Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Gulzar Badroo | -- | 2770 | 2022-12-12 16:04:50 | | | |
| 2 | Jessie Wu | Meta information modification | 2770 | 2022-12-13 07:26:02 | | | | |
| 3 | Jessie Wu | -3 word(s) | 2767 | 2022-12-13 07:27:51 | | | | |
| 4 | Jessie Wu | + 8 word(s) | 2775 | 2022-12-13 07:30:57 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Gazal, S.; Sharma, N.; Gazal, S.; Tikoo, M.; Shikha, D.; Badroo, G.A.; Rashid, M.; Lee, S. Strategies for Nipah and Hendra Viruses. Encyclopedia. Available online: https://encyclopedia.pub/entry/38609 (accessed on 27 July 2026).
Gazal S, Sharma N, Gazal S, Tikoo M, Shikha D, Badroo GA, et al. Strategies for Nipah and Hendra Viruses. Encyclopedia. Available at: https://encyclopedia.pub/entry/38609. Accessed July 27, 2026.
Gazal, Sabahat, Neelesh Sharma, Sundus Gazal, Mehak Tikoo, Deep Shikha, Gulzar Ahmed Badroo, Mohd Rashid, Sung-Jin Lee. "Strategies for Nipah and Hendra Viruses" Encyclopedia, https://encyclopedia.pub/entry/38609 (accessed July 27, 2026).
Gazal, S., Sharma, N., Gazal, S., Tikoo, M., Shikha, D., Badroo, G.A., Rashid, M., & Lee, S. (2022, December 12). Strategies for Nipah and Hendra Viruses. In Encyclopedia. https://encyclopedia.pub/entry/38609
Gazal, Sabahat, et al. "Strategies for Nipah and Hendra Viruses." Encyclopedia. Web. 12 December, 2022.
Copy Citation
Nipah and Hendra viruses are deadly zoonotic paramyxoviruses with a case fatality rate of upto 75%. The viruses belong to the genus henipavirus in the family Paramyxoviridae, a family of negative-sense single-stranded RNA viruses. The natural reservoirs of Nipah virus (NiV) and Hendra virus (HeV) are bats (flying foxes) in which the virus infection is asymptomatic. The intermediate hosts for NiV and HeV are swine and equine, respectively. In humans, NiV infections result in severe and often fatal respiratory and neurological manifestations. The Nipah virus was first identified in Malaysia and Singapore following an outbreak of encephalitis in pig farmers and subsequent outbreaks have been reported in Bangladesh and India almost every year.
NiV
Nipah virus
Hendra virus
1. Henipaviruses: A Cause of Grave Concern
Henipaviruses, particularly the Nipah virus, have many characteristics which make them a significant threat to human and animal health. Firstly, fruit bats (Pteropus genus), which are natural reservoirs of the virus, are widely distributed throughout Asia, leading to frequent spillover events. Second, they can be transmitted from bats to humans directly or via domestic animals. Third, they are capable of human-to-human transmission. Fourth, spillover has frequently occurred in highly populated regions. Fifth, they have a high mortality rate in humans. Lastly, there are no vaccines to prevent or antivirals to mitigate the disease [1]. Thus, considering all of the abovementioned factors, Nipah virus (NiV) poses a significant threat to human health. In humans, NiV infection has been associated with a range of clinical manifestations, including widespread vasculitis, encephalitis, and myocarditis, with approximate mortality rates of 40% to 70% [2]. Infected individuals can sometimes exhibit symptoms such as drowsiness and mental confusion, which can progress to coma. Residual neurological complications have been reported in some survivors [3]. The clinical presentation of NiV infections in humans, swine, and equine is shown in Figure 1.
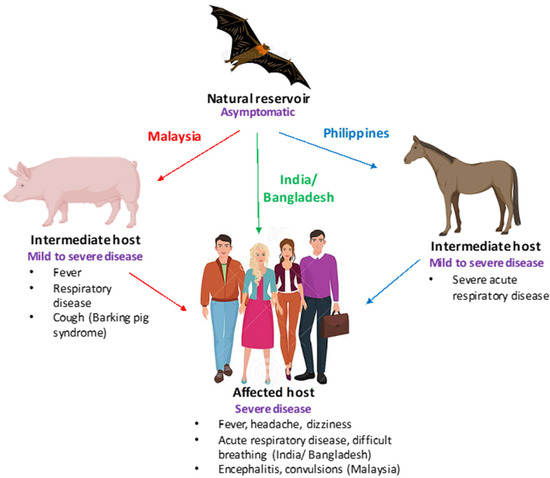
Figure 1. Clinical presentation of Nipah virus (NiV) infections. A schematic depicting the clinical presentation of NiV infection is shown. Bats (flying foxes) are the natural reservoirs of NiV and are asymptomatic. In swine (Malaysia) and equine (Philippines),NiV infection is characterized by severe respiratory disease, cough, and neurological signs, while in humans fever, encephalitis (Malaysia), and acute respiratory disease (Bangladesh) have been reported.
NiV was first identified in Malaysia and Singapore in 1998–1999 after an outbreak of respiratory and neurological disease in pigs, soon followed by cases of encephalitis in pig farmers. The virus isolated from the cerebrospinal fluid of patients that had died of the infection cross-reacted with Hendra virus (HeV) antibodies and was genetically closely related to HeV. The virus was named Nipah after the Malaysian village where one of the patients lived. The outbreak led to 105 human fatalities, with a total of 265 reported cases in Malaysia, and 1 human fatality with 11 cases reported in abattoir workers in Singapore. The disease had a huge economic impact, as 1.1 million swine had to be sacrificed to contain the infection [4]. In 2001, a different strain of NiV was identified as an etiological agent responsible for cases of encephalitis reported in Bangladesh, namely NiV-B. Since then, NiV infections in humans have been reported in Bangladesh almost annually. In fact, from 2001 to 2015, a total of 260 NiV cases have been confirmed in Bangladesh [5].
NiV outbreaks have also been reported in India, with the largest one being in Siliguri, West Bengal in 2001 (around 66 cases and 45 deaths) and a smaller outbreak with five cases and a 100% fatality rate reported in 2007 in Nadia district, West Bengal [6]. These outbreaks occurred across the border from the Nipah belt in Bangladesh; however, recently, in 2018, a NiV outbreak occurred in the Kozhikode and Malappuram districts of Kerala, a region in South India which is geographically disconnected from the areas affected by previous outbreaks [7]. In 2019, a single NiV case was identified in Kerala’s Ernakulum district. More recently, on 1 September 2021, NiV encephalitis was confirmed in a 12-year-old boy in Kozhikode, Kerala who succumbed to his illness on 5 September 2021 [3].
Cases of NiV encephalitis have also been reported in Philippines; in 2014, a total of 17 human cases were reported, out of which 11 exhibited symptoms of acute encephalitis while 6 cases exhibited influenza-like illness or meningitis.
In sum, a total of 650 human cases of NiV infection have occurred in Southeast Asia since 2001 with a combined case fatality rate of 60% [2].
Similar to NiV, the natural reservoirs of HeV are Pteropus bats, which result in fatal disease in both horses and humans. The virus can be transmitted to horses after exposure to excretions of HeV-infected bats, particularly urine. HeV can further be transmitted from horses to humans through excretions or the body fluids of horses infected with HeV. In humans, the symptoms of HeV infection can range from mild influenza-like symptoms to fatal respiratory disease or neurological manifestations. As with NiV, no vaccine or antivirals have been licensed for use against HeV in humans; however, a vaccine has been approved for use in horses (discussed in Vaccine section under subunit vaccines).
The first outbreak of the Hendra virus presented as a severe respiratory disease occurring in horses in 1994 in Brisbane (a suburb of Hendra), Australia, leading to the death of 14 horses and their trainer. Seven other horses and one person were also infected, but did not succumb to the infection. A novel paramyxovirus was identified as the etiological agent and was termed equine morbillivirus, which is now referred to as HeV [8][9][10]. However, the first known case of HeV occurred a few months prior when there were cases of HeV infection in horses; a person who assisted in the necropsy of the horses developed fatal encephalitis and died 13 months later [11]. Since the first identification of HeV in Australia, these viruses have emerged 62 times in Australia, resulting in 104 equine fatalities either due to fatal infection or euthanization [12]. In addition, a total of seven human cases and four deaths have been attributed to HeV with all human infections acquired via virus shed by horses [13].
Other than being fatal to humans and animals, henipavirus infection can also result in huge economic losses, which can be ascertained by the fact that 1.1 million swine had to be sacrificed to contain the 1998 NiV outbreak in Malaysia [4]. This resulted in a loss of US $97 million in compensation for the pigs that had to be culled to contain the outbreak. In addition, an indirect cost of US $229 million was incurred due to lost tax revenue to the government as well as losses in international trade. Furthermore, an additional US $136 million was lost for control programmes, viz. for biosecurity and animal slaughter [14]. The pork industry was severely affected by the Malaysian outbreak, which resulted in long-term effects post-outbreak as a result of altered pork consumption and pork export. The pork consumption and export dropped by around 80% during the Malaysian outbreak, while a dip of 30% was observed post-outbreak [15].
The importance of henipaviruses, especially NiV as a human pathogen, is further accentuated by the fact that NiV has crossed the species barrier on several occasions, resulting in infections in humans and animals with limited person-to-person transmission [16]. The transmission of NiV to humans occurs mainly in places where bats, pigs, and humans come in close proximity to one another. People rear pigs for economic benefits and in the vicinage of the farms, plant fruit-bearing trees for shade. Fruit bats belonging to the genus Pteropus are attracted to fruits and drop partially eaten saliva-laden fruits, which are consumed by domestic animals and thus result in spillover of infection to domestic animals/pigs from which transmission to humans can occur. This was particularly the case with the Malaysian outbreak. NiV outbreaks have also occurred by consumption of raw date palm sap (in Bangladesh), undercooked infected meat, and by handling infected animals [17]. Thus, NiV infection in humans can occur via intermediate hosts or directly from bats to humans and can also result in human-to-human transmission. Considering all these factors and due to the human-to human-transmission patterns for NiV infections coupled with high case fatality rates, NiV poses a significant threat to human health, particularly due to the lack of available vaccines and antivirals.
2. Recent Strategies for the Control of the Nipah and Hendra Viruses
2.1. Passive Immunization Using Monoclonal Antibodies
Using recombinant antibody technology, a human monoclonal antibody has been generated that is specific for henipavirus G protein (m102.4). The m102.4 binds to ephrin B2/B3 binding sites on the G protein and thus blocks viral infection. The antibody can neutralize HeV, NiV-M (Malaysian strain), and NiV-B (Bangladesh strain) and can provide protection in ferrets and non-human primates when inoculated with a lethal dose of NiV (NiV-M and NiV-B) at various time points after exposure within a short therapeutic window [18][19]. The immunization with m102.4 would be most suited for post-exposure treatment in case of any future outbreak situations. To date, m102.4 has been administered to 14 individuals exposed to HeV in Australia (13 individuals) and NiV in the United States (1 individual) and no adverse effects have been noted in any of the individuals. m102.4 antibody was used in a randomized and controlled phase 1 study in Australia [20]. The antibody was found to be safe with no immunogenicity and a have half-life ranging from approximately 16.5 to 17 days.
Another monoclonal antibody that has been generated for henipaviruses is 5B3 (h5B3.1). The antibody is cross-reactive to NiV and HeV F proteins and binds to F protein on a pre-fusion conformation epitope, thereby preventing membrane fusion [21]. The antibody provides protection to NiV and HeV in ferrets when administered one to several days post challenge. Taken together, these studies show that passive immunization can be used to provide therapeutic benefit, allowing the infected individuals an extended period to mount a protective immune response [22].
2.2. Recent Developments in Vaccine Production
Since the G (attachment protein) and F (fusion protein) proteins of paramyxoviruses are the viral proteins against which neutralizing antibodies are directed to, NiV and HeV G and F proteins have been the targets for vaccination strategies.
2.2.1. Subunit Vaccines
A soluble oligomeric form of the HeV G protein (HeV-sG) has been used as a subunit vaccine and has been found to protect animals including cats, ferrets, and African green monkeys against HeV and NiV challenge without any clinical signs of the disease, viral pathology, or even virus replication. The vaccinated animals developed high levels of neutralizing antibodies. The subunit vaccine has been approved in Australia for use in equines under the trade name of Equivac® [23]. However, the vaccine resulted in the generation of low levels of neutralizing antibodies in pigs following NiV challenge [24].
2.2.2. Vectored Vaccines
Various viral vectors have been used for the generation of NiV vaccine candidates including pox viruses, adeno-associated viruses, rhabdoviruses, and paramyxoviruses.
Poxviruses including the vaccinia virus that expresses NiV for G proteins have been produced and used in experiments to vaccinate a hamster model. These proteins, irrespective of being expressed alone or together, were able to provide complete protection against NiV challenge. Passive antibody transfer from vaccinated hamsters (exhibiting a high titer of neutralizing antibodies) to naïve hamsters was also observed [25]. A canarypox vaccine vector that expresses NiV F and G proteins either alone or in combination has been recently developed. This vaccine has been used in experiments to immunize pigs and was found to result in the production of neutralizing antibodies without virus shedding or the exhibition of histopathological lesions. The simultaneous expression of both of NiV surface glycoproteins, viz. F and G proteins, was found to produce the highest neutralizing antibody responses [26].
Adeno virus and Adeno-associated virus (AAV) have also been used as vectors for NiV vaccine production. Adeno virus belongs to the Adenoviridae family; however, Adeno-associated virus (AAV) belongs to the Parvoviridae family. AAV-expressing NiV (Malaysian strain) G protein has been produced and provided complete resistance to NiV-M challenge in hamsters with no symptoms of clinical illness [27]. Human adenovirus (HAV) vectors possess a problem with preexisting immunity; however, chimpanzee adenoviral (ChAd) vectors do not have such issues [28]. Previously, Oxford 1 (ChAdOx1), a genetically modified chimpanzee adenoviral vector, was investigated for use in the generation of a NiV/HeV vaccine [29]. The ChAdOx1-expressing G glycoprotein from NiV-B (ChAdOx1 NiV-B) has been used to immunize hamsters via the intramuscular route, either as a single shot or as part of a prime-boost strategy followed by virus challenge 42 days after the booster or the single vaccine with NiV-B. Only one dose of ChAdOx1 expressing NiV-B G protein was shown to be adequate to provide complete protection against NiV-B.
Rhabdoviruses, including VSV and rabies virus, have also been utilized in an attempt to develop a NiV vaccine. As vaccine candidates, VSV vector platforms have been exploited in a variety of ways. These include using replication-competent VSV that expresses NiV F, G, or N proteins [30]. The VSV vaccines expressing G from NiV-M and NiV-B strains were sufficient to protect both Syrian hamsters and ferrets in a single dose. Animals vaccinated with NiV F- or G-expressing vectors exhibited high neutralizing Ab titers, low viral RNA/antigen, and exhibited no pathological alterations [31][32]. Only partial protection was conferred in hamsters that had been vaccinated with recombinant VSV-expressing N protein from NiV, implying the possibility that both cellular and non-neutralizing antibody responses contribute to protection against disease caused by NiV [33].
Evaluations have also been conducted on rabies virus SAD B19 vaccine strain that expresses HeV or NiV G proteins [34]. In mice, this recombinant virus was utilized to express the wild-type or codon-optimized version of the HeV G gene [30]. The F or G protein-expressing recombinant rabies virus(Evelyn–Rokitnicki–Abelseth strain) was recently tested in mice and pigs [35]. This vector can also serve as an oral vaccine in dogs. This live-attenuated vector, while not suited for human use, is a potential veterinary vaccine for NiV because it is already authorized for use in some species and may be modified for use in emergency situations to guard against NiV infection in livestock, particularly swine.
Paramyxoviruses including recombinant Measles virus and recombinant Newcastle disease virus have also been used as vaccine vectors. The recombinant measles virus expressing NiV glycoprotein provided complete protection against NiV challenge in hamsters [36]. In addition, a paramyxovirus a recombinant Newcastle disease virus that expresses the NiV G or F proteins has also been described [37]. NDV vectors are better vaccine platforms, as they can be propagated in chicken eggs in high titers without the need to use cell culture.
Other viruses, such as the Venezuelan equine encephalitis virus encoding NiV F or G proteins, have been used as vaccine candidates and have shown highly potent neutralizing antibody responses in mice [38].
2.3. Virus-like Particles
Virus-like particles (VLPs) are nanostructures thatclosely resemble viruses but do not contain viral genome and are therefore non-infectious. The expression of selected viral proteins via expression plasmids results in the spontaneous assembly and release of VLPs. Since these particles possess many characteristics of authentic virus particles, such surface dimensions and structure, they are very efficient immunogens. Furthermore, these particles are very safe owing to the absence of viral genetic material. Vaccines based on the VLP platform that have been approved for use in humans against viruses include Gardasil against human papillomavirus and Sci-B-Vac and Bio-HepB against Hepatitis B virus. VLP-based vaccines are being developed for many non-enveloped and enveloped viruses, including NiV. For the production of NiV VLPs, three NiV proteins, namely, F, G, and M proteins, have been shown to be crucial. These three proteins have been expressed simultaneously in mammalian cells for NiV VLP production. The VLPs produced by the simultaneous expression of F, G, and M proteins have been found to be fusogenic and to induce syncytia formation. The VLPs have been found to be composed of all the three expressed proteins with intracellular processing similar to that observed for NiV virions. Moreover, the VLP morphology was similar to the NiV virions. Furthermore, these VLPs activated the innate immune system, as evidenced by PCR array analysis [39]. These VLPs have also been evaluated in a Balb/c mouse model and have been proven to generate neutralizing antibodies, as well as to provide protection to hamsters against NiV challenge following the administration of three doses of the vaccine and also with a single-shot [40].
2.4. Single-Dose Lipid Nanoparticle mRNA Vaccine
Because of their safety, efficacy, and ease of use, messenger RNA (mRNA)-based vaccines have become a popular vaccine technique lately. The limited efficiency of HeV-sG mRNA lipid nanoparticle has been reported. Though encouraging, there exists the need for further adjuvant addition, adjuvant route modification, and/or prime-boost vaccination strategies. Recently, the National Institute of Allergy and Infectious Diseases (USA) launched a phase 1 clinical trial to evaluate an experimental vaccine based on an mRNA platform. The vaccine, mRNA-1215 Nipah vaccine, was developed by Moderna (Cambridge, MA, USA) and involves the same technology as was used in many approved COVID-19 vaccines. The phase 1 trial is being carried out in Maryland at the NIH Clinical Centre, Bethesda [41].
References
- Epstein, J.H.; Anthony, S.J.; Islam, A.; Kilpatrick, A.M.; Khan, S.A.; Balkey, M.D.; Ross, N.; Smith, I.; Zambrana-Torrelio, C.; Tao, Y.; et al. Nipah virus dynamics in bats and implications for spillover to humans. Proc. Natl. Acad. Sci. USA 2020, 117, 29190–29201.
- Luby, S.P.; Hossain, M.J.; Gurley, E.S.; Ahmed, B.N.; Banu, S.; Khan, S.U.; Homaira, N.; Rota, P.A.; Rollin, P.E.; Comer, J.A.; et al. Recurrent zoonotic transmission of Nipah virus into humans, Bangladesh, 2001. Emerg. Infect. Dis. 2009, 15, 1229–1235.
- Thakur, V.; Thakur, P.; Ratho, R.K. Nipah Outbreak: Is it the beginning of another pandemic in the era of COVID-19 and Zika. Brain. Behav. Immun. 2022, 99, 25–26.
- Chua, K.B. Nipah virus outbreak in Malaysia. J. Clin. Virol. 2003, 26, 265–275.
- Satterfield, B.A.; Cross, R.W.; Fenton, K.A.; Borisevich, V.; Agans, K.N.; Deer, D.J.; Graber, J.; Basler, C.F.; Geisbert, T.W.; Mire, C.E. Nipah virus C and W proteins contribute to respiratory disease in ferrets. J. Virol. 2016, 90, 6326–6343.
- Arankalle, V.A.; Bandyopadhyay, B.T.; Ramdasi, A.Y.; Jadi, R.; Patil, D.R.; Rahman, M.; Majumdar, M.; Banerjee, P.S.; Hati, A.K.; Goswami, R.P.; et al. Genomic Characterization of Nipah Virus, West Bengal, India. Emerg. Infec. Dis. 2011, 17, 907–909.
- Aditi; Shariff, M. Nipah virus infection: A review. Epidemiol. Infect. 2019, 147, 95.
- Murray, K.; Rogers, R.; Selvey, L.; Selleck, P.; Hyatt, A.; Gould, A.; Gleeson, L.; Hooper, P.; Westbury, H. A novel morbillivirus pneumonia of horses and its transmission to humans. Emerg. Infect. Dis. 1995, 1, 31–33.
- Murray, K.; Selleck, P.; Hooper, P.; Hyatt, A.; Gould, A.; Gleeson, L.; Westbury, H.; Hiley, L.R.; Selvey, L.; Rodwell, B.; et al. A morbilli virus that caused fatal disease in horses and humans. Science 1995, 268, 94–97.
- Selvey, L.A.; Wells, R.M.; McCormack, J.G.; Ansford, A.J.; Murray, K.; Rogers, R.J.; Lavercombe, P.S.; Selleck, P.; Sherdian, J.W. Infection of humans and horses by a newly described morbillivirus. Med. J. Aust. 1995, 162, 642–645.
- O’Sullivan, J.D.; Allworth, A.M.; Paterson, D.L.; Snow, T.M.; Boots, R.; Geelson, L.J.; Gould, A.R.; Hyatt, A.D.; Bradfield, J. Fatal encephalitis due to novel paramyxovirus transmitted from horses. Lancet 1997, 349, 93–95.
- Queensland Government. Summary of Hendra Virus Incidents in Horses. 2019. Available online: https://www.business.qld.gov.au/industries/service-industries-professionals/service-industries/veterinary-surgeons/guidelines-hendra/incident-summary (accessed on 10 April 2022).
- Playford, E.G.; McCall, B.; Smith, G.; Slinko, V.; Allen, G.; Smith, I.; Moore, F.; Taylor, C.; Kung, H.H.; Field, H. Human Hendra virus encephalitis associated with equine outbreak, Australia. Emerg. Infect. Dis. 2010, 16, 219–223.
- Dimmock, N.J.; Easton, A.J.; Leppard, K.N. (Eds.) Introduction to Modern Virology; School of Life Sciences University of Warwick: Coventry, UK; John Wiley & Sons: Hoboken, NJ, USA, 2016.
- Hosono, H.; Kono, H.; Ito, S.; Shirai, J. Economic impact of Nipah virus infection outbreak in Malaysia, Obihiro University of Agriculture and Veterinary Medicine; National Institute of Animal Health. In Proceedings of the 11th International Symposium on Veterinary Epidemiology and Economics, Cairns, Australia, 6–11 August 2006.
- Gurley, E.S.; Hegde, S.T.; Hossain, K.; Sazzad, H.M.S.; Hossain, M.J.; Rahman, M.; Sharker, M.A.Y.; Salje, H.; Islam, M.S.; Epstein, J.H.; et al. Convergence of humans, bats, trees, and culture in Nipah virus transmission, Bangladesh. Emerg. Infect. Dis. 2017, 23, 1446–1453.
- Chanchal, D.K.; Alok, S.; Sabharwal, M.; Bijauliya, R.K.; Rashi, S. Nipah: Silently rising infection. Int. J. Pharm. Sci. Res. 2018, 9, 3128–3135.
- Bossart, K.N.; Zhu, Z.; Middleton, D.; Klippel, J.; Crameri, G.; Bingham, J.; McEachern, J.A.; Green, D.; Hancock, T.J.; Chan, Y.P.; et al. A neutralizing human monoclonal antibody protects against lethal disease in a new ferret model of acute Nipah virus infection. PLoS Pathog. 2008, 5, 1000642.
- Xu, K.; Rockx, B.; Xie, Y.; Debuysscher, B.L.; Fusco, D.L.; Zhu, Z.; Chan, Y.P.; Xu, Y.; Luu, T.; Cer, R.Z.; et al. Crystal structure of the Hendra virusattachment G glycoprotein bound to a potent cross-reactive neutralizing human monoclonal antibody. PLoS Pathog. 2013, 9, 1003684.
- Playford, E.G.; Munro, T.; Mahler, S.M.; Elliott, S.; Gerometta, M.; Hoger, K.L.; Jones, M.L.; Griffin, P.; Lynch, K.D.; Carroll, H.; et al. Safety, tolerability, pharmacokinetics, and immunogenicity of a human monoclonal antibody targeting the G glycoprotein of henipaviruses in healthy adults: A first-in-human, randomised, controlled, phase 1 study. Lancet Infect. Dis. 2020, 20, 445–454.
- Dang, H.V.; Chan, Y.P.; Park, Y.J.; Snijder, J.; DaSilva, S.C.; Vu, B.; Yan, L.; Feng, Y.R.; Rockx, B.; Geisbert, T.W.; et al. An antibody against the F glycoprotein inhibits Nipah and Hendra virus infections. Nat. Struct. Mol. Biol. 2019, 26, 980–987.
- Amaya, M.; Broder, C.C. Vaccines to Emerging Viruses: Nipah and Hendra. Ann. Rev. Virol. 2020, 7, 447–473.
- Broder, C.C.; Xu, K.; Nikolov, D.B.; Zhu, Z.; Dimitrov, D.S.; Middleton, D. A treatment for and vaccine against the deadly Hendra and Nipah viruses. Antivir. Res. 2013, 100, 8–13.
- Pickering, B.S.; Hardham, J.M.; Smith, G.; Weingartl, E.T.; Dominowski, P.J.; Foss, D.L.; Mwangi, D.; Broder, C.C.; Roth, J.A.; Weingartl, H.M. Protection against henipaviruses in swine requires both, cell-mediated and humoral immune response. Vaccine 2016, 34, 4777–4786.
- Guillaume, V.; Contamin, H.; Loth, P.; Georges-Courbot, M.C.; Lefeuvre, A.; Marianneau, P.; Wild, T.F. Nipah virus: Vaccination and passive protection studies in a hamster model. J. Virol. 2004, 78, 834–840.
- Weingartl, H.M.; Berhane, Y.; Caswell, J.L. Recombinantnipah virus vaccines protect pigs against challenge. J. Virol. 2006, 80, 7929–7938.
- Ploquin, A.; Szecsi, J.; Mathieu, C.; Guillaume, V.; Barateau, V.; Ong, K.C.; Wong, K.T.; Cosset, F.L.; Horvat, B.; Salvetti, A. Protection against henipavirus infection by use of recombinant adeno-associated virus–vector vaccines. J. Infect. Dis. 2013, 207, 469–478.
- Ewer, K.; Sebastian, S.; Spencer, A.J.; Gilbert, S.; Hill, A.V.S.; Lambe, T. Chimpanzee adenoviral vectors as vaccines for outbreak pathogens. Hum. Vaccines Immunother. 2017, 13, 3020–3032.
- Van Doremalen, N.; Lambe, T.; Sebastian, S.; Bushmaker, T.; Fischer, R.; Feldmann, F.; Haddock, E.; Letko, M.; Avanzato, V.A.; Rissanen, I.; et al. single-dose ChAdOx1- vectored vaccine provides complete protection against Nipah Bangladesh and Malaysia in Syrian golden hamsters. PLoS Negl. Trop. Dis. 2019, 13, 0007462.
- Mire, C.E.; Versteeg, K.M.; Cross, R.W. Single injection recombinant vesicular stomatitis virus vaccines protect ferrets against lethal Nipah virus disease. J. Virol. 2013, 10, 353.
- Lo, M.K.; Bird, B.H.; Chattopadhyay, A.; Drew, C.P.; Martin, B.E.; Coleman, J.D.; Rose, J.K.; Nichol, S.T.; Spiropoulou, C.F. Single-dose replication-defective VSV-based Nipah virus vaccines provide protection from lethal challenge in Syrian hamsters. Antiviral Res. 2014, 101, 26–29.
- Prescott, J.; DeBuysscher, B.L.; Feldmann, F.; Gardner, D.J.; Haddock, E.; Martellaro, C.; Scott, D.; Feldmann, H. Single-dose live-attenuated vesicular stomatitis virus-based vaccine protects African green monkeys from Nipah virus disease. Vaccine 2015, 4, 2823–2829.
- DeBuysscher, B.L.; Scott, D.; Marzi, A.; Prescott, J.; Feldmann, H. Single-dose live-attenuated Nipah virus vaccines confer complete protection by eliciting antibodies directed against surface glycoproteins. Vaccine 2014, 32, 2637–2644.
- Kurup, D.; Wirblich, C.; Feldmann, H.; Marzi, A.; Schnell, M.J. Rhabdovirus-based vaccine platforms against henipaviruses. J. Virol. 2015, 89, 144–154.
- Shuai, L.; Ge, J.; Wen, Z.; Wang, J.; Wang, X.; Bu, Z. Immune responses in mice and pigs after oral vaccination with rabies virus vectored Nipah disease vaccines. Vet. Microbiol. 2020, 241, 108549.
- Yoneda, M.; Georges-Courbot, M.C.; Ikeda, F.; Ishii, M.; Nagata, N.; Jacquot, F.; Raoul, H.; Sato, H.; Kai, C. Recombinant measles virus vaccine expressing the Nipah virus glycoprotein protects against lethal Nipah virus challenge. PLoS ONE 2013, 8, 58414.
- Kong, D.; Wen, Z.; Su, H.; Ge, J.; Chen, W.; Wang, X.; Wu, C.; Yang, C.; Chen, H.; Bu, Z. Newcastle disease virus-vectored Nipah encephalitis vaccines induce B and T cell responses in mice and long-lasting neutralizing antibodies in pigs. Virology 2012, 25, 327–335.
- Defang, G.N.; Khetawat, D.; Broder, C.C.; Quinnan, G.V., Jr. Induction of neutralizing antibodies to Hendra and Nipah glycoproteins using a Venezuelan equine encephalitis virus in vivo expression system. Vaccines 2010, 29, 212–220.
- Walpita, P.; Cong, Y.; Jahrling, P.B.; Rojas, O.; Postnikova, E.; Yu, S.; Johns, L.; Holbrook, M.R. A VLP-based vaccine provides complete protection against Nipah virus challenge following multiple-dose or single-dose vaccination schedules in a hamster model. NPJ Vaccines 2017, 2, 21.
- Walpita, P.; Barr, J.; Sherman, M.; Basler, C.F.; Wang, L. Vaccine potential of Nipah Virus Like Particles. PLoS ONE 2011, 6, 18437.
- National Institute of Allergy and Infectious Diseases. Available online: https://www.niaid.nih.gov/news-events/nih-launches-clinical-trial-mrna-nipah-virus-vaccine (accessed on 11 November 2022).
More
Information
Subjects:
Agriculture, Dairy & Animal Science
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
755
Revisions:
4 times
(View History)
Update Date:
13 Dec 2022
Table of Contents



Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No