 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Camila Xu | -- | 1434 | 2022-11-25 01:30:35 |
Video Upload Options
Amyloid precursor protein (APP) is an integral membrane protein expressed in many tissues and concentrated in the synapses of neurons. Its primary function is not known, though it has been implicated as a regulator of synapse formation, neural plasticity, antimicrobial activity , and iron export. APP is best known as the precursor molecule whose proteolysis generates amyloid beta (Aβ), a polypeptide containing 37 to 49 amino acid residues, whose amyloid fibrillar form is the primary component of amyloid plaques found in the brains of Alzheimer's disease patients.
1. Genetics
APP is an ancient and highly conserved protein.[1] In humans, the gene for APP is located on chromosome 21 and contains 18 exons spanning 290 kilobases.[2][3] Several alternative splicing isoforms of APP have been observed in humans, ranging in length from 639 to 770 amino acids, with certain isoforms preferentially expressed in neurons; changes in the neuronal ratio of these isoforms have been associated with Alzheimer's disease.[4] Homologous proteins have been identified in other organisms such as Drosophila (fruit flies), C. elegans (roundworms)[5], and all mammals.[6] The amyloid beta region of the protein, located in the membrane-spanning domain, is not well conserved across species and has no obvious connection with APP's native-state biological functions.[6]
Mutations in critical regions of amyloid precursor protein, including the region that generates amyloid beta (Aβ), cause familial susceptibility to Alzheimer's disease.[7][8][9] For example, several mutations outside the Aβ region associated with familial Alzheimer's have been found to dramatically increase production of Aβ.[10]
A mutation (A673T) in the APP gene protects against Alzheimer’s disease. This substitution is adjacent to the beta secretase cleavage site and results in a 40% reduction in the formation of amyloid beta in vitro.[11]
2. Structure
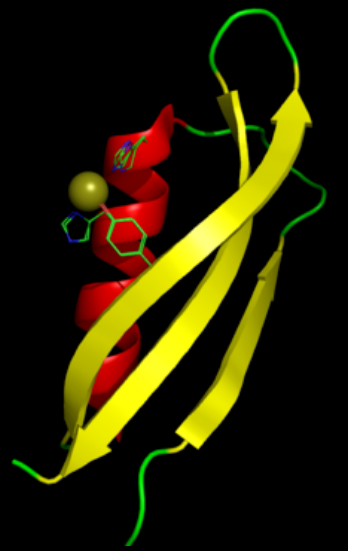
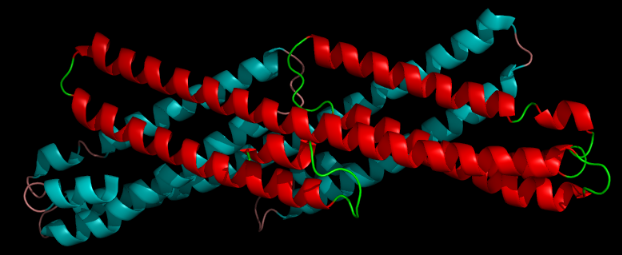
A number of distinct, largely independently-folding structural domains have been identified in the APP sequence. The extracellular region, much larger than the intracellular region, is divided into the E1 and E2 domains, linked by an acidic domain (AcD); E1 contains two subdomains including a growth factor-like domain (GFLD) and a copper-binding domain (CuBD) interacting tightly together.[13] A serine protease inhibitor domain, absent from the isoform differentially expressed in the brain, is found between acidic region and E2 domain.[14] The complete crystal structure of APP has not yet been solved; however, individual domains have been successfully crystallized, the growth factor-like domain,[15] the copper-binding domain,[16] the complete E1 domain[13] and the E2 domain.[12]
3. Post-Translational Processing
APP undergoes extensive post-translational modification including glycosylation, phosphorylation, sialylation, and tyrosine sulfation, as well as many types of proteolytic processing to generate peptide fragments.[17] It is commonly cleaved by proteases in the secretase family; alpha secretase and beta secretase both remove nearly the entire extracellular domain to release membrane-anchored carboxy-terminal fragments that may be associated with apoptosis.[6] Cleavage by gamma secretase within the membrane-spanning domain after beta-secretase cleavage generates the amyloid-beta fragment; gamma secretase is a large multi-subunit complex whose components have not yet been fully characterized, but include presenilin, whose gene has been identified as a major genetic risk factor for Alzheimer's.[18]
The amyloidogenic processing of APP has been linked to its presence in lipid rafts. When APP molecules occupy a lipid raft region of membrane, they are more accessible to and differentially cleaved by beta secretase, whereas APP molecules outside a raft are differentially cleaved by the non-amyloidogenic alpha secretase.[19] Gamma secretase activity has also been associated with lipid rafts.[20] The role of cholesterol in lipid raft maintenance has been cited as a likely explanation for observations that high cholesterol and apolipoprotein E genotype are major risk factors for Alzheimer's disease.[21]
4. Biological Function
Although the native biological role of APP is of obvious interest to Alzheimer's research, thorough understanding has remained elusive.
4.1. Synaptic Formation and Repair
The most-substantiated role for APP is in synaptic formation and repair;[22] its expression is upregulated during neuronal differentiation and after neural injury. Roles in cell signaling, long-term potentiation, and cell adhesion have been proposed and supported by as-yet limited research.[6] In particular, similarities in post-translational processing have invited comparisons to the signaling role of the surface receptor protein Notch.[23]
APP knockout mice are viable and have relatively minor phenotypic effects including impaired long-term potentiation and memory loss without general neuron loss.[24] On the other hand, transgenic mice with upregulated APP expression have also been reported to show impaired long-term potentiation.[25]
The logical inference is that because Aβ accumulates excessively in Alzheimer's disease its precursor, APP, would be elevated as well. However, neuronal cell bodies contain less APP as a function of their proximity to amyloid plaques.[26] The data indicate that this deficit in APP results from a decline in production rather than an increase in catalysis. Loss of a neuron's APP may affect physiological deficits that contribute to dementia.
4.2. Somatic Recombination
In neurons of the human brain, somatic recombination occurs frequently in the gene that encodes APP.[27] Neurons from individuals with sporadic Alzheimer's disease show greater APP gene diversity due to somatic recombination than neurons from healthy individuals.[27]
4.3. Anterograde Neuronal Transport
Molecules synthesized in the cell bodies of neurons must be conveyed outward to the distal synapses. This is accomplished via fast anterograde transport. It has been found that APP can mediate interaction between cargo and kinesin and thus facilitate this transport. Specifically, a short peptide 15-amino-acid sequence from the cytoplasmic carboxy-terminus is necessary for interaction with the motor protein.[28]
Additionally, it has been shown that the interaction between APP and kinesin is specific to the peptide sequence of APP.[29] In a recent experiment involving transport of peptide-conjugated colored beads, controls were conjugated to a single amino acid, glycine, such that they display the same terminal carboxylic acid group as APP without the intervening 15-amino-acid sequence mentioned above. The control beads were not motile, which demonstrated that the terminal COOH moiety of peptides is not sufficient to mediate transport.
4.4. Iron Export
A different perspective on Alzheimer's is revealed by a mouse study that has found that APP possesses ferroxidase activity similar to ceruloplasmin, facilitating iron export through interaction with ferroportin; it seems that this activity is blocked by zinc trapped by accumulated Aβ in Alzheimer's.[30] It has been shown that a single nucleotide polymorphism in the 5'UTR of APP mRNA can disrupt its translation.[31]
The hypothesis that APP has ferroxidase activity in its E2 domain and facilitates export of Fe(II) is possibly incorrect since the proposed ferroxidase site of APP located in the E2 domain does not have ferroxidase activity.[32][33]
As APP does not possess ferroxidase activity within its E2 domain, the mechanism of APP-modulated iron efflux from ferroportin has come under scrutiny. One model suggests that APP acts to stabilize the iron efflux protein ferroportin in the plasma membrane of cells thereby increasing the total number of ferroportin molecules at the membrane. These iron-transporters can then be activated by known mammalian ferroxidases (i.e. ceruloplasmin or hephaestin).[34]
4.5. Hormonal Regulation
The amyloid-β precursor protein (AβPP), and all associated secretases, are expressed early in development and play a key role in the endocrinology of reproduction – with the differential processing of AβPP by secretases regulating human embryonic stem cell (hESC) proliferation as well as their differentiation into neural precursor cells (NPC). The pregnancy hormone human chorionic gonadotropin (hCG) increases AβPP expression[35] and hESC proliferation while progesterone directs AβPP processing towards the non-amyloidogenic pathway, which promotes hESC differentiation into NPC.[36][37][38]
AβPP and its cleavage products do not promote the proliferation and differentiation of post-mitotic neurons; rather, the overexpression of either wild-type or mutant AβPP in post-mitotic neurons induces apoptotic death following their re-entry into the cell cycle.[39] It is postulated that the loss of sex steroids (including progesterone) but the elevation in luteinizing hormone, the adult equivalent of hCG, post-menopause and during andropause drives amyloid-β production[40] and re-entry of post-mitotic neurons into the cell cycle.
5. Interactions
Amyloid precursor protein has been shown to interact with:
- APBA1,[41][42]
- APBA2,[41][43][44]
- APBA3,[41][45]
- APBB1,[42][46][47][48][49]
- APPBP1,[50]
- APPBP2,[51]
- BCAP31,[52]
- BLMH[53]
- CLSTN1,[43][54]
- CAV1,[55]
- COL25A1,[56]
- FBLN1,[57]
- GSN,[58]
- HSD17B10,[59] and
- SHC1.[60]
APP interacts with reelin, a protein implicated in a number of brain disorders, including Alzheimer's disease.[61]
References
- "Origins of amyloid-β". BMC Genomics 14 (1): 290. April 2013. doi:10.1186/1471-2164-14-290. PMID 23627794. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3660159
- "Genomic organization of the human amyloid beta-protein precursor gene". Gene 87 (2): 257–63. Mar 1990. doi:10.1016/0378-1119(90)90310-N. PMID 2110105. https://dx.doi.org/10.1016%2F0378-1119%2890%2990310-N
- "Introduction and expression of the 400 kilobase amyloid precursor protein gene in transgenic mice [corrected]". Nature Genetics 5 (1): 22–30. Sep 1993. doi:10.1038/ng0993-22. PMID 8220418. https://dx.doi.org/10.1038%2Fng0993-22
- "Expression of APP pathway mRNAs and proteins in Alzheimer's disease". Brain Research 1161: 116–23. Aug 2007. doi:10.1016/j.brainres.2007.05.050. PMID 17586478. https://dx.doi.org/10.1016%2Fj.brainres.2007.05.050
- Ewald, Collin Y.; Li, Chris (2012-04-01). "Caenorhabditis elegans as a model organism to study APP function" (in en). Experimental Brain Research 217 (3–4): 397–411. doi:10.1007/s00221-011-2905-7. ISSN 0014-4819. PMID 22038715. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3746071
- "The amyloid precursor protein: beyond amyloid". Molecular Neurodegeneration 1 (1): 5. 2006. doi:10.1186/1750-1326-1-5. PMID 16930452. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1538601
- "Segregation of a missense mutation in the amyloid precursor protein gene with familial Alzheimer's disease". Nature 349 (6311): 704–6. Feb 1991. doi:10.1038/349704a0. PMID 1671712. Bibcode: 1991Natur.349..704G. https://dx.doi.org/10.1038%2F349704a0
- "A mutation in the amyloid precursor protein associated with hereditary Alzheimer's disease". Science 254 (5028): 97–9. Oct 1991. doi:10.1126/science.1925564. PMID 1925564. Bibcode: 1991Sci...254...97M. https://dx.doi.org/10.1126%2Fscience.1925564
- "Early-onset Alzheimer's disease caused by mutations at codon 717 of the beta-amyloid precursor protein gene". Nature 353 (6347): 844–6. Oct 1991. doi:10.1038/353844a0. PMID 1944558. Bibcode: 1991Natur.353..844C. https://dx.doi.org/10.1038%2F353844a0
- "Mutation of the beta-amyloid precursor protein in familial Alzheimer's disease increases beta-protein production". Nature 360 (6405): 672–4. Dec 1992. doi:10.1038/360672a0. PMID 1465129. https://dx.doi.org/10.1038%2F360672a0
- "A mutation in APP protects against Alzheimer's disease and age-related cognitive decline". Nature 488 (7409): 96–9. Aug 2012. doi:10.1038/nature11283. PMID 22801501. Bibcode: 2012Natur.488...96J. https://dx.doi.org/10.1038%2Fnature11283
- PDB: 1RW6; "The X-ray structure of an antiparallel dimer of the human amyloid precursor protein E2 domain". Molecular Cell 15 (3): 343–53. Aug 2004. doi:10.1016/j.molcel.2004.06.037. PMID 15304215. https://www.rcsb.org/structure/1RW6
- "Structure and biochemical analysis of the heparin-induced E1 dimer of the amyloid precursor protein". Proceedings of the National Academy of Sciences of the United States of America 107 (12): 5381–6. Mar 2010. doi:10.1073/pnas.0911326107. PMID 20212142. Bibcode: 2010PNAS..107.5381D. ; see also PDB ID 3KTM http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2851805
- "Identification and transport of full-length amyloid precursor proteins in rat peripheral nervous system". The Journal of Neuroscience 13 (7): 3136–42. Jul 1993. doi:10.1523/JNEUROSCI.13-07-03136.1993. PMID 8331390. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6576678
- "Crystal structure of the N-terminal, growth factor-like domain of Alzheimer amyloid precursor protein". Nature Structural Biology 6 (4): 327–31. Apr 1999. doi:10.1038/7562. PMID 10201399. ; see also PDB ID 1MWP https://dx.doi.org/10.1038%2F7562
- "Structural studies of the Alzheimer's amyloid precursor protein copper-binding domain reveal how it binds copper ions". Journal of Molecular Biology 367 (1): 148–61. Mar 2007. doi:10.1016/j.jmb.2006.12.041. PMID 17239395. ; See also 2007 PDB IDs 2FJZ, 2FK2, 2FKL. https://dx.doi.org/10.1016%2Fj.jmb.2006.12.041
- "Proteolytic processing and cell biological functions of the amyloid precursor protein". Journal of Cell Science 113 (11): 1857–70. Jun 2000. PMID 10806097. http://www.ncbi.nlm.nih.gov/pubmed/10806097
- "TMP21 is a presenilin complex component that modulates gamma-secretase but not epsilon-secretase activity". Nature 440 (7088): 1208–12. Apr 2006. doi:10.1038/nature04667. PMID 16641999. https://dx.doi.org/10.1038%2Fnature04667
- "Amyloidogenic processing of the Alzheimer beta-amyloid precursor protein depends on lipid rafts". The Journal of Cell Biology 160 (1): 113–23. Jan 2003. doi:10.1083/jcb.200207113. PMID 12515826. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2172747
- "Association of gamma-secretase with lipid rafts in post-Golgi and endosome membranes". The Journal of Biological Chemistry 279 (43): 44945–54. Oct 2004. doi:10.1074/jbc.M407986200. PMID 15322084. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1201506
- "Compartmentalization of beta-secretase (Asp2) into low-buoyant density, noncaveolar lipid rafts". Current Biology 11 (16): 1288–93. Aug 2001. doi:10.1016/S0960-9822(01)00394-3. PMID 11525745. https://dx.doi.org/10.1016%2FS0960-9822%2801%2900394-3
- "Synapse formation and function is modulated by the amyloid precursor protein". The Journal of Neuroscience 26 (27): 7212–21. Jul 2006. doi:10.1523/JNEUROSCI.1450-06.2006. PMID 16822978. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6673945
- "Notch and Presenilin: regulated intramembrane proteolysis links development and degeneration". Annual Review of Neuroscience 26 (1): 565–97. 2003. doi:10.1146/annurev.neuro.26.041002.131334. PMID 12730322. https://dx.doi.org/10.1146%2Fannurev.neuro.26.041002.131334
- "No hippocampal neuron or synaptic bouton loss in learning-impaired aged beta-amyloid precursor protein-null mice". Neuroscience 90 (4): 1207–16. 1999. doi:10.1016/S0306-4522(98)00645-9. PMID 10338291. https://dx.doi.org/10.1016%2FS0306-4522%2898%2900645-9
- "Inverse correlation between amyloid precursor protein and synaptic plasticity in transgenic mice". NeuroReport 18 (10): 1083–7. 2007. doi:10.1097/WNR.0b013e3281e72b18. PMID 17558301. https://dx.doi.org/10.1097%2FWNR.0b013e3281e72b18
- "Relationships between expression of apolipoprotein E and beta-amyloid precursor protein are altered in proximity to Alzheimer beta-amyloid plaques: potential explanations from cell culture studies". Journal of Neuropathology and Experimental Neurology 67 (8): 773–83. Aug 2008. doi:10.1097/NEN.0b013e318180ec47. PMID 18648325. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3334532
- "Somatic APP gene recombination in Alzheimer's disease and normal neurons". Nature 563 (7733): 639–645. November 2018. doi:10.1038/s41586-018-0718-6. PMID 30464338. Bibcode: 2018Natur.563..639L. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6391999
- "A peptide zipcode sufficient for anterograde transport within amyloid precursor protein". Proceedings of the National Academy of Sciences of the United States of America 103 (44): 16532–7. Oct 2006. doi:10.1073/pnas.0607527103. PMID 17062754. Bibcode: 2006PNAS..10316532S. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1621108
- "Quantitative measurements and modeling of cargo-motor interactions during fast transport in the living axon". Physical Biology 9 (5): 055005. Oct 2012. doi:10.1088/1478-3975/9/5/055005. PMID 23011729. Bibcode: 2012PhBio...9e5005S. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3625656
- "Iron-export ferroxidase activity of β-amyloid precursor protein is inhibited by zinc in Alzheimer's disease". Cell 142 (6): 857–67. Sep 2010. doi:10.1016/j.cell.2010.08.014. PMID 20817278. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2943017
- "Iron and the translation of the amyloid precursor protein (APP) and ferritin mRNAs: riboregulation against neural oxidative damage in Alzheimer's disease". Biochemical Society Transactions 36 (Pt 6): 1282–7. Dec 2008. doi:10.1042/BST0361282. PMID 19021541. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2746665
- "A synthetic peptide with the putative iron binding motif of amyloid precursor protein (APP) does not catalytically oxidize iron". PLOS ONE 7 (8): e40287. 2012. doi:10.1371/journal.pone.0040287. PMID 22916096. Bibcode: 2012PLoSO...740287E. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3419245
- "The amyloid precursor protein (APP) does not have a ferroxidase site in its E2 domain". PLOS ONE 8 (8): e72177. 2013. doi:10.1371/journal.pone.0072177. PMID 23977245. Bibcode: 2013PLoSO...872177H. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3747053
- "sAPP modulates iron efflux from brain microvascular endothelial cells by stabilizing the ferrous iron exporter ferroportin". EMBO Reports 15 (7): 809–15. Jul 2014. doi:10.15252/embr.201338064. PMID 24867889. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=4196985
- "Amyloid-beta precursor protein expression and modulation in human embryonic stem cells: a novel role for human chorionic gonadotropin". Biochemical and Biophysical Research Communications 364 (3): 522–7. Dec 2007. doi:10.1016/j.bbrc.2007.10.021. PMID 17959150. https://dx.doi.org/10.1016%2Fj.bbrc.2007.10.021
- "Differential processing of amyloid-beta precursor protein directs human embryonic stem cell proliferation and differentiation into neuronal precursor cells". The Journal of Biological Chemistry 284 (35): 23806–17. Aug 2009. doi:10.1074/jbc.M109.026328. PMID 19542221. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2749153
- "Opioid and progesterone signaling is obligatory for early human embryogenesis". Stem Cells and Development 18 (5): 737–40. Jun 2009. doi:10.1089/scd.2008.0190. PMID 18803462. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2891507
- "The pregnancy hormones human chorionic gonadotropin and progesterone induce human embryonic stem cell proliferation and differentiation into neuroectodermal rosettes". Stem Cell Research & Therapy 1 (4): 28. 2010. doi:10.1186/scrt28. PMID 20836886. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2983441
- "DNA synthesis and neuronal apoptosis caused by familial Alzheimer disease mutants of the amyloid precursor protein are mediated by the p21 activated kinase PAK3". The Journal of Neuroscience 23 (17): 6914–27. Jul 2003. doi:10.1523/JNEUROSCI.23-17-06914.2003. PMID 12890786. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6740729
- "Luteinizing hormone, a reproductive regulator that modulates the processing of amyloid-beta precursor protein and amyloid-beta deposition". The Journal of Biological Chemistry 279 (19): 20539–45. May 2004. doi:10.1074/jbc.M311993200. PMID 14871891. https://dx.doi.org/10.1074%2Fjbc.M311993200
- "Regulation of APP-dependent transcription complexes by Mint/X11s: differential functions of Mint isoforms". The Journal of Neuroscience 22 (17): 7340–51. Sep 2002. doi:10.1523/JNEUROSCI.22-17-07340.2002. PMID 12196555. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=6757996
- "The phosphotyrosine interaction domains of X11 and FE65 bind to distinct sites on the YENPTY motif of amyloid precursor protein". Molecular and Cellular Biology 16 (11): 6229–41. Nov 1996. doi:10.1128/mcb.16.11.6229. PMID 8887653. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=231626
- "Novel cadherin-related membrane proteins, Alcadeins, enhance the X11-like protein-mediated stabilization of amyloid beta-protein precursor metabolism". The Journal of Biological Chemistry 278 (49): 49448–58. Dec 2003. doi:10.1074/jbc.M306024200. PMID 12972431. https://dx.doi.org/10.1074%2Fjbc.M306024200
- "Interaction of a neuron-specific protein containing PDZ domains with Alzheimer's amyloid precursor protein". The Journal of Biological Chemistry 274 (4): 2243–54. Jan 1999. doi:10.1074/jbc.274.4.2243. PMID 9890987. https://dx.doi.org/10.1074%2Fjbc.274.4.2243
- "X11L2, a new member of the X11 protein family, interacts with Alzheimer's beta-amyloid precursor protein". Biochemical and Biophysical Research Communications 255 (3): 663–7. Feb 1999. doi:10.1006/bbrc.1999.0265. PMID 10049767. https://dx.doi.org/10.1006%2Fbbrc.1999.0265
- "Interaction of the phosphotyrosine interaction/phosphotyrosine binding-related domains of Fe65 with wild-type and mutant Alzheimer's beta-amyloid precursor proteins". The Journal of Biological Chemistry 272 (10): 6399–405. Mar 1997. doi:10.1074/jbc.272.10.6399. PMID 9045663. https://dx.doi.org/10.1074%2Fjbc.272.10.6399
- "Association of a novel human FE65-like protein with the cytoplasmic domain of the beta-amyloid precursor protein". Proceedings of the National Academy of Sciences of the United States of America 93 (20): 10832–7. Oct 1996. doi:10.1073/pnas.93.20.10832. PMID 8855266. Bibcode: 1996PNAS...9310832G. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=38241
- "Molecular cloning of human Fe65L2 and its interaction with the Alzheimer's beta-amyloid precursor protein". Neuroscience Letters 261 (3): 143–6. Feb 1999. doi:10.1016/S0304-3940(98)00995-1. PMID 10081969. https://dx.doi.org/10.1016%2FS0304-3940%2898%2900995-1
- "Interaction of cytosolic adaptor proteins with neuronal apolipoprotein E receptors and the amyloid precursor protein". The Journal of Biological Chemistry 273 (50): 33556–60. Dec 1998. doi:10.1074/jbc.273.50.33556. PMID 9837937. https://dx.doi.org/10.1074%2Fjbc.273.50.33556
- "APP-BP1, a novel protein that binds to the carboxyl-terminal region of the amyloid precursor protein". The Journal of Biological Chemistry 271 (19): 11339–46. May 1996. doi:10.1074/jbc.271.19.11339. PMID 8626687. https://dx.doi.org/10.1074%2Fjbc.271.19.11339
- "PAT1, a microtubule-interacting protein, recognizes the basolateral sorting signal of amyloid precursor protein". Proceedings of the National Academy of Sciences of the United States of America 95 (25): 14745–50. Dec 1998. doi:10.1073/pnas.95.25.14745. PMID 9843960. Bibcode: 1998PNAS...9514745Z. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=24520
- "Uncleaved BAP31 in association with A4 protein at the endoplasmic reticulum is an inhibitor of Fas-initiated release of cytochrome c from mitochondria". The Journal of Biological Chemistry 278 (16): 14461–8. Apr 2003. doi:10.1074/jbc.M209684200. PMID 12529377. https://dx.doi.org/10.1074%2Fjbc.M209684200
- "Human bleomycin hydrolase regulates the secretion of amyloid precursor protein". FASEB Journal 14 (12): 1837–47. Sep 2000. doi:10.1096/fj.99-0938com. PMID 10973933. https://dx.doi.org/10.1096%2Ffj.99-0938com
- "Coordinated metabolism of Alcadein and amyloid beta-protein precursor regulates FE65-dependent gene transactivation". The Journal of Biological Chemistry 279 (23): 24343–54. Jun 2004. doi:10.1074/jbc.M401925200. PMID 15037614. https://dx.doi.org/10.1074%2Fjbc.M401925200
- "Caveolae, plasma membrane microdomains for alpha-secretase-mediated processing of the amyloid precursor protein". The Journal of Biological Chemistry 273 (17): 10485–95. Apr 1998. doi:10.1074/jbc.273.17.10485. PMID 9553108. https://dx.doi.org/10.1074%2Fjbc.273.17.10485
- "CLAC: a novel Alzheimer amyloid plaque component derived from a transmembrane precursor, CLAC-P/collagen type XXV". The EMBO Journal 21 (7): 1524–34. Apr 2002. doi:10.1093/emboj/21.7.1524. PMID 11927537. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=125364
- "Fibulin-1 binds the amino-terminal head of beta-amyloid precursor protein and modulates its physiological function". Journal of Neurochemistry 76 (5): 1411–20. Mar 2001. doi:10.1046/j.1471-4159.2001.00144.x. PMID 11238726. https://dx.doi.org/10.1046%2Fj.1471-4159.2001.00144.x
- "Binding of gelsolin, a secretory protein, to amyloid beta-protein". Biochemical and Biophysical Research Communications 258 (2): 241–6. May 1999. doi:10.1006/bbrc.1999.0623. PMID 10329371. https://dx.doi.org/10.1006%2Fbbrc.1999.0623
- "An intracellular protein that binds amyloid-beta peptide and mediates neurotoxicity in Alzheimer's disease". Nature 389 (6652): 689–95. Oct 1997. doi:10.1038/39522. PMID 9338779. Bibcode: 1997Natur.389..689D. https://dx.doi.org/10.1038%2F39522
- "Tyrosine phosphorylation of the beta-amyloid precursor protein cytoplasmic tail promotes interaction with Shc". The Journal of Biological Chemistry 277 (19): 16798–804. May 2002. doi:10.1074/jbc.M110286200. PMID 11877420. https://dx.doi.org/10.1074%2Fjbc.M110286200
- "Interaction of reelin with amyloid precursor protein promotes neurite outgrowth". The Journal of Neuroscience 29 (23): 7459–73. Jun 2009. doi:10.1523/JNEUROSCI.4872-08.2009. PMID 19515914. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2759694

