a single primer can be extended with numerous telomere repeats before complete disassociation from the telomerase enzyme [13][14].
For telomerase activity, TERT is usually considered the limiting factor, because in humans it is repressed in most somatic tissues, while TERC is constitutively expressed [15]. It is true that tissues with no TERT expression show no telomerase activity (and not only in humans). However, while databanks for the expression of TERT in different types of cells and tissues are abundant, the same cannot be said for TERC (as for many other ncRNAs). Thus, so far, it is difficult to have a clear picture of cell- and tissue-specific TERC expression. Moreover, Avillon et al. [16] stated that TERC expression is a bad predictor of telomerase activity (while TERT expression is a good one), but they also found that TERC is expressed at very low levels in human normal brain, muscle and lung tissues and at low levels in breast and liver. More recently, Castle et al. [17] measured the expression of TERC in different human tissues. Moreover, Hartmann et al. [18] measured TERT and TERC expression in different tissues of the turquoise killifish (Nothobranchius furzeri), finding the same trend for both and their agreement with telomerase activity. Finally, it was also shown that TERC upregulation is an essential characteristic for telomerase induction in induced pluripotent cells [19]. Therefore, it can be summed up that TERC is expressed in more human tissues than TERT and in higher abundance, but also TERC expression shows significant differences between tissues and cell types.
2.2. TERT Structure
The TERT protein is the catalytic component of the core of telomerase enzymes. This protein is composed of four conserved structural domains (Figure 1): the telomerase essential N-terminal domain (TEN), the telomerase RNA binding domain (TRBD), a central catalytic RT domain, and C-terminal extension (CTE).
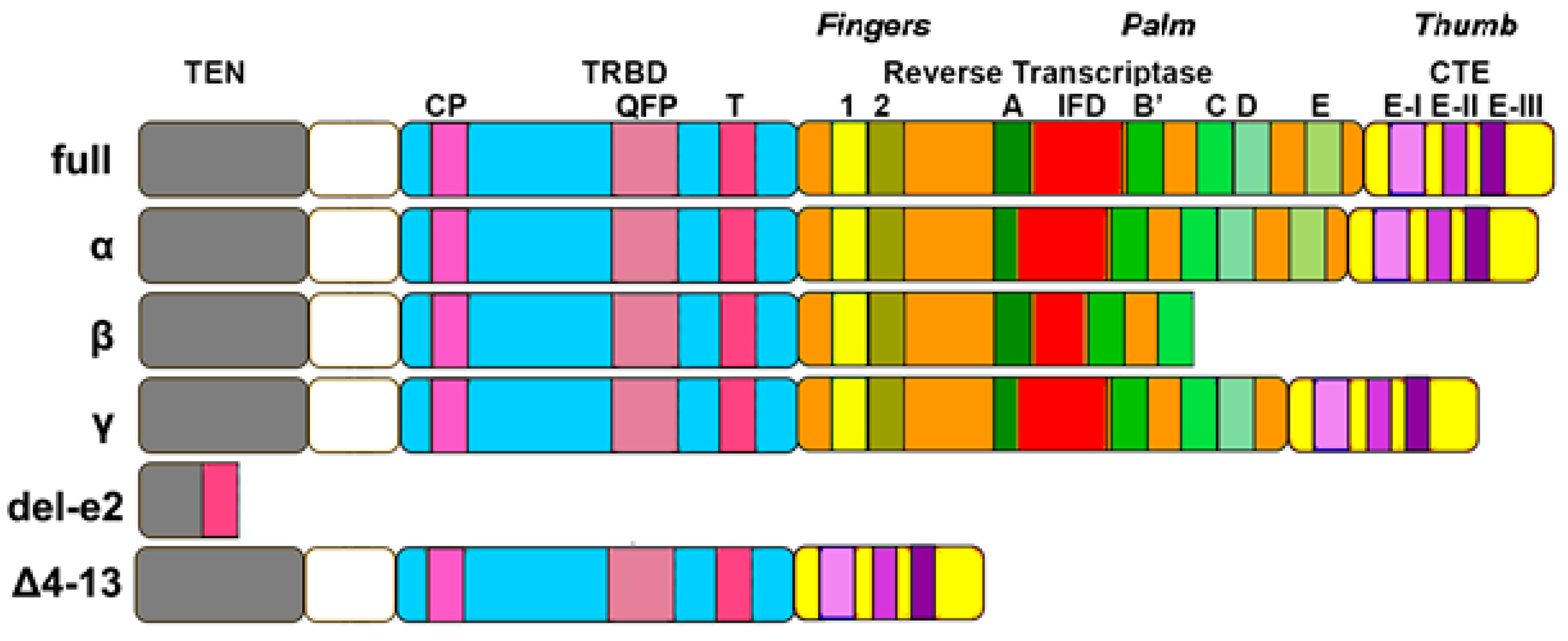
Figure 1. Architecture of of human TERT and alternative isoforms. TEN: telomerase essential N-terminal domain; TRBD: telomerase RNA binding domain; CTE: C-terminal extension. Isoform α shows a partial loss of Reverse Transcriptase-motif A. Isoform β shows partial loss and sequence variation of the IFD domain, as well as loss of the D and E fingers motifs and CTE domain. Isoform γ is similar to α, but also shows the loss of the E finger motif. Del-e2 is composed only of part of the TEN domain and T-motif (Figure by Ion Udroiu).
The TEN domain is necessary for telomerase functioning in vitro and in vivo. The N-terminal contains several conserved telomerase-specific motifs, which are important for TERT–TERC binding interactions and for the rate of template copying during telomere synthesis [20][21][22]. This domain also contains ‘anchor’ sites that bind single-stranded telomeric DNA [23], allowing the complete delay disassociation of the DNA product from the enzyme and increasing repeat addition processivity. The TEN domains include RNA interacting domain 1 (RID 1), with a low-affinity binding site for the RT template/pseudoknot domain [24].
TRBD domain is unique to TERT protein, conferring the ability to use internal RNA templates upon addition of telomeric repeats. In addition, it contains RNA interacting domain 2 (RID 2) which has a high affinity for the three-way junction domain of TERC [25]. The protein-RNA interactions through RID1 and RID2 are essential for telomerase assembly [26]. The TRBD domain contains mainly helical motifs, including the CP, QFP and T motifs that participate in RNA binding [27][28].
The catalytic domain of TERT is the most characterized region of the protein and contains seven evolutionary conserved RT domains, essential for its enzymatic activities [20][29]. This domain has seven motifs: 1, 2, and A, B, C, D and E. The RT domain is described as a right hand divided in two subdomain that resemble the ‘fingers’ and ‘palm’ [30]. The finger domain helps to bind incoming nucleotides, while the palm represents the catalytic site [31]. Within the RT domain, a telomeric-specific motif (motif 3) is specific to high repeat addition processivity [32].
The CTE domain represents the so-called “thumb”, constituted by a helical bundle containing several surface-exposed loops that contributes to the formation and stabilization of an RNA-DNA heteroduplex in the enzyme active site [33]. Moreover, a nuclear export signal (NES) motif is present in these domains that allows TERT to exit through nuclear pores [34][35].
2.3. TERC Structure
A unique feature of telomerase is that the RNA template for DNA synthesis is an integral component of the holoenzyme. Being an ncRNA, TERC shows very little sequence conservation between species. In fact, mutations in ncRNA genes are not constrained by the genetic code and what is conserved is the secondary structure and not the sequence. Therefore, TERC is extremely difficult to find, even in not-so-distant species. So far, it has been found in ciliates [36], Plasmodium [37], land plants [38], Ascomycota [39], and metazoans [40].
Despite no sequence conservation, TERC shows two domains conserved in all species: a pseudoknot/template core domain and a three-way junction domain [41]. Ciliate telomerase RNA, the simplest and shortest one, comprises only those. In yeasts (with the longest sequences), several other conserved domains are present, one of them binding to Ku and another to Sm proteins [39]. In echinoderms and vertebrates (Figure 2), the third one is the box H/ACA (typical of small Cajal body-specific RNAs, scaRNAs) domain [42].
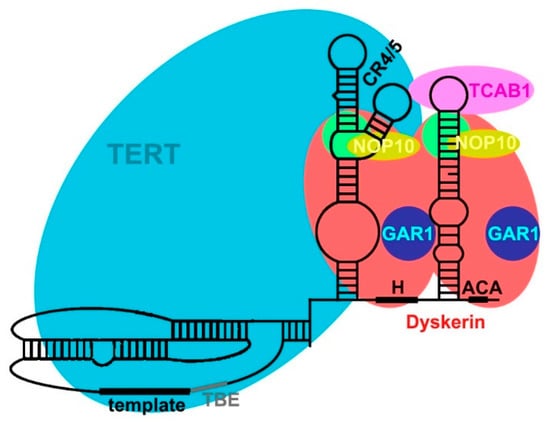
Figure 2. Human telomerase holoenzyme. TERC (structure in black) comprises the pseudoknot/template core domain (including template and template boundary element, TBE) and the CR4/5 domain (three-way junction), which are associated with TERT. In the scaRNA domain, two dyskerin molecules are bound to TERC through the H and ACA boxes. NHP2 (in green) and TCAB1 are also bound to TERC, while NOP10 and GAR1 are bound to dyskerin (Figure by Ion Udroiu).
The core domain is essential for telomerase activity in vitro and in vivo [43]. This region contains the template for telomere addition, the 5′ boundary element, TERT binding site and a conserved pseudoknot structure [42][44]. The template sequences can be divided into a 5′ region encoding for telomeric DNA repeats and a 3′ region annealing to the DNA primer after template translocation. In addition, the template is near, on the 5′ end, to the template boundary element (TBE) that defines the end of the templating sequence (Figure 2). The pseudoknot contains a triple helix and the loss or disruption of this structure drastically reduces telomerase activity [45][46]. It is thought that its conformation is involved in the translocation of the template [12], which happens while the binding between TERT and the telomere is maintained. In summary, the TERC template/pseudoknot domain has a complex structure, necessary for the correct functioning of the holoenzyme [44].
A domain shared by all studied TERCs is a three-way junction distal to the template/pseudoknot domain, which is indispensable for enzymatic activity [47][48]. In animals [40], it is composed by the CR4/5 domain (Figure 2). With its L-shaped three-way-junction conformation, its two arms clamp onto the TRBD domain of TERT [49].
In animals [40], TERC has a conserved H/ACA domain located at the 3′ end (Figure 2). The H/ACA domain contains two stem-loops separated by a conserved H box that is located at the hinge region. The ACA box is located at the 3′-end and serves as binding site for dyskerin [50][51][52]. The bulge of the 3′ stem-loop of the H/ACA domain shows a Cajal body localization (CAB box) moiety, allowing the binding of TCAB1 protein [10][50][53]. This domain is essential for TERC stability, processing, nuclear localization and telomerase activity in vivo [43][54][55].
2.4. Secondary and Accessory Proteins
TERT binds only to TERC and to telomeres (including both telomeric DNA and telomeric proteins). TERC, instead, binds to different accessory proteins, essential to its localization, maturation, 3′ processing and ribonucleoprotein biogenesis [56]. TERC, through two stems present in the H/ACA domains (Figure 2), interacts with the protein complex formed by dyskerin, NOP10, NHP2, and GAR1 proteins [57].
Dyskerin is a nucleolar protein, fundamental for many cellular processes. The main function of dyskerin is to bind scaRNAs, thanks to the H and ACA boxes [58]. Dyskerin catalyzes the pseudouridylation of specific residues in ribosomal RNA and in small nuclear RNA [59][60][61][62]. In association with NHP2 and NOP10 (Figure 2), it forms a core trimer that directly binds to the H/ACA domain of TERC and regulates its stability [63]. GAR1, instead, binds only to dyskerin (Figure 2).
GAR1 is characterized by glycine–arginine-rich domains and is required for pre-rRNA processing [64]. NOP10 is characterized by a zinc ribbon domain in the N-terminal region [53]. Both NOP10 and GAR1 bind to dyskerin and are not directly bound to TERC [53][65]. Conversely, NHP2 is an RNA binding protein, but the specificity for the binding of H/ACA scaRNAs by NHP2 comes through its association with dyskerin via the small intermediate protein NOP10 [62].
Telomerase Cajal body protein 1 (TCAB1 or WRAP53) binds TERC for localization in the Cajal bodies, where the telomerase biogenesis to generate an active telomerase complex occurs [10][66].
Other proteins interact, temporarily, with telomerase; Hsp90, p23, pontin, reptin and others are required for the assembly of the telomerase complex and its translocation from the cytosol to the nucleus [67].
2.5. Has Telomerase Any Non-Telomeric Function?
2.5.1. Addition of Telomeric Repeats at Double-Strand Break Sites?
As said before, dozens of reviews have discussed the extra-telomeric functions of “telomerase”. In reality, all these articles are about TERT, and not about the telomerase holoenzyme. The only known putative extra-telomeric function of this enzyme is its activity (addition of telomeric repeats) at double strand break (DSB) sites [68]. This is often cited in studies on karyotype evolution to justify the presence of interstitial telomeric sequences (ITS) [69]. The article by Flint et al. [68], however, did not investigate telomerase presence or activity at DSB sites, but studied telomeric sequences present on truncation breakpoints on the 16p chromosome arm of six patients. Their conclusion is based on the fact that, in five patients, the three or four nucleotides preceding the breakpoint are complementary to the TERC template. However, how can telomerase recognize a broken chromosome? Indeed, telomerase is recruited to telomeres by TPP1 [70], which, so far, is known to be present only in the shelterin complex.
The reason for the presence of telomeric repeats at “truncated” chromosomes is that, in reality, these are not truncated, i.e., the result of terminal deletions, but represent interstitial deletions. This hypothesis was already made by Furuya et al. [71] in order to explain the presence of telomeric sequences on the deleted short arms of chromosomes 9 and 10 in promyelocytic cell line HL-60. Moreover, Meltzer et al. [72] demonstrated that apparent terminal deletions characteristic of tumor cells and syndromes such as Miller–Dieker and Wolf–Hirschhorn are, in reality, subtelomeric translocations which were undetectable using conventional cytogenetics. These phenomena could be experimentally reproduced by X-irradiating fibroblasts (which do not express telomerase) and investigating whether or not some “truncated” chromosomes in reality show telomeric sequences (by Fluorescence In Situ Hybridization with a telomeric probe).
Nonetheless, telomeric sequences addition at broken chromosomes has been reported in a variety of species. McClintock [73] observed that dicentric-derived broken chromosomes acquire telomeres in maize. Fan and Yao [74] observed telomere formation during programmed chromosome breakage in Tetrahymena thermophila, and this was not dependent on the presence of a telomeric sequence at the break site. Matsumoto et al. [75] found novel telomeres on minichromosomes obtained by gamma-ray cleavage in the fission yeast Schizosaccharomyces pombe; they excluded the possibility of a recombinational event because “their chromosomal counterparts showed no sign of gross rearrangement”. Moreover, in the budding yeast Saccharomyces cerevisiae, Kramer and Haber [76] observed the de novo addition of telomeric sequence, but only when a telomeric sequence was present proximal to the break site, suggesting that this was needed as a primer for telomerase. Since they used an RAD52-deficeint strain, they concluded that recombination was impossible. However, in this same species, Wang and Zakian [77] observed telomere acquisition through a “novel recombination process involving a gene conversion event that requires little homology, occurs at or near the boundary of telomeric and nontelomeric DNA”; they suggested that this non-reciprocal recombination can also happen at non-telomeric ends, either through bypass or digestion until a telomere-like sequence. It should be added that Break-Induced Replication (BIR), one mode of action of ALT, can be both RAD52-dependent and -independent [78]. Therefore, in some of the species listed above, de novo telomere addition could be due to ALT and not telomerase activity. In any case, all these species being so phylogenetically far from vertebrates, whether or not telomerase activity at DSB is really possible in human cells should be investigated, as DSB, lacking TPP1, seems unable to recruit telomerase.
It should be noted, however, that the article by Flint et al. [68] was not about ITS, but about de novo telomeric sequences at truncated chromosomes. On an evolutionary timescale, these could fuse with other telomeric ends and give place to ITS, but this can also happen with normal telomeres without the need to find new mechanisms, involving telomerase or not.
2.5.2. NOP2-Dependent Recruitment of Telomerase to Cyclin D1 Promoter
Recently, Hong et al. [79] identified NOP2 as a new TERC-binding protein found in catalytically active telomerase. They found that telomerase is recruited to the cyclin D1 (CCND1) promoter in a TERC-dependent manner through the interaction with NOP2, enhancing transcription of this gene, whereas TERT alone (i.e., in a TERC- and NOP2-independent manner) binds to the promoter of Myc [79]. The authors also provided a hypothesis, according to which the assembled telomerase complex can comprise either TCAB1 or NOP2. In the first case, telomerase is recruited to telomeres for their elongation; in the second one, it binds to the CCND1 promoter. In any case, this mechanism of cyclin D1 activation is different from all the other cases of gene regulation, which are exerted by TERT without any other telomerase component, and is the only case (so far revealed) of extra-telomeric function of telomerase.
References
- Greider, C.W.; Blackburn, E.H. Identification of a Specific Telomere Terminal Transferase Activity in Tetrahymena Extracts. Cell 1985, 43, 405–413.
- Olovnikov, A.M. Principle of Marginotomy in Template Synthesis of Polynucleotides. Dokl. Akad. Nauk SSSR 1971, 201, 1496–1499.
- Shay, J.W.; Wright, W.E. Telomerase: A Target for Cancer Therapeutics. Cancer Cell 2002, 2, 257–265.
- Morrison, S.J.; Prowse, K.R.; Ho, P.; Weissman, I.L. Telomerase Activity in Hematopoietic Cells Is Associated with Self-Renewal Potential. Immunity 1996, 5, 207–216.
- Härle-Bachor, C.; Boukamp, P. Telomerase Activity in the Regenerative Basal Layer of the Epidermis Inhuman Skin and in Immortal and Carcinoma-Derived Skin Keratinocytes. Proc. Natl. Acad. Sci. USA 1996, 93, 6476–6481.
- Colitz, C.M.H.; Davidson, M.G.; McGahan, M.C. Telomerase Activity in Lens Epithelial Cells of Normal and Cataractous Lenses. Exp. Eye Res. 1999, 69, 641–649.
- Udroiu, I.; Russo, V.; Persichini, T.; Colasanti, M.; Sgura, A. Telomeres and Telomerase in Basal Metazoa. Invertebr. Surviv. J. 2017, 14, 233–240.
- Autexier, C.; Lue, N.F. The Structure and Function of Telomerase Reverse Transcriptase. Annu. Rev. Biochem. 2006, 75, 493–517.
- Greider, C.W.; Blackburn, E.H. A Telomeric Sequence in the RNA of Tetrahymena Telomerase Required for Telomere Repeat Synthesis. Nature 1989, 337, 331–337.
- Venteicher, A.S.; Artandi, S.E. TCAB1: Driving Telomerase to Cajal Bodies. Cell Cycle 2009, 8, 1329–1331.
- Hockemeyer, D.; Collins, K. Control of Telomerase Action at Human Telomeres. Nat. Struct. Mol. Biol. 2015, 22, 848–852.
- Zhang, Q.; Kim, N.K.; Feigon, J. Architecture of Human Telomerase RNA. Proc. Natl. Acad. Sci. USA 2011, 108, 20325–20332.
- Morin, G.B. The Human Telomere Terminal Transferase Enzyme Is a Ribonucleoprotein That Synthesizes TTAGGG Repeats. Cell 1989, 59, 521–529.
- Greider, C.W. Telomerase Is Processive. Mol. Cell. Biol. 1991, 11, 4572–4580.
- Weise, J.M.; Günes, C. Differential Regulation of Human and Mouse Telomerase Reverse Transcriptase (TERT) Promoter Activity during Testis Development. Mol. Reprod. Dev. 2009, 76, 309–317.
- Avilion, A.A.; Piatyszek, M.A.; Gupta, J.; Shay, J.W.; Bacchetti, S.; Greider, C.W. Human Telomerase RNA and Telomerase Activity in Immortal Cell Lines and Tumor Tissues. Cancer Res. 1996, 56.
- Castle, J.C.; Armour, C.D.; Löwer, M.; Haynor, D.; Biery, M.; Bouzek, H.; Chen, R.; Jackson, S.; Johnson, J.M.; Rohl, C.A.; et al. Digital Genome-Wide NcRNA Expression, Including SnoRNAs, across 11 Human Tissues Using PolyA-Neutral Amplification. PLoS One 2010, 5.
- Hartmann, N.; Reichwald, K.; Lechel, A.; Graf, M.; Kirschner, J.; Dorn, A.; Terzibasi, E.; Wellner, J.; Platzer, M.; Rudolph, K.L.; et al. Telomeres Shorten While Tert Expression Increases during Ageing of the Short-Lived Fish Nothobranchius Furzeri. Mech. Ageing Dev. 2009, 130, 290–296.
- Agarwal, S.; Loh, Y.H.; McLoughlin, E.M.; Huang, J.; Park, I.H.; Miller, J.D.; Huo, H.; Okuka, M.; Dos Reis, R.M.; Loewer, S.; et al. Telomere Elongation in Induced Pluripotent Stem Cells from Dyskeratosis Congenita Patients. Nature 2010, 464, 292–296.
- Weinrich, S.L.; Pruzan, R.; Ma, L.; Ouellette, M.; Tesmer, V.M.; Holt, S.E.; Bodnar, A.G.; Lichtsteiner, S.; Kim, N.W.; Trager, J.B.; et al. Reconstitution of Human Telomerase with the Template RNA Component HTR and the Catalytic Protein Subunit HTRT. Nat. Genet. 1997, 17, 498–502.
- Miller, M.C.; Liu, J.K.; Collins, K. Template Definition by Tetrahymena Telomerase Reverse Transcriptase. EMBO J. 2000, 19, 4412–4422.
- Moriarty, T.J.; Huard, S.; Dupuis, S.; Autexier, C. Functional Multimerization of Human Telomerase Requires an RNA Interaction Domain in the N Terminus of the Catalytic Subunit. Mol. Cell. Biol. 2002, 22, 1253–1265.
- Sealey, D.C.F.; Zheng, L.; Taboski, M.A.S.; Cruickshank, J.; Ikura, M.; Harrington, L.A. The N-Terminus of HTERT Contains a DNA-Binding Domain and Is Required for Telomerase Activity and Cellular Immortalization. Nucleic Acids Res. 2010, 38, 2019–2035.
- Lai, C.K.; Mitchell, J.R.; Collins, K. RNA Binding Domain of Telomerase Reverse Transcriptase. Mol. Cell. Biol. 2001, 21, 990–1000.
- Moriarty, T.J.; Marie-Egyptienne, D.T.; Autexier, C. Functional Organization of Repeat Addition Processivity and DNA Synthesis Determinants in the Human Telomerase Multimer. Mol. Cell. Biol. 2004, 24, 3720–3733.
- Mitchell, J.R.; Collins, K. Human Telomerase Activation Requires Two Independent Interactions between Telomerase RNA and Telomerase Reverse Transcriptase. Mol. Cell 2000, 6, 361–371.
- Friedman, K.L.; Cech, T.R. Essential Functions of Amino-Terminal Domains in the Yeast Telomerase Catalytic Subunit Revealed by Selection for Viable Mutants. Genes Dev. 1999, 13, 2863–2874.
- Willers, H.; McCarthy, E.E.; Wu, B.; Wunsch, H.; Tang, W.; Taghian, D.G.; Xia, F.; Powell, S.N. Dissociation of P53-Mediated Suppression of Homologous Recombination from G1/S Cell Cycle Checkpoint Control. Oncogene 2000, 19, 632–639.
- Beattie, T.L.; Zhou, W.; Robinson, M.O.; Harrington, L. Reconstitution of Human Telomerase Activity in Vitro. Curr. Biol. 1998, 8, 177–180.
- Nakamura, T.M.; Morin, G.B.; Chapman, K.B.; Weinrich, S.L.; Andrews, W.H.; Lingner, J.; Harley, C.B.; Cech, T.R. Telomerase Catalytic Subunit Homologs from Fission Yeast and Human. Science 1997, 277, 955–959.
- Bosoy, D.; Lue, N.F. Functional Analysis of Conserved Residues in the Putative “Finger” Domain of Telomerase Reverse Transcriptase. J. Biol. Chem. 2001, 276, 46305–46312.
- Xie, M.; Podlevsky, J.D.; Qi, X.; Bley, C.J.; Chen, J.J.L. A Novel Motif in Telomerase Reverse Transcriptase Regulates Telomere Repeat Addition Rate and Processivity. Nucleic Acids Res. 2010, 38, 1982–1996.
- Gillis, A.J.; Schuller, A.P.; Skordalakes, E. Structure of the Tribolium Castaneum Telomerase Catalytic Subunit TERT. Nature 2008, 455, 633–637.
- Seimiya, H.; Sawada, H.; Muramatsu, Y.; Shimizu, M.; Ohko, K.; Yamane, K.; Tsuruo, T. Involvement of 14-3-3 Proteins in Nuclear Localization of Telomerase. EMBO J. 2000, 19, 2652–2661.
- Haendeler, J.; Hoffmann, J.; Brandes, R.P.; Zeiher, A.M.; Dimmeler, S. Hydrogen Peroxide Triggers Nuclear Export of Telomerase Reverse Transcriptase via Src Kinase Family-Dependent Phosphorylation of Tyrosine 707. Mol. Cell. Biol. 2003, 23, 4598–4610.
- Ye, A.J.; Romero, D.P. Phylogenetic Relationships amongst Tetrahymenine Ciliates Inferred by a Comparison of Telomerase RNAs. Int. J. Syst. Evol. Microbiol. 2002, 52, 2297–2302.
- Chakrabarti, K.; Pearson, M.; Grate, L.; Sterne-Weiler, T.; Deans, J.; Donohue, J.P.; Ares, M. Structural RNAs of Known and Unknown Function Identified in Malaria Parasites by Comparative Genomics and RNA Analysis. RNA 2007, 13, 1923–1939.
- Song, J.; Logeswaran, D.; Castillo-González, C.; Li, Y.; Bose, S.; Aklilu, B.B.; Ma, Z.; Polkhovskiy, A.; Chen, J.J.L.; Shippen, D.E. The Conserved Structure of Plant Telomerase RNA Provides the Missing Link for an Evolutionary Pathway from Ciliates to Humans. Proc. Natl. Acad. Sci. USA 2019, 116, 24542–24550.
- Qi, X.; Li, Y.; Honda, S.; Hoffmann, S.; Marz, M.; Mosig, A.; Podlevsky, J.D.; Stadler, P.F.; Selker, E.U.; Chen, J.J.L. The Common Ancestral Core of Vertebrate and Fungal Telomerase RNAs. Nucleic Acids Res. 2013, 41, 450–462.
- Logeswaran, D.; Li, Y.; Podlevsky, J.D.; Chen, J.J.L. Monophyletic Origin and Divergent Evolution of Animal Telomerase RNA. Mol. Biol. Evol. 2021, 38, 215–228.
- Podlevsky, J.D.; Chen, J.J.L. Evolutionary Perspectives of Telomerase RNA Structure and Function. RNA Biol. 2016, 13, 720–732.
- Chen, J.L.; Blasco, M.A.; Greider, C.W. Secondary Structure of Vertebrate Telomerase RNA. Cell 2000, 100, 503–514.
- Theimer, C.A.; Feigon, J. Structure and Function of Telomerase RNA. Curr. Opin. Struct. Biol. 2006, 16, 307–318.
- Chen, J.L.; Greider, C.W. Template Boundary Definition in Mammalian Telomerase. Genes Dev. 2003, 17, 2747–2752.
- Hossain, S.; Singh, S.; Lue, N.F. Functional Analysis of the C-Terminal Extension of Telomerase Reverse Transcriptase. A Putative “Thumb” Domain. J. Biol. Chem. 2002, 277, 36174–36180.
- Förstemann, K.; Lingner, J. Telomerase Limits the Extent of Base Pairing between Template RNA and Telomeric DNA. EMBO Rep. 2005, 6, 361–366.
- Brown, Y.; Abraham, M.; Pearl, S.; Kabaha, M.M.; Elboher, E.; Tzfati, Y. A Critical Three-Way Junction Is Conserved in Budding Yeast and Vertebrate Telomerase RNAs. Nucleic Acids Res. 2007, 35, 6280–6289.
- Blackburn, E.H.; Collins, K. Telomerase: An RNP Enzyme Synthesizes DNA. Cold Spring Harb. Perspect. Biol. 2011, 3, 1–9.
- Huang, J.; Brown, A.F.; Wu, J.; Xue, J.; Bley, C.J.; Rand, D.P.; Wu, L.; Zhang, R.; Chen, J.J.L.; Lei, M. Structural Basis for Protein-RNA Recognition in Telomerase. Nat. Struct. Mol. Biol. 2014, 21, 507–512.
- Mitchell, J.R.; Cheng, J.; Collins, K. A Box H/ACA Small Nucleolar RNA-like Domain at the Human Telomerase RNA 3’ End. Mol. Cell. Biol. 1999, 19, 567–576.
- Vulliamy, T.J.; Marrone, A.; Knight, S.W.; Walne, A.; Mason, P.J.; Dokal, I. Mutations in Dyskeratosis Congenita: Their Impact on Telomere Length and the Diversity of Clinical Presentation. Blood 2006, 107, 2680–2685.
- Li, H. Unveiling Substrate RNA Binding to H/ACA RNPs: One Side Fits All. Curr. Opin. Struct. Biol. 2008, 18, 78–85.
- Reichow, S.L.; Hamma, T.; Ferré-D’Amaré, A.R.; Varani, G. The Structure and Function of Small Nucleolar Ribonucleoproteins. Nucleic Acids Res. 2007, 35, 1452–1464.
- Cristofari, G.; Adolf, E.; Reichenbach, P.; Sikora, K.; Terns, R.M.; Terns, M.P.; Lingner, J. Human Telomerase RNA Accumulation in Cajal Bodies Facilitates Telomerase Recruitment to Telomeres and Telomere Elongation. Mol. Cell 2007, 27, 882–889.
- Theimer, C.A.; Jády, B.E.; Chim, N.; Richard, P.; Breece, K.E.; Kiss, T.; Feigon, J. Structural and Functional Characterization of Human Telomerase RNA Processing and Cajal Body Localization Signals. Mol. Cell 2007, 27, 869–881.
- Kiss, T.; Fayet-Lebaron, E.; Jády, B.E. Box H/ACA Small Ribonucleoproteins. Mol. Cell 2010, 37, 597–606.
- Egan, E.D.; Collins, K. Specificity and Stoichiometry of Subunit Interactions in the Human Telomerase Holoenzyme Assembled in Vivo. Mol. Cell. Biol. 2010, 30, 2775–2786.
- Lafontaine, D.L.J.; Bousquet-Antonelli, C.; Henry, Y.; Caizergues-Ferrer, M.; Tollervey, D. The Box H + ACA SnoRNAs Carry Cbf5p, the Putative RRNA Pseudouridine Synthase. Genes Dev. 1998, 12, 527–537.
- Ganot, P.; Bortolin, M.L.; Kiss, T. Site-Specific Pseudouridine Formation in Preribosomal RNA Is Guided by Small Nucleolar RNAs. Cell 1997, 89, 799–809.
- Kiss, A.M.; Jády, B.E.; Darzacq, X.; Verheggen, C.; Bertrand, E.; Kiss, T. A Cajal Body-Specific Pseudouridylation Guide RNA Is Composed of Two Box H/ACA SnoRNA-like Domains. Nucleic Acids Res. 2002, 30, 4643–4649.
- Richard, P.; Darzacq, X.; Bertrand, E.; Jády, B.E.; Verheggen, C.; Kiss, T. A Common Sequence Motif Determines the Cajal Body-Specific Localization of Box H/ACA ScaRNAs. EMBO J. 2003, 22, 4283–4293.
- Wang, C.; Meier, U.T. Architecture and Assembly of Mammalian H/ACA Small Nucleolar and Telomerase Ribonucleoproteins. EMBO J. 2004, 23, 1857–1867.
- Grozdanov, P.N.; Roy, S.; Kittur, N.; Meier, U.T. SHQ1 Is Required Prior to NAF1 for Assembly of H/ACA Small Nucleolar and Telomerase RNPs. RNA 2009, 15, 1188–1197.
- Girard, J.P.; Lehtonen, H.; Caizergues-Ferrer, M.; Amalric, F.; Tollervey, D.; Lapeyre, B. GAR1 Is an Essential Small Nucleolar RNP Protein Required for Pre-RRNA Processing in Yeast. EMBO J. 1992, 11, 673.
- Maiorano, D.; Brimage, L.J.E.; Leroy, D.; Kearsey, S.E. Functional Conservation and Cell Cycle Localization of the Nhp2 Core Component of H + ACA SnoRNPs in Fission and Budding Yeasts. Exp. Cell Res. 1999, 252, 165–174.
- Venteicher, A.S.; Abreu, E.B.; Meng, Z.; McCann, K.E.; Terns, R.M.; Veenstra, T.D.; Terns, M.P.; Artandi, S.E. A Human Telomerase Holoenzyme Protein Required for Cajal Body Localization and Telomere Synthesis. Science 2009, 323, 644–648.
- Freund, A.; Zhong, F.L.; Venteicher, A.S.; Meng, Z.; Veenstra, T.D.; Frydman, J.; Artandi, S.E. Proteostatic Control of Telomerase Function through TRiC-Mediated Folding of TCAB1. Cell 2014, 159, 1389–1403.
- Flint, J.; Craddock, C.F.; Villegas, A.; Bentley, D.P.; Williams, H.J.; Galanello, R.; Cao, A.; Wood, W.G.; Ayyub, H.; Higgs, D.R. Healing of Broken Human Chromosomes by the Addition of Telomeric Repeats. Am. J. Hum. Genet. 1994, 55, 505.
- Nergadze, S.G.; Santagostino, M.A.; Salzano, A.; Mondello, C.; Giulotto, E. Contribution of Telomerase RNA Retrotranscription to DNA Double-Strand Break Repair during Mammalian Genome Evolution. Genome Biol. 2007, 8.
- Nandakumar, J.; Cech, T.R. Finding the End: Recruitment of Telomerase to Telomeres. Nat. Rev. Mol. Cell Biol. 2013, 14, 69–82.
- Furuya, T.; Morgan, R.; Berger, C.S.; Sandberg, A.A. Presence of Telomeric Sequences on Deleted Chromosomes and Their Absence on Double Minutes in Cell Line HL-60. Cancer Genet. Cytogenet. 1993, 70, 132–135.
- Meltzer, P.S.; Guan, X.Y.; Trent, J.M. Telomere Capture Stabilizes Chromosome Breakage. Nat. Genet. 1993 43 1993, 4, 252–255.
- McClintock, B. The Stability of Broken Ends of Chromosomes in Zea Mays. Genetics 1941, 26, 234.
- Q, F.; M, Y. New Telomere Formation Coupled with Site-Specific Chromosome Breakage in Tetrahymena Thermophila. Mol. Cell. Biol. 1996, 16, 1267–1274.
- Matsumoto, T.; Fukui, K.; Niwa, O.; Sugawara, N.; Szostak, J.W.; Yanagida, M. Identification of Healed Terminal DNA Fragments in Linear Minichromosomes of Schizosaccharomyces Pombe. Mol. Cell. Biol. 1987, 7, 4424–4430.
- Kramer, K.M.; Haber, J.E. New Telomeres in Yeast Are Initiated with a Highly Selected Subset of TG1-3 Repeats. Genes Dev. 1993, 7, 2345–2356.
- Wang, S.S.; Zakian, V.A. Telomere-Telomere Recombination Provides an Express Pathway for Telomere Acquisition. Nature 1990, 345, 456–458.
- Zhang, J.M.; Yadav, T.; Ouyang, J.; Lan, L.; Zou, L. Alternative Lengthening of Telomeres through Two Distinct Break-Induced Replication Pathways. Cell Rep. 2019, 26, 955–968.
- Hong, J.; Lee, J.H.; Chung, I.K. Telomerase Activates Transcription of Cyclin D1 Gene through an Interaction with NOL1. J. Cell Sci. 2016, 129, 1566–1579.
More
Information
Subjects:
Biochemistry & Molecular Biology
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.9K
Revisions:
2 times
(View History)
Update Date:
20 Dec 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No