 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Benoît Chénais | + 1695 word(s) | 1695 | 2020-12-08 09:38:19 | | | |
| 2 | Vivi Li | -27 word(s) | 1668 | 2020-12-22 09:52:26 | | |
Video Upload Options
Fucoxanthin is a well-known carotenoid of the xanthophyll family, mainly produced by marine organisms such as the macroalgae of the fucus genus or microalgae such as Phaeodactylum tricornutum. Fucoxanthin has antioxidant and anti-inflammatory properties but also several anticancer effects. Fucoxanthin induces cell growth arrest, apoptosis, and/or autophagy in several cancer cell lines as well as in animal models of cancer. Fucoxanthin treatment leads to the inhibition of metastasis-related migration, invasion, epithelial–mesenchymal transition, and angiogenesis. Fucoxanthin also affects the DNA repair pathways, which could be involved in the resistance phenotype of tumor cells. Moreover, combined treatments of fucoxanthin, or its metabolite fucoxanthinol, with usual anticancer treatments can support conventional therapeutic strategies by reducing drug resistance.
1. Introduction
Carotenoids are colored and natural pigments widely distributed in nature. They include more than 1100 molecules [1], divided into two classes: xanthophylls, which contain oxygen, and carotenes, which are pure hydrocarbons. In photosynthetic organisms such as plants and algae, carotenoids have two major roles: they absorb energy for photosynthesis and also protect chlorophyll from photodamage [2]. At a structural level, xanthophyll pigments are close to each other, they share a long carbon chain containing one or more oxygen atoms, which differentiates them from carotenes. These carotenoids pigments are well-known for their antioxidant and anti-inflammatory properties [3][4][5], but they also display some potential anticancer effects [6].
First isolated in 1914 by Willstätter and Page, fucoxanthin (Figure 1) is an orange-colored xanthophyll pigment derived from brown algae and microalgae [7][8][9]. It is found in high content in taxons such as Phaeophyceae, Haptophyta, Bacillariophyceae, and Chrysophyceae, and to a lesser extent in Rhodophyta, Raphidophyceae, and Dinophyta [10][6]. Fucoxanthin is one of the most abundant carotenoid pigments, which contributes to more than 10% of the estimated total carotenoid production in nature, particularly in the marine environment [11].
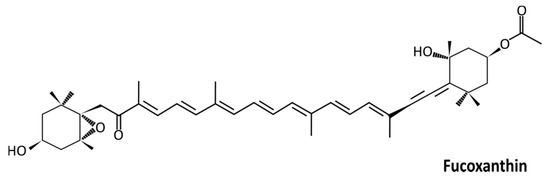
Figure 1. Structure of fucoxanthin (3′-acetoxy-5,6-epoxy-3,5′-dihydroxy-6′,7′-didehyro-5,6,7,8,5′,6′-hexahydro-β,β-carotene-8-one; C42H58O6; 658.91 g/mol).
Popular sources of fucoxanthin include macroalgae such as Laminaria japonica, Eisenia bicyclis, and the well-known brown seaweed Wakame (Undaria pinnatifida), as well as diatoms microalgae such as Phaeodactylum tricornutum [12]. Fucoxanthin exhibit several biological activities that are beneficial to human health including antioxidant, anti-inflammatory, anti-obesity, anti-diabetic, anti-angiogenic, and anticancer properties [6][13][14][15][16][17][18][19].
2. Antiproliferative Effects of Fucoxanthin
Antiproliferative effect of fucoxanthin has been reported for numerous cancer cell lines and the inhibition of cell proliferation by fucoxanthin is due to cell growth arrest at G0/G1 or G1 phase of the cell cycle [6]. The molecular mechanism of the G0/G1 phase arrest mainly involves the downregulation of cyclin D1 and/or D2 and cyclin-dependent kinase-4 (CDK4). However, cyclin E and CDK2 or CDK6 could also be involved in some cell types. Furthermore, studies showing tumor growth arrest in the presence of fucoxanthin with several types of cancer has confirmed the antiproliferative effect of fucoxanthin in vivo [20][21]. Fucoxanthin also induces cancer cell death through the induction of apoptosis and/or autophagy in various cellular cancer cell lines, including HeLa, SGC-7901 and human cervical cancer [22][23][24]. In addition, it appears that fucoxanthin-induced autophagy occurs prior to apoptosis and may be a promoter of apoptosis [23]. Molecular mechanisms that sustain the cell death induction by fucoxanthin include, but are not limited to, inhibition of Akt/mTOR signaling pathway [22], downregulation of NF-κB pathway, increase of Bax and decrease of Bcl2 and/or BclXL through the MAP kinase pathway (reviewed in [6]). The effects of fucoxanthin on cell death of cancer cell lines in vitro have been confirmed by in vivo studies in mice bearing xenografted sarcoma 180, where fucoxanthin, at 50 and 100 mg/kg, significantly inhibited the growth of sarcoma and induced apoptosis as demonstrated by the decrease of Bcl-2 expression and a clear increase of cleaved caspase-3 [25].
3. Anti-Metastatic Effect of Fucoxanthin
Fucoxanthin was found to possess strong anticancer and anti-metastatic activities that work irrespective of the p53 status of cancer cells and cause a decrease in hallmark proteins associated with the metastatic spread of cancer cells at doses that were relatively safe to the normal cells [26]. Fucoxanthin inhibited the expression and secretion of MMP-9, which plays a critical role in tumor invasion and migration, and suppressed invasion of highly metastatic B16-F10 melanoma cells [27] as well as the human glioblastoma cell line U87 [28]. This effect, in glioblastoma cell line U87, is dependent on the p38 MAPK signalling [28]. Furthermore, fucoxanthin decreased the expression of the cell surface glycoprotein CD44 and C-X-C motif chemokine receptor-4 (CXCR4), which are involved in migration, invasion and adhesion of cancer cells to endothelial cells. Indeed, the adhesion of B16-F10 melanoma cells to the endothelial cells was significantly inhibited by fucoxanthin [27]. Moreover, the development of osteosarcoma in mice inoculated with osteosarcoma cells was inhibited by a treatment with fucoxanthin [29] and in vivo metastasis was reduced by fucoxanthin as shown by a significant reduction of tumor nodules in an experimental lung metastasis in vivo assay [27] and glioblastoma xenografts [28].
4. Anti-Angiogenic Effect of Fucoxanthin
Angiogenesis could be defined as the process of remodeling the primitive network of blood vessels and its growth into a complex network that is regulated by the balance between pro- and anti-angiogenic molecules. During this process, vascular endothelial cells secrete proteases and then migrate through the extracellular matrix, proliferate, and differentiate into new blood vessels [30]. Pathological angiogenesis is involved in many diseases, including rheumatoid arthritis, atherosclerosis, diabetic retinopathy, and cancer [31]. Angiogenesis is mandatory for tumor progression because newly formed blood vessels are needed to supply oxygen and nutrients, which are essential to the growing tumor, and to remove waste products. Also, the metastasis process depends on angiogenesis, as tumor cells migrate from the primary tumor and grow in distant target organs. Few studies highlighted the anti-angiogenic potential of fucoxanthin [15][32][33]. First, fucoxanthin was shown to inhibit human umbilical vein endothelial cells’ (HUVEC) proliferation and tube formation, but without a significant effect on HUVEC chemotaxis [32]. Fucoxanthin (10–20 µM) also suppressed the development of blood vessel-like structures from CD31-positive cells, and then may suppress the differentiation of endothelial progenitor cells into endothelial cells involving new blood vessel formation. The molecular mechanism of this anti-angiogenic effect of fucoxanthin involves the downregulation of the mRNA level of FGF-2 and its receptor (FGFR-1) as well as their trans-activation factor, EGR-1, as shown in HUVEC treated by fucoxanthin [15]. Moreover, fucoxanthin downregulates the FGF-2-mediated phosphorylation of signaling proteins such as ERK1/2 and Akt, which leads to the repression of endothelial cells’ migration as well as their differentiation into tube-like structures on Matrigel® [15]. Fucoxanthin also inhibited the formation of tube-like structures from human lymphatic endothelial cells as well as it suppressed the malignant phenotype of human breast cancer MDA-MB-231 cells and decreased tumor-induced lymphangiogenesis when used in combination with a conditional medium culture system [33]. In vivo, using a MDA-MB-231 nude mouse model of breast cancer, high doses of fucoxanthin (100–500 µM) decreased micro-lymphatic vascular density, suggesting that fucoxanthin inhibits tumor-induced lymphangiogenesis in vitro and in vivo. At the cellular level, the mechanism of action of fucoxanthin involves decreased levels of VEGF-C, VEGF receptor-3, NF-κB, p-Akt, and p-PI3K in HLEC [33].
5. Anti-Inflammatory Effects of Fucoxanthin
Pro-inflammatory mediators including interleukins, tumor necrosis factor α (TNFα), prostaglandin E2, and nitric oxide contribute to the development of a variety of inflammatory diseases. Natural products such as carotenoids have been used in the prevention of oxidative stress due to their antioxidant activities [34]. Despite the fact that the oxidative or anti-oxidative properties of fucoxanthin are discussed and seem to depend on the cellular context, this compound has been studied in an anti-inflammatory context in vivo and in vitro [35][36].
Fucoxanthin inhibited cell viability of adult T-cell leukemia, which is a fatal malignancy of T lymphocytes caused by human T-cell leukemia virus type 1 (HTLV-1) [37]. Interestingly, uninfected cell lines and normal peripheral blood mononuclear cells were resistant to fucoxanthin [37]. The combination of fucoxanthin and rosmarinic acid (a bioactive compound from Lamiaceae plants) improved anti-inflammatory effects from each [16]. Indeed, the inflammatory response is modulated through downregulation of inflammasome components such as “NOD-like receptor family, pyrin domain containing 3” (NLRP3), “apoptosis-associated speck-like protein” (ASC), caspase-1, and interleukins. These results suggest that fucoxanthin, in combination with rosmarinic acid, exerts anti-inflammatory effects by downregulating NRLP3-inflammasome and increasing the NFE2L2/Nrf2 signaling pathway in UVB-exposed HaCaT keratinocytes [16].
6. Fucoxanthin Effect on DNA damage and DNA repair
The antioxidant properties of fucoxantin can limit ROS-induced DNA damage, such as UVB-induced DNA-damage in human fibroblast [38] or in rat glioma cells [39]. In addition, a protective effect of fucoxanthin toward bleomycin-induced DNA damage has been observed in MCF-7 breast cancer cells [40]. By contrast, fucoxanthin can also lead to DNA damages in cancer cells [41][42], and this can be used for anticancer therapy by increasing the cytotoxic effect of common drugs such as 5-fluoro-uracile in colon cancer cells [42]. This induction of DNA damage is mediated by the production of reactive oxygen species and the subsequent activation of ATM/ATR/p53 pathways and the phosphorylation of Ser139 of histone H2AX [43]. Fucoxanthin decreases the mRNA expression of the DNA repair gene ERCC1, which is induced by cisplatin and this leads to the improvement of chemotherapeutic efficacy of cisplatin [44].
7. Conclusions
Cellular and animal studies have shown that fucoxanthin has anticancer effects. However, investigation of this role in humans is lacking. Clinical trials are required to assess the effect of fucoxanthin in close connection with the study of the mechanisms involved in the antitumoral action of fucoxanthin. Moreover, anticancer effects of fucoxanthin are regulated by several mechanisms leading to cell cycle arrest, induction of cell death and DNA damages, inhibition of metastasis-related migration, invasion and epithelial–mesenchymal transition, anti-angiogenic, and anti-inflammatory effects. Furthermore, combined treatments of fucoxanthin or fucoxanthinol with usual anticancer treatments can support conventional therapeutic strategies by reducing drug resistance. Indeed, as an anticancer molecule, fucoxanthinol appears to be a more effective bioactive compound than fucoxanthin. Therefore, the potential use of fucoxanthinol and other fucoxanthin derivatives (such as apo-9′-fucoxanthinone and apo-13-fucoxanthinone) as co-adjuvant agents in the treatment of cancer should be further investigated. However, further studies are necessary for precise and complete inhibitory effects of fucoxanthin and its derivatives, but regarding these hallmarks, fucoxanthin appears to be a promising compound for cancer therapy.
References
- Junko Yabuzaki; Carotenoids Database: structures, chemical fingerprints and distribution among organisms. Database 2017, 2017, ?, 10.1093/database/bax004.
- Gregory A. Armstrong; John E. Hearst; Genetics and molecular biology of carotenoid pigment biosynthesis. The FASEB Journal 1996, 10, 228-237, 10.1096/fasebj.10.2.8641556.
- Jane H. Tinkler; Fritz Böhm; W. Schalch; T. George Truscott; Dietary carotenoids protect human cells from damage. Journal of Photochemistry and Photobiology B: Biology 1994, 26, 283-285, 10.1016/1011-1344(94)07049-0.
- Christopher S. Foote; Robert W. Denny; Chemistry of singlet oxygen. VII. Quenching by .beta.-carotene. Journal of the American Chemical Society 1968, 90, 6233-6235, 10.1021/ja01024a061.
- Kevin Jørgensen; Leif H. Skibsted; Carotenoid scavenging of radicals. European Food Research and Technology 1993, 196, 423-429, 10.1007/bf01190806.
- Sarah Méresse; Mostefa Fodil; Fabrice Fleury; Benoît Chénais; Fucoxanthin, a Marine-Derived Carotenoid from Brown Seaweeds and Microalgae: A Promising Bioactive Compound for Cancer Therapy. International Journal of Molecular Sciences 2020, 21, 9273, 10.3390/ijms21239273.
- Richard Willstätter; Harold J. Page; Untersuchungen über Chlorophyll. XXIV. Über die Pigmente der Braunalgen. European Journal of Organic Chemistry 1914, 404, 237-271, 10.1002/jlac.19144040302.
- R. Bonnett; A. K. Mallams; B. C. L. Weedon; A. McCormick; J. L. Tee; Fucoxanthin and related pigments. Chemical Communications (London) 1966, ?, 515-516, 10.1039/c19660000515.
- R. Bonnett; A. K. Mallams; A. A. Spark; J. L. Tee; B. C. L. Weedon; A. McCormick; Carotenoids and related compounds. Part XX. Structure and reactions of fucoxanthin. J. Chem. Soc. C 1969, ?, 429, 10.1039/j39690000429.
- Shinichi Takaichi; Carotenoids in Algae: Distributions, Biosyntheses and Functions. Marine Drugs 2011, 9, 1101-1118, 10.3390/md9061101.
- Liaaen-Jensen, S.. Marine Natural Products; Elsevier: Amsterdam, 1978; pp. 1-73.
- Sang Min Kim; Yu-Jin Jung; Oh-Nam Kwon; Kwang Hyun Cha; Byung-Hun Um; Donghwa Chung; Cheol-Ho Pan; A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Applied Biochemistry and Biotechnology 2012, 166, 1843-1855, 10.1007/s12010-012-9602-2.
- Hayato Maeda; Masashi Hosokawa; Tokutake Sashima; Katsura Funayama; Kazuo Miyashita; Fucoxanthin from edible seaweed, Undaria pinnatifida, shows antiobesity effect through UCP1 expression in white adipose tissues. Biochemical and Biophysical Research Communications 2005, 332, 392-397, 10.1016/j.bbrc.2005.05.002.
- Hayato Maeda; Masashi Hosokawa; Tokutake Sashima; Kazuo Miyashita; Dietary Combination of Fucoxanthin and Fish Oil Attenuates the Weight Gain of White Adipose Tissue and Decreases Blood Glucose in Obese/Diabetic KK-AyMice. Journal of Agricultural and Food Chemistry 2007, 55, 7701-7706, 10.1021/jf071569n.
- Ponesakki Ganesan; Kiminori Matsubara; Tatsuya Sugawara; Takashi Hirata; Marine algal carotenoids inhibit angiogenesis by down-regulating FGF-2-mediated intracellular signals in vascular endothelial cells. Molecular and Cellular Biochemistry 2013, 380, 1-9, 10.1007/s11010-013-1651-5.
- Azahara Rodríguez-Luna; Javier Ávila-Román; Helena Oliveira; Virginia Motilva; Elena Talero; Fucoxanthin and Rosmarinic Acid Combination Has Anti-Inflammatory Effects through Regulation of NLRP3 Inflammasome in UVB-Exposed HaCaT Keratinocytes.. Marine Drugs 2019, 17, 451, 10.3390/md17080451.
- Pai-An Hwang; Nam Nhut Phan; Wen-Jung Lu; Bui Thi Ngoc Hieu; Yen-Chang Lin; Low-molecular-weight fucoidan and high-stability fucoxanthin from brown seaweed exert prebiotics and anti-inflammatory activities in Caco-2 cells. Food & Nutrition Research 2016, 60, 32033, 10.3402/fnr.v60.32033.
- Claire Mayer; Martine Côme; Lionel Ulmann; Graziella Chini Zittelli; Cecilia Faraloni; Hassan Nazih; Khadija Ouguerram; Benoît Chénais; Virginie Mimouni; Preventive Effects of the Marine Microalga Phaeodactylum tricornutum, Used as a Food Supplement, on Risk Factors Associated with Metabolic Syndrome in Wistar Rats. Nutrients 2019, 11, 1069, 10.3390/nu11051069.
- Claire Mayer; Martine Côme; V. Blanckaert; Graziella Chini Zittelli; Cecilia Faraloni; Hassan Nazih; Khadija Ouguerram; Virginie Mimouni; Benoît Chénais; Effect of Carotenoids from Phaeodactylum tricornutum on Palmitate-Treated HepG2 Cells. Molecules 2020, 25, 2845, 10.3390/molecules25122845.
- Kil-Nam Kim; Ginnae Ahn; Soo-Jin Heo; Sung-Myung Kang; Min-Cheol Kang; Hye-Mi Yang; Daekyung Kim; Seong Woon Roh; Se-Kwon Kim; Byong-Tae Jeon; et al.Pyo-Jam ParkWon-Kyo JungYou-Jin Jeon Inhibition of tumor growth in vitro and in vivo by fucoxanthin against melanoma B16F10 cells. Environmental Toxicology and Pharmacology 2013, 35, 39-46, 10.1016/j.etap.2012.10.002.
- Chenghan Mei; Shunchang Zhou; Lin Zhu; Jiaxiong Ming; Fan-Dian Zeng; Rong Xu; Antitumor Effects of Laminaria Extract Fucoxanthin on Lung Cancer. Marine Drugs 2017, 15, 39, 10.3390/md15020039.
- Li-Li Hou; Chao Gao; Liang Chen; Guo-Qiang Hu; Song-Qiang Xie; Essential role of autophagy in fucoxanthin-induced cytotoxicity to human epithelial cervical cancer HeLa cells. Acta Pharmacologica Sinica 2013, 34, 1403-1410, 10.1038/aps.2013.90.
- Yue Zhu; Jing Cheng; Zhenli Min; Tingzi Yin; Rong Zhang; Wei Zhang; Ling Hu; Zhiwen Cui; Chengzhi Gao; Shiqiang Xu; et al.Chunxiang ZhangXia-Min Hu Effects of fucoxanthin on autophagy and apoptosis in SGC-7901cells and the mechanism. Journal of Cellular Biochemistry 2018, 119, 7274-7284, 10.1002/jcb.27022.
- Ye Jin; Shuang Qiu; Na Shao; Jianhua Zheng; Fucoxanthin and Tumor Necrosis Factor-Related Apoptosis-Inducing Ligand (TRAIL) Synergistically Promotes Apoptosis of Human Cervical Cancer Cells by Targeting PI3K/Akt/NF-κB Signaling Pathway. Medical Science Monitor 2018, 24, 11-18, 10.12659/msm.905360.
- Jun Wang; Shihui Chen; Shiqiang Xu; Xing Yu; Dongqing Ma; Xiamin Hu; Xiao-Lu Cao; In Vivo Induction of Apoptosis by Fucoxanthin, a Marine Carotenoid, Associated with Down-Regulating STAT3/EGFR Signaling in Sarcoma 180 (S180) Xenografts-Bearing Mice. Marine Drugs 2012, 10, 2055-2068, 10.3390/md10092055.
- Sukant Garg; Sajal Afzal; Ahmed Elwakeel; Damini Sharma; Navaneethan Radhakrishnan; Jaspreet Kaur Dhanjal; Durai Sundar; Sunil C. Kaul; Renu Wadhwa; Marine Carotenoid Fucoxanthin Possesses Anti-Metastasis Activity: Molecular Evidence. Marine Drugs 2019, 17, 338, 10.3390/md17060338.
- Tae-Wook Chung; Hee-Jung Choi; Ji-Yeon Lee; Han-Sol Jeong; Cheorl-Ho Kim; Myungsoo Joo; Jun-Yong Choi; Chang-Woo Han; So-Yeon Kim; Jae-Sue Choi; et al.Ki-Tae Ha Marine algal fucoxanthin inhibits the metastatic potential of cancer cells. Biochemical and Biophysical Research Communications 2013, 439, 580-585, 10.1016/j.bbrc.2013.09.019.
- Yugang Liu; Jian Zheng; Yan Zhang; Zhaotao Wang; Yang Yang; MiaoChun Bai; Yiwu Dai; Fucoxanthin Activates Apoptosis via Inhibition of PI3K/Akt/mTOR Pathway and Suppresses Invasion and Migration by Restriction of p38-MMP-2/9 Pathway in Human Glioblastoma Cells. Neurochemical Research 2016, 41, 2728-2751, 10.1007/s11064-016-1989-7.
- Takayoshi Rokkaku; Ryuichiro Kimura; Chie Ishikawa; Takeshi Yasumoto; Masachika Senba; Fuminori Kanaya; Naoki Mori; Anticancer effects of marine carotenoids, fucoxanthin and its deacetylated product, fucoxanthinol, on osteosarcoma. International Journal of Oncology 2013, 43, 1176-1186, 10.3892/ijo.2013.2019.
- Folkman, J.; Shing, Y.; Angiogenesis. J. Biol. Chem. 1992, 267, 10931–10934.
- J Folkman; M Klagsbrun; Angiogenic factors. Science 1987, 235, 442-447, 10.1126/science.2432664.
- Tatsuya Sugawara; Kiminori Matsubara; Reiko Akagi; Masaharu Mori; Takashi Hirata; Antiangiogenic Activity of Brown Algae Fucoxanthin and Its Deacetylated Product, Fucoxanthinol. Journal of Agricultural and Food Chemistry 2006, 54, 9805-9810, 10.1021/jf062204q.
- Jia Wang; Yanhong Ma; Jingshi Yang; Lu Jin; Zixiang Gao; Lingyun Xue; Lin Hou; Linlin Sui; Jing Liu; Xiangyang Zou; et al. Fucoxanthin inhibits tumour‐related lymphangiogenesis and growth of breast cancer. Journal of Cellular and Molecular Medicine 2019, 23, 2219-2229, 10.1111/jcmm.14151.
- Palanisamy Arulselvan; Masoumeh Tangestani Fard; Woan Sean Tan; Sivapragasam Gothai; Sharida Fakurazi; Norhaizan Mohd Esa; S. Suresh Kumar; Role of Antioxidants and Natural Products in Inflammation. Oxidative Medicine and Cellular Longevity 2016, 2016, 1-15, 10.1155/2016/5276130.
- Kariyappa Vijay; Poorigali Raghavendra-Rao Sowmya; Bangalore Prabhashankar Arathi; Shivaprasad Shilpa; Hulikere Jagdish Shwetha; Marisiddaiah Raju; Vallikanan Baskaran; Rangaswamy Lakshminarayana; Low-dose doxorubicin with carotenoids selectively alters redox status and upregulates oxidative stress-mediated apoptosis in breast cancer cells. Food and Chemical Toxicology 2018, 118, 675-690, 10.1016/j.fct.2018.06.027.
- Juhyun Shin; Min-Ho Song; Jae-Wook Oh; Young-Soo Keum; Ramesh Kumar Saini; Pro-oxidant Actions of Carotenoids in Triggering Apoptosis of Cancer Cells: A Review of Emerging Evidence. Antioxidants 2020, 9, 532, 10.3390/antiox9060532.
- Chie Ishikawa; Senji Tafuku; Takashi Kadekaru; Shigeki Sawada; Mariko Tomita; Taeko Okudaira; Tetsuro Nakazato; Takayoshi Toda; Jun-Nosuke Uchihara; Naoya Taira; et al.Kazuiku OhshiroTakeshi YasumotoTakao OhtaNaoki Mori Antiadult T-cell leukemia effects of brown algae fucoxanthin and its deacetylated product, fucoxanthinol. International Journal of Cancer 2008, 123, 2702-2712, 10.1002/ijc.23860.
- Soo-Jin Heo; You-Jin Jeon; Protective effect of fucoxanthin isolated from Sargassum siliquastrum on UV-B induced cell damage. Journal of Photochemistry and Photobiology B: Biology 2009, 95, 101-107, 10.1016/j.jphotobiol.2008.11.011.
- Sajal Afzal; Sukant Garg; Yoshiyuki Ishida; Keiji Terao; Sunil C. Kaul; Renu Wadhwa; Rat Glioma Cell-Based Functional Characterization of Anti-Stress and Protein Deaggregation Activities in the Marine Carotenoids, Astaxanthin and Fucoxanthin.. Marine Drugs 2019, 17, 189, 10.3390/md17030189.
- Seif-Eldin N. Ayyad; Salim A. Basaif; Adel F. Badria; Saleh T. Ezmirly; Walied M. Alarif; Farid A. Badria; Antioxidant, cytotoxic, antitumor, and protective DNA damage metabolites from the red sea brown alga Sargassum sp. Pharmacognosy Research 2011, 3, 160-5, 10.4103/0974-8490.85000.
- Izumi Konishi; Masashi Hosokawa; Tokutake Sashima; Hidetaka Kobayashi; Kazuo Miyashita; Halocynthiaxanthin and fucoxanthinol isolated from Halocynthia roretzi induce apoptosis in human leukemia, breast and colon cancer cells. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology 2006, 142, 53-59, 10.1016/j.cbpc.2005.10.005.
- Eduarda Lopes-Costa; Mariana Abreu; Daniela Gargiulo; Eduardo Rocha; Alice A. Ramos; Anticancer effects of seaweed compounds fucoxanthin and phloroglucinol, alone and in combination with 5-fluorouracil in colon cells. Journal of Toxicology and Environmental Health, Part A 2017, 80, 776-787, 10.1080/15287394.2017.1357297.
- Hua-Lian Wu; Xiao-Yan Fu; Wen-Qiang Cao; Wen-Zhou Xiang; Ya-Jun Hou; Jin-Kui Ma; Ying Wang; Cun-Dong Fan; Induction of Apoptosis in Human Glioma Cells by Fucoxanthin via Triggering of ROS-Mediated Oxidative Damage and Regulation of MAPKs and PI3K–AKT Pathways. Journal of Agricultural and Food Chemistry 2019, 67, 2212-2219, 10.1021/acs.jafc.8b07126.
- Cheng-Ling Liu; Yun-Ping Lim; Miao-Lin Hu; Fucoxanthin Enhances Cisplatin-Induced Cytotoxicity via NFκB-Mediated Pathway and Downregulates DNA Repair Gene Expression in Human Hepatoma HepG2 Cells. Marine Drugs 2013, 11, 50-66, 10.3390/md11010050.



