 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Olga Senko | -- | 3612 | 2022-11-24 10:49:02 | | | |
| 2 | Camila Xu | Meta information modification | 3612 | 2022-11-25 03:50:55 | | |
Video Upload Options
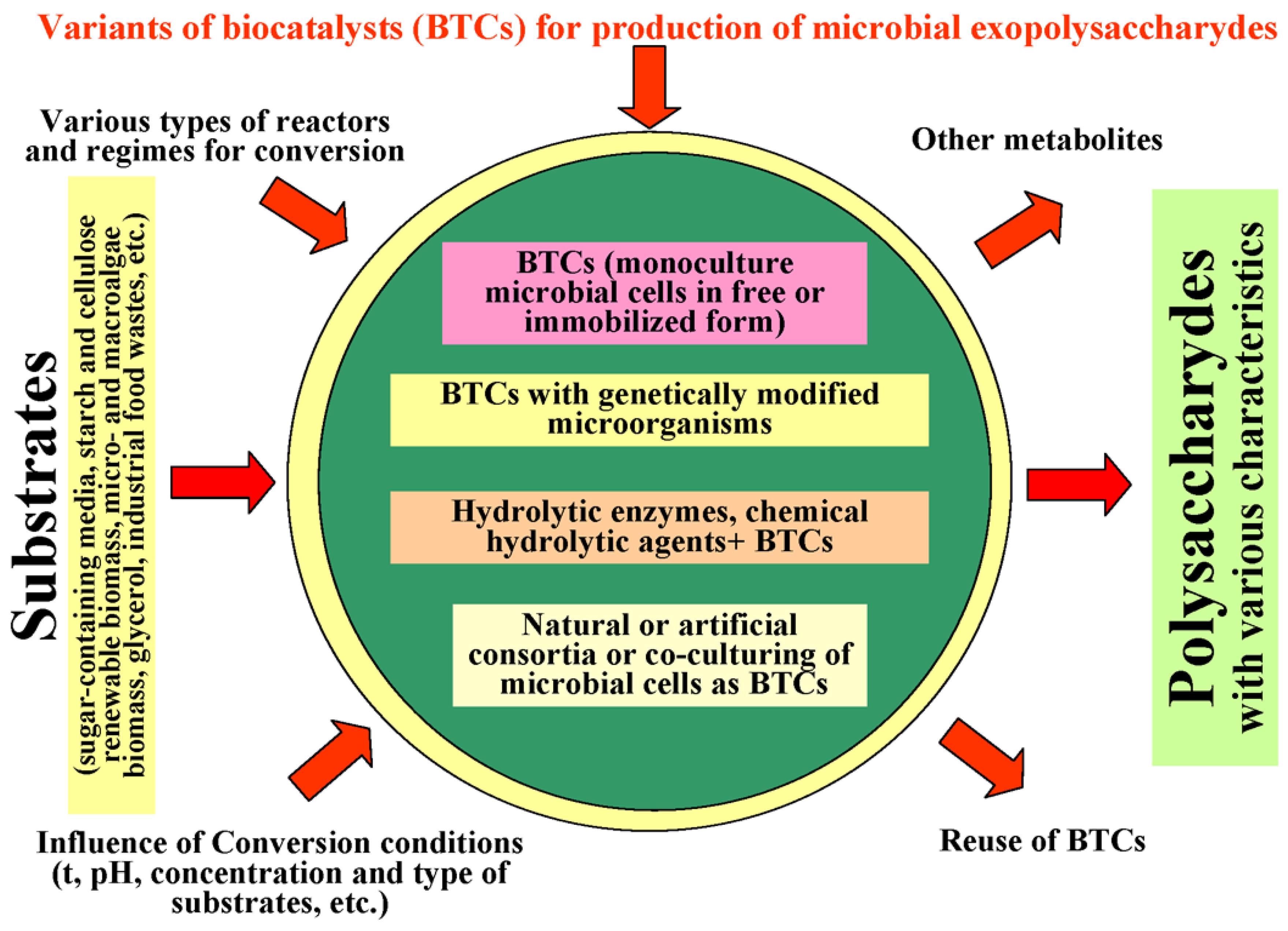
Biocatalytic synthesis of polysaccharides (PSs) is one of the promising and topical areas of the development of modern biotechnology. The variety of useful properties (the ability for gelation, the formation of viscous solutions, high adhesive ability, etc.) helps the PSs to find still newer applications in a plethora of fields. Biocatalysts determine the possible range of renewable raw materials which can be used as substrates for such synthesis, as well as the biochemistry of the process and the rate of molecular transformations. The functioning of biocatalysts can be optimized using the following main approaches of synthetic biology: the use of recombinant biocatalysts, the creation of artificial consortia, the combination of nano- and microbiocatalysts, and their immobilization. New biocatalysts can help expand the variety of the polysaccharides’ useful properties.
1. Introduction
2. Immobilized Biocatalysts for the Synthesis of Exopolysaccharides
| Biocatalyst [Reference]; Features of Cells |
Substrate Specificity of BTCs and Main Product (PS) | Conditions for BTCs’ Use | Rate of PS Synthesis |
|---|---|---|---|
| Dextran | |||
| Leuconostoc mesenteroides SF3 [33] | Sucrose-100.0 g/L Dextran-22.5 g/L |
pH 6.5, 30 °C, 20 h | 1.13 g/L/h |
| L. mesenteroides SF3 [34] | Sucrose-100.0 g/L Dextran-20.8 g/L |
pH 6.0, 28 °C, 24 h | 0.87 g/L/h |
| Weissella confusa R003 [35] | Sucrose-100.0 g/L/ Dextran-25.0 g/L |
pH 7.5, 30 °C, 24 h, 125 rpm | 1.04 g/L/h |
| L. mesenteroides N7 [36] | Sucrose 100.0 g/L/ Dextran-13.2 g/L |
30 °C, 24 h, 100 rpm |
0.55 g/L/h |
| L. citreum B-2 [37] | Sucrose 75.0 g/L/ Dextran-28.3 g/L |
30 °C, 48 h, 80 rpm | 0.59 g/L/h |
| L. pseudomesenteroides XG5 [38] | Sucrose-125.0 g/L/ Dextran-35.5 g/L |
30 °C, 48 h | 0.74 g/L/h |
| L. pseudomesenteroides G29 [39] | Sucrose-101.4 g/L/ Dextran-38.4 g/L |
pH 5.5, 30 °C, 10 h, 200 rpm |
3.84 g/L/h |
| W. cibaria 27 [40] | Sucrose-60.0 g/L Dextran-24.8 |
pH 6.2, 22 °C,24 h | 1.03 g/L/h |
| W. confusa Ck15 [41] | Sucrose-20.0 g/L Chickpea flour-280.0 g/L Dextran-14.9 g/L |
pH 6.3, 30 °C, 24 h |
0.62 g/L/h |
| L. mesenteroides NRRL B-512F [42] | Sucrose-200.0 g/L Milk permeate powder-150.0 g/L Dextran-42.9 g/L |
30 °C, 24 h | 1.79 g/L/h |
| L. mesenteroides MTCC 7337 [43] | Sugars in sugarcane juice-50.0 g/L Dextran-14.3 g/L |
pH 7.0, 30 °C, 72 h, 150 rpm |
0.20 g/L/h |
| L. pseudomesenteroides DSM20193 [44] | Sucrose-40.0 g/L Brewers’ spent grain-100.0 g/L Dextran-11.1 g/L |
pH 6.4, 25 °C, 24 h, 200 rpm |
0.46 g/L/h |
| Lactobacillus mali CUPV271 [45] | Sucrose-20 g/L/ Dextran-11.7 g/L |
pH 5.5; 28 °C; 48 h |
0.24 g/L/h |
| Weissella cibaria 10 M; mutant strain [46] | Sucrose-171.0 g/L Dextran-14.0 g/L |
pH 5.4–6.2, 25 °C, 24 h | 0.58 g/L/h |
| L. mesenteroides KIBGE-IB22M20; mutant strain [47] * | Sucrose-250.0 g/L Dextran-10.5 g/L |
pH 7.5, 25 °C, 12 h | 0.88 g/L/h |
| L. mesenteroides subsp. dextranicum B-5481; immobilized in PVA cryogel [48] | Sucrose-200.0 g/L/ Dextran-63.0 g/L |
pH 7.0, 28 °C, 15 h, 200 rpm |
4.20 g/L/h; reuse in 5 cycles |
| L. mesenteroides KIBGE HA1 [49]; immobilized in Ca-alginate gel |
Sucrose-100.0 g/L Dextran-8.0 g/L |
pH 5.0, 30 °C, 24 h 200 rpm |
0.33 g/L/h; reuse in 12 cycles |
| Levan | |||
| Bacillus subtilis MTCC 441 [50] | Sucrose-100.0 g/L Levan-30.4 g/L |
pH 7.0, 37 °C, 20 h, 150 rpm |
1.52 g/L/h |
| Z. mobilis PTCC 1718 [51] | Sucrose-300.0 g/L Levan-57.0 g/L |
28 °C, 48 h | 1.19 g/L/h |
| Bacillus sp. MTCC 1434 [52] | Sucrose-250.0 g/L Levan-61.0 g/L |
pH 6.0, 30 °C, 30 h, 100 rpm |
2.03 g/L/h |
| Brachybacterium phenoliresistens KX139300 [53] |
Sucrose-300.0 g/L Levan-8.6 g/L |
pH 7.8, 30 °C, 72 h, 150 rpm |
0.12 g/L/h |
| B. subtilis (NCIM 5021) [50] | Fresh coconut inflorescence sap (sugars g/L: sucrose–172.3 glucose-16.2, fructose-6.2) Levan-62.1 g/L |
pH 6.5, 35 °C, 17 h, 150 rpm |
3.65 g/L/h |
| Zymomonas mobilis CCT4494; immobilized in PVA cryogel [54] | Sucrose-300.0 g/L Levan-81.2 g/L |
pH 7.0, 30 °C, 12 h |
6.77 g/L/h |
| Z. mobilis CCT4494; immobilized in Ca-alginate gel [55] | Sucrose-350.0 g/L Levan-21.1 g/L |
pH 4.0, 30 °C, 24 h, 200 rpm |
0.88 g/L/h; reuse in 12 cycles |
| Z. mobilis CCT4494; immobilized on sugarcane bagasse [56] | Sucrose-350.0 g/L Levan-32.1 g/L |
pH 4.0, 30 °C, 24 h |
1.34 g/L/h; reuse in 12 cycles |
| Xanthan | |||
| Xanthomonas campestris AM001 [57] | Maltose-70.0 g/L Xanthan-40.7 g/L |
pH 7.0, 32 °C, 80 h, 600 rpm |
0.51 g/L/h |
| X. campestris ATCC 13951 [58] | Winery wastewater (sugars g/L-30.7) Xanthan-23.9 g/L |
pH 7.0, 29 °C, 96 h, 475 rpm |
0.25 g/L/h |
| X. campestris pv. campestris 1866 and 1867 [59] | Coconut shells or cocoa husks hydrolysates-20.0 g/L (25.0 g/L of sugars) Xanthan-3.6 g/L (coconut shells) Xanthan-4.5 g/L (cocoa husks) |
28 °C, 96 h, 250 rpm |
0.04–0.05 g/L/h |
| X. campestris WXLB-006; mutant strain [60] | Glycerol-40.0 g/L (+fed batch 1–3 g/L/h) Xanthan-33.9 g/L |
pH 7.0, 30 °C, 60 h, 200 rpm |
0.57 g/L/h |
| X. campestris ATCC 13951; immobilized in polyurethane foam [61] | Sucrose-50.0 g/L/ Xanthan-59.9 g/L |
28 °C, 96 h, 180 rpm |
0.62 g/L/h; reuse in 12 cycles |
| X. campestris PTCC 1473; self-immobilized cells on stainless-steel support [62] | Glucose-20.0 g/L/ Xanthan-3.5 g/L |
pH-6.9, 30 °C, 47 h, 180 rpm |
0.08 g/L/h |
| X. campestris PTCC1473; immobilized in calcium alginate–polyvinyl alcohol-boric acid gel [63] | Hydrolyzed starch-20.0 g/L Xanthan-9.2 g/L |
pH 6.6, 28 °C, 48 h, 180 rpm |
0.19 g/L/h; reuse in 3 cycles |
| X. campestris PTCC 1473; immobilized on plastic (polyethylene) support [64] | Glucose-20.0 g/L Xanthan-8.0 g/L |
pH 7.2, 30 °C, 48 h, 180 rpm |
0.17 g/L/h |
| Alginate | |||
| Pseudomonas stutzeri [65] | Sucrose-20.0 g/L Alginate-5.0 g/L |
pH 7.0, 30 °C, 600 h, 200 rpm | 0.008 g/L/h |
| Azotobacter vinelandii 12 [66] | Sucrose-35.0 g/L Alginate-2.7 g/L |
pH 7.2, 28 °C, 72 h, 210 rpm |
0.04 g/L/h |
| A. vinelandii, NRRL-14641 [67] | Apple peels-10.0 g/ Alginate-180.6 mg/g |
pH 7.5, 38 °C, 48 h |
3.76 mg/g/h. |
| A. vinelandii AT9; mutant strain [68] | Sucrose-20.0 g/L Alginate-3.8 g/L |
pH 7.2, 29 °C, 72 h, 200 rpm |
0.05 g/L/h |
| A. vinelandii B10436; immobilized in PVA cryogel [22] | Sucrose-30.0 g/L Alginate-2.5 g/L |
pH 7.0, 29 °C, 72 h, 200 rpm |
0.035 g/L/h; reuse in 5cycles |
| Pullulan | |||
| Aureobasidium melanogenum TN1-2 [69] | Sucrose-140.0 g/L Pullulan-114.0 g/L |
28 °C, 132 h, 250 rpm |
0.86 g/L/h |
| Rhodosporidium paludigenum PUPY-06 [70] | Sucrose-50.0 g/L Pullulan-21.0 g/L |
pH 6.0, 25 °C, 168 h, 150 rpm | 0.125 g/L/h |
| A. melanogenum A4 [71] | Maltose-303.0 g/L/ Pullulan-122.3 g/L |
pH 7.0, 30 °C, 120 h, 180 rpm | 1.02 g/L/h |
| A. pullulans Y-4137; immobilized in PVA cryogel [15] | Hydrolysate of Jerusalem artichoke tubers, hydrolysate of potato pulp, hydrolysate of Chlorella vulgaris biomass (glucose-15.0–25.0 g/L) Pullulan-3.5–16.8 g/L |
pH 5.5, 26 °C, 50 h, 200 rpm | 0.07–0.33 g/L/h reuse in 15 cycles |
| Bacterial cellulose (BC) | |||
| Lactobacillus hilgardii IITRKH159 [72] | Fructose-50.0 g/L BC-7.2 g/L |
28 °C, 384 h | 0.02 g/L/h |
| Komagataeibacter maltaceti [73] | Dextrin-8.0 g/L BC-6.5 g/L |
pH 6.0, 30 °C, 134 h | 0.05 g/L/h |
| K. nataicola [73] | Maltose-10.0 g/L BC-5.4 g/L |
pH 6.0, 30 °C, 134 h | 0.04 g/L/h |
| K. rhaeticus [74] | Acerola waste hydrolysate- 50.0 g/L + glucose 20.0 g/L BC-2.3 g/L |
pH 3.6, 30 °C, 288 h |
0.008 g/L/h |
| Medusomyces gisevii Sa-12 [75] | Miscanthus biomass hydrolysate (sugars-20.1–21.2 g/L) BC-1.24 g/L |
pH 4.0–4.6, 27 °C, 288 h | 0.004 g/L/h |
| Gluconacetobacter xylinus CGMCC 2955; mutant strain [76] | Glucose-25.0 g/L BC-4.3 g/L |
pH 6.0, 30 °C, 360 h |
0.012 g/L/h |
| K. xylinum; immobilized in PVA cryogel [32] | Glycerol-20.0 g/L BC-2.8 g/L |
pH 7.0, 28 °C, 168 h |
0.017 g/L/h |
| Aspen sawdust hydrolysate (sugars-42.0 g/L) BC-2.9 g/L |
0.017 g/L/h | ||
| Wheat straw hydrolysate (sugars-38.0 g/L) BC-3.6 g/L |
0.021 g/L/h | ||
| Rice straw hydrolysate (sugars-40.0 g/L) BC-3.5 g/L |
0.021 g/L/h | ||
| Jerusalem artichoke tubers hydrolysate (sugars-53.0 g/L) BC-4.5 g/L |
0.027 g/L/h | ||
| Chlorella vulgaris C1 biomass hydrolysate (sugars-45.1 g/L) BC-2.6 g/L |
0.015 g/L/h | ||
| Laminaria saccharina biomass hydrolysate (sugars-36.6 g/L) BC-0.07 g/L |
0.0004 g/L/h | ||
| Acanthophora muscoide biomass hydrolysate (sugars-56.0 g/L BC-0.4 g/L |
0.002 g/L/h | ||
| Ulva lactuca biomass hydrolysate (sugars-24.1 g/L) BC-0.08 g/L |
0.0005 g/L/h reuse in 10 cycles |
||
| Fructo-oligosaccharides | |||
| A. pullulans FRR 5284 [77] | Sucrose-500.0 g/L Fructo-oligosaccharides-306.3 g/L |
pH 5.5, 55 °C, 3 h | 102.1 g/L/h |
| A. pullulans CCY 27-1-94; immobilized on reticulated polyurethane foam [78] | Sucrose-200.0 g/L Fructo-oligosaccharides-108.2 g/L |
pH 5.5, 28 °C, 25 h, 150 rpm |
4.33 g/L/h |
| A. pullulans CCY 27-1-94; immobilized on walnut shell [78] | Sucrose-200.0 g/L Fructo-oligosaccharides-126.5 g/L |
pH 5.5; 28 °C; 36 h, 150 rpm |
3.51 g/L/h |
| Curdlan | |||
| Bacillus cereus PR3 [79] | Starch-100.0 g/L Curdlan-20.9 g/L |
30 °C, 96 h, 200 rpm |
0.22 g/L/h |
| Paenibacillus sp. NBR-10 [80] | Sucrose-50.0 g/L Curdlan-4.8 g/L |
pH 7.0, 35 °C, 84 h, 200 rpm |
0.06 g/L/h |
| Agrobacterium sp. ATCC 13140 [81] | Juice of discarded asparagus– 100 g/L (Sucrose-50.0 g/L) Curdlan-40.2 g/L |
pH 5.5, 30 °C, 168 h, 200 rpm |
0.24 g/L/h |
| Agrobacterium sp. DH-2 [82] | Cassava starch hydrolysate (sugars-90 g/L) Curdlan-28.4 g/L |
pH 5.5, 30 °C, 96 h, 250 rpm | 0.30 g/L/h |
| Agrobacterium sp. CGMCC 11546; mutant strain [83] | Sucrose-60.0 g/L Curdlan-48.0 g/L |
pH 5.0, 30 °C, 96 h, 280 rpm | 0.50 g/L/h |
| Agrobacterium sp. IFO 13140; immobilized on loofa sponge [84]; | Glucose-50.0 g/L Curdlan-17.8 g/L |
pH 6.5, 30 °C, 240 h, 150 rpm |
0.07 g/L/h; reuse in 5 cycles |
| Succinoglycan | |||
| Rhizobium radiobacter ATCC4720 [85] | Rice husk hydrolysate-100.0 g/L Succinoglycan-69.0 g/L |
pH 7.0, 30 °C, 72 h, 100 rpm | 0.96 g/L/h |
| R. radiobacter ATCC4720 [86] | Deproteinized whey–50.0 g/L Succinoglycan-13.7 g/L |
pH 7.0, 30 °C, 192 h,180 rpm |
0.07 g/L/h |
| R. radiobacter 18052 N-11; mutant strain [87] | Sucrose-70.0 g/L Succinoglycan-32.5 g/L |
pH 7.2, 30 °C, 72 h, 250 rpm |
0.45 g/L/h |
| Agrobacterium radiobacter NBRC 12665; immobilized on loofa sponge [88] | Sugarcane molasses-75.0 g/L Succinoglycan-14.1 g/L |
pH 7.0, 30 °C, 192 h, 180 rpm |
0.07 g/L/h; reuse in 5 cycles |
References
- Moradali, M.F.; Rehm, B.H. Bacterial biopolymers: From pathogenesis to advanced materials. Nat. Rev. Microbiol. 2020, 18, 195–210.
- Aanisah, N.; Wardhana, Y.W.; Chaerunisaa, A.Y.; Budiman, A. Review on modification of glucomannan as an excipient in solid dosage forms. Polymers 2022, 14, 2550.
- Melnikova, N.; Knyazev, A.; Nikolskiy, V.; Peretyagin, P.; Belyaeva, K.; Nazarova, N.; Liyaskina, E.; Malygina, D.; Revin, V. Wound healing composite materials of bacterial cellulose and zinc oxide nanoparticles with immobilized betulin diphosphate. Nanomaterials 2021, 11, 713.
- Liu, L.; Li, M.; Yu, M.; Shen, M.; Wang, Q.; Yu, Y.; Xie, J. Natural polysaccharides exhibit anti-tumor activity by targeting gut microbiota. Int. J. Biol. Macromol. 2019, 121, 743–751.
- Moscovici, M.; Balas, C. Bacterial polysaccharides versatile medical uses. In Polysaccharides of Microbial Origin: Biomedical Applications; Oliveira, J.M., Randhouani, H., Reis, R.L., Eds.; Springer Cham: Cham, Switzerland, 2022; pp. 859–891.
- Chakraborty, I.; Sen, I.K.; Mondal, S.; Rout, D.; Bhanja, S.K.; Maity, G.N.; Maity, P. Bioactive polysaccharides from natural sources: A review on the antitumor and immunomodulating activities. Biocatal. Agric. Biotechnol. 2019, 22, 101425.
- Atykyan, N.; Revin, V.; Shutova, V. Raman and FT-IR Spectroscopy investigation the cellulose structural differences from bacteria Gluconacetobacter sucrofermentans during the different regimes of cultivation on a molasses media. AMB Express 2020, 10, 84.
- Stumpf, T.R.; Yang, X.; Zhang, J.; Cao, X. In situ and ex situ modifications of bacterial cellulose for applications in tissue engineering. Mater. Sci. Eng. C 2018, 82, 372–383.
- Schmid, J. Recent insights in microbial exopolysaccharide biosynthesis and engineering strategies. Curr. Opin. Biotechnol. 2018, 53, 130–136.
- Sun, X.; Zhang, J. Bacterial exopolysaccharides: Chemical structures, gene clusters and genetic engineering. Int. J. Biol. Macromol. 2021, 173, 481–490.
- Jurášková, D.; Ribeiro, S.C.; Silva, C.C. Exopolysaccharides produced by lactic acid bacteria: From biosynthesis to health-promoting properties. Foods 2022, 11, 156.
- Wang, Y.; Feng, K.; Jin, J.; Murad, M.S.; Mu, G.; Wu, X. Comparison on properties between normal and A2 bovine milk fermented using commercial bacteria mixed with/without two probiotics from human milk. Int. J. Biol. Macromol 2022, 216, 105–113.
- Xie, Z.; Meng, K.; Yang, X.; Liu, J.; Yu, J.; Zheng, C.; Cao, W.; Liu, H. Identification of a quorum sensing system regulating capsule polysaccharide production and biofilm formation in Streptococcus zooepidemicus. Front. Cell. Infect. Microbiol. 2019, 9, 121.
- Wang, X.L.; Chen, N.; Li, J.; Han, C.F.; Wang, S.; Hao, L.M.; Jia, S.R.; Han, P.P. The effects of quorum sensing molecule farnesol on the yield and activity of extracellular polysaccharide from Grifola frondosa in liquid fermentation. Int. J. Biol. Macromol. 2021, 191, 377–384.
- Senko, O.V.; Efremenko, E.N. Highly concentrated populations of Aureobasidium pullulans cells in biocatalytic pullulan production processes. Catal. Ind. 2017, 9, 344–348.
- Zur, J.; Wojcieszynska, D.; Guzik, U. Metabolic responses of bacterial cells to immobilization. Molecules 2016, 21, 958.
- Bergmaier, D.; Champagne, C.P.; Lacroix, C. Exopolysaccharide production during batch cultures with free and immobilized Lactobacillus rhamnosus RW-9595M. J. Appl. Microbiol. 2003, 95, 1049–1057.
- Bergmaier, D.; Champagne, C.P.; Lacroix, C. Growth and exopolysaccharide production during free and immobilized cell chemostat culture of Lactobacillus rhamnosus RW-9595M. J. Appl. Microbiol. 2005, 98, 272–284.
- EL-Gizawy, S.A.; Barakat, O.S.; Sharaf, O.M.; EL-Shafei, K.; Fathy, F.A.; EL-Sayed, H.S. Effect of growth conditions on the production of exopolysaccharides by microencapsulated Lactobacillus bulgaricus and use it to improve quality of Kareish cheese. J. Appl. Sci. Res. 2013, 9, 1097–1109.
- Adebayo-Tayo, B.; Agidigbi, O.; Alao, S. Comparative influence of immobilization medium and mutation on EPS-production by L. plantarum MK 02 isolated from fermented milk. Trakia J. Sci. 2017, 1, 30.
- Ismail, B.; Nampoothiri, K.M. Exopolysaccharide production and prevention of syneresis in starch using encapsulated probiotic Lactobacillus plantarum. Food Technol. Biotechnol. 2010, 48, 484–489.
- Efremenko, E.N. Immobilized Cells: Biocatalysts and Processes; RIOR: Moscow, Russia, 2018; p. 409.
- Nugroho, D.A.; Aji, P. Characterization of nata de coco produced by fermentation of immobilized Acetobacter xylinum. Agric. Agric. Sci. Procedia 2015, 3, 278–282.
- Matyash, M.; Despang, F.; Ikonomidou, C.; Gelinsky, M. Swelling and mechanical properties of alginate hydrogels with respect to promotion of neural growth. Tissue Eng. Part C Methods 2014, 20, 401–411.
- Maslova, O.; Stepanov, N.; Senko, O.; Efremenko, E. Production of various organic acids from different renewable sources by immobilized cells in the regimes of separate hydrolysis and fermentation (SHF) and simultaneous saccharification and fermentation (SSF). Bioresour. Technol. 2019, 272, 1–9.
- Maslova, O.; Senko, O.; Stepanov, N.; Gladchenko, M.; Gaydamaka, S.; Akopyan, A.; Eseva, E.; Anisimov, A.; Efremenko, E. Sulfur containing mixed wastes in anaerobic processing by new immobilized synthetic consortia. Bioresour. Technol. 2022, 362, 127794.
- Efremenko, E.; Stepanov, N.; Maslova, O.; Senko, O.; Aslanli, A.; Lyagin, I. “Unity and struggle of opposites” as a basis for the functioning of synthetic bacterial immobilized consortium that continuously degrades organophosphorus pesticides. Microorganisms 2022, 10, 1394.
- Efremenko, E.N.; Tatarinova, N.Y. The effect of long-term preservation of bacterial cells immobilized in poly(vinyl alcohol) cryogel on their viability and biosynthesis of target metabolites. Microbiology 2007, 76, 336–341.
- Senko, O.; Gladchenko, M.; Maslova, O.; Efremenko, E. Long-term storage and use of artificially immobilized anaerobic sludge as a powerful biocatalyst for conversion of various wastes including those containing xenobiotics to biogas. Catalysts 2019, 9, 326.
- Efremenko, E.N.; Nikolskaya, A.B.; Lyagin, I.V.; Senko, O.V.; Makhlis, T.A.; Stepanov, N.A.; Maslova, O.V.; Mamedova, F.; Varfolomeev, S.D. Production of biofuels from pretreated microalgae biomass by anaerobic fermentation with immobilized Clostridium acetobutylicum cells. Bioresour. Technol. 2012, 114, 342–348.
- Stepanov, N.; Efremenko, E. Immobilised cells of Pachysolen tannophilus yeast for ethanol production from crude glycerol. New Biotechnol. 2017, 34, 54–58.
- Stepanov, N.; Efremenko, E. “Deceived” concentrated immobilized cells as biocatalyst for intensive bacterial cellulose production from various sources. Catalysts 2018, 8, 33.
- Díaz-Montes, E.; Yáñez-Fernández, J.; Castro-Muñoz, R. Microfiltration-mediated extraction of dextran produced by Leuconostoc mesenteroides SF3. Food Bioprod. Process. 2020, 119, 317–328.
- Diana, C.R.; Humberto, H.S.; Jorge, Y.F. Structural characterization and rheological properties of dextran produced by native strains isolated of Agave salmiana. Food Hydrocoll. 2019, 90, 1–8.
- Netsopa, S.; Niamsanit, S.; Sakloetsakun, D.; Milintawisamai, N. Characterization and rheological behavior of dextran from Weissella confusa R003. Int. J. Polym. Sci. 2018, 2018, 5790526.
- Ma’unatin, A.; Harijono, H.; Zubaidah, E.; Rifa’i, M. Dextran production using Leuconostoc mesenteroides strains isolated from Borassus flabellifer sap. Biodivers. J. Biol. Divers. 2022, 23, 1154–1158.
- Feng, F.; Zhou, Q.; Yang, Y.; Zhao, F.; Du, R.; Han, Y.; Xiao, H.; Zhou, Z. Characterization of highly branched dextran produced by Leuconostoc citreum B-2 from pineapple fermented product. Int. J. Biol. Macromol. 2018, 113, 45–50.
- Zhou, Q.; Feng, F.; Yang, Y.; Zhao, F.; Du, R.; Zhou, Z.; Han, Y. Characterization of a dextran produced by Leuconostoc pseudomesenteroides XG5 from homemade wine. Int. J. Biol. Macromol. 2018, 107, 2234–2241.
- Wang, Y.; Sun, T.; Wang, Y.; Wu, H.; Fang, Y.; Ma, J.; Jiang, M. Production and characterization of insoluble α-1, 3-linked glucan and soluble α-1, 6-linked dextran from Leuconostoc pseudomesenteroides G29. Chin. J. Chem. Eng. 2021, 39, 211–218.
- Yu, Y.J.; Chen, Z.; Chen, P.T.; Ng, I.S. Production, characterization and antibacterial activity of exopolysaccharide from a newly isolated Weissella cibaria under sucrose effect. J. Biosci. Bioeng. 2018, 126, 769–777.
- Galli, V.; Venturi, M.; Coda, R.; Maina, N.H.; Granchi, L. Isolation and characterization of indigenous Weissella confusa for in situ bacterial exopolysaccharides (EPS) production in chickpea sourdough. Food Res. Int. 2020, 138, 109785.
- Esmaeilnejad-Moghadam, B.; Mokarram, R.R.; Hejazi, M.A.; Khiabani, M.S.; Keivaninahr, F. Low molecular weight dextran production by Leuconostoc mesenteroides strains: Optimization of a new culture medium and the rheological assessments. Bioact. Carbohydr. Diet. Fibre 2019, 18, 100181.
- Rahman, S.S.A.; Venkatachalam, P.; Karuppiah, S. Cost-effective production of dextran using Saccharum officinarum juice (SOJ) as a potential feedstock: Downstream processing and characterization. Biomass Convers. Biorefin. 2022, 12, 4863–4875.
- Koirala, P.; Maina, N.H.; Nihtilä, H.; Katina, K.; Coda, R. Brewers’ spent grain as substrate for dextran biosynthesis by Leuconostoc pseudomesenteroides DSM20193 and Weissella confusa A16. Microb. Cell Fact. 2021, 20, 23.
- Llamas-Arriba, M.G.; Puertas, A.I.; Prieto, A.; López, P.; Cobos, M.; Miranda, J.I.; Marieta, C.; Ruas-Madiedo, P.; Dueñas, M.T. Characterization of dextrans produced by Lactobacillus mali CUPV271 and Leuconostoc carnosum CUPV411. Food Hydrocoll. 2019, 89, 613–622.
- Hu, Y.; Gänzle, M.G. Effect of temperature on production of oligosaccharides and dextran by Weissella cibaria 10 M. Int. J. Food Microbiol. 2018, 280, 27–34.
- Siddiqui, N.N.; Aman, A.; Silipo, A.; Qader, S.A.U.; Molinaro, A. Structural analysis and characterization of dextran produced by wild and mutant strains of Leuconostoc mesenteroides. Carbohydr. Polym. 2014, 99, 331–338.
- Stepanov, N.A.; Senko, O.V.; Efremenko, E.N. Biocatalytic production of extracellular exopolysaccharide dextran synthesized by cells of Leuconostoc mesenteroides. Catal. Ind. 2017, 9, 339–343.
- Qader, S.A.U.; Aman, A. Low molecular weight dextran: Immobilization of cells of Leuconostoc mesenteroides KIBGE HA1 on calcium alginate beads. Carbohydr. Polym. 2012, 87, 2589–2592.
- Veerapandian, B.; Shanmugam, S.R.; Varadhan, S.; Sarwareddy, K.K.; Mani, K.P.; Ponnusami, V. Levan production from sucrose using chicken feather peptone as a low cost supplemental nutrient source. Carbohydr. Polym. 2020, 227, 115361.
- Taran, M.; Lotfi, M.; Safaei, M. Optimal conditions for levan biopolymer production and its use in the synthesis of bactericidal levan-zno nanocomposite. Biotechnologia 2019, 100, 397–405.
- Ramya, P.; Sangeetha, D.; Anooj, E.S.; Gangadhar, L. Studies on the production and optimization of levan from Bacillus sp. Ann. Trop. Med. Public Health 2020, 23, 1188–1197.
- Moussa, T.A.; Al-Qaysi, S.A.; Thabit, Z.A.; Kadhem, S.B. Microbial levan from Brachybacterium phenoliresistens: Characterization and enhancement of production. Process Biochem. 2017, 57, 9–15.
- Lorenzetti, M.F.S.; Moro, M.R.; García-Cruz, C.H. Alginate/PVA beads for levan production by Zymomonas mobilis. J. Food Process Eng. 2015, 38, 31–36.
- Santos, V.A.Q.; Cruz, C.H.G. Ethanol and levan production by sequential bath using Zymomonas mobilis immobilized on alginate and chitosan beads. Acta Sci.–Technol. 2016, 38, 263–271.
- Santos, V.A.Q.; Cruz, C.H.G. Zymomonas mobilis immobilized on loofa sponge and sugarcane bagasse for levan and ethanol production using repeated batch fermentation. Braz. J. Chem. Eng. 2017, 34, 407–418.
- Mohsin, A.; Akyliyaevna, K.A.; Zaman, W.Q.; Hussain, M.H.; Mohsin, M.Z.; Al-Rashed, S.; Tan, X.; Tian, X.; Aida, K.; Tariq, M.; et al. Kinetically modelled approach of xanthan production using different carbon sources: A study on molecular weight and rheological properties of xanthan. Int. J. Biol. Macromol. 2021, 193, 1226–1236.
- Rončević, Z.; Grahovac, J.; Dodić, S.; Vučurović, D.; Dodić, J. Utilisation of winery wastewater for xanthan production in stirred tank bioreactor: Bioprocess modelling and optimisation. Food Bioprod. Process. 2019, 117, 113–125.
- da Silva, J.A.; Cardoso, L.G.; de Jesus Assis, D.; Gomes, G.V.P.; Oliveira, M.B.P.P.; de Souza, C.O.; Druzian, J.I. Xanthan gum production by Xanthomonas campestris pv. campestris IBSBF 1866 and 1867 from lignocellulosic agroindustrial wastes. Appl. Biochem. Biotechnol. 2018, 186, 750–763.
- Wang, Z.; Wu, J.; Gao, M.J.; Zhu, L.; Zhan, X.B. High production of xanthan gum by a glycerol-tolerant strain Xanthomonas campestris WXLB-006. Prep. Biochem. Biotechnol. 2017, 47, 468–472.
- Mesquita, R.A.; Hassemer, G.; Marchiori, V.; Kiedis, J.; Valduga, E.; Junges, A.; Malvessi, E.; Cansian, R.L.; Zeni, J. Synthesis of xanthan gum from Xanthomonas campestris immobilized in polyurethane. Ind. Biotechnol. 2018, 14, 276–281.
- Nejadmansouri, M.; Razmjooei, M.; Safdarianghomsheh, R.; Shad, E.; Delvigne, F.; Khalesi, M. Semi-continuous production of xanthan in biofilm reactor using Xanthomonas campestris. J. Biotechnol. 2021, 328, 1–11.
- Niknezhad, S.V.; Asadollahi, M.A.; Zamani, A.; Biria, D. Production of xanthan gum by free and immobilized cells of Xanthomonas campestris and Xanthomonas pelargonii. Int. J. Biol. Macromol. 2016, 82, 751–756.
- Nejadmansouri, M.; Shad, E.; Razmjooei, M.; Safdarianghomsheh, R.; Delvigne, F.; Khalesi, M. Production of xanthan gum using immobilized Xanthomonas campestris cells: Effects of support type. Biochem. Eng. J. 2020, 157, 107554.
- Meena, S.; Tripathi, A.D.; Ts, R.L. Optimization and characterization of Alginic acid synthesized from a novel strain of Pseudomonas stutzeri. Biotechnol. Rep. 2020, 27, e00517.
- Dudun, A.A.; Akoulina, E.A.; Zhuikov, V.A.; Makhina, T.K.; Voinova, V.V.; Belishev, N.V.; Khaydapova, D.D.; Shaitan, K.V.; Bonartseva, G.A.; Bonartsev, A.P. Competitive biosynthesis of bacterial alginate using Azotobacter vinelandii 12 for tissue engineering applications. Polymers 2022, 14, 131.
- Saeed, S.; Mehmood, T.; Irfan, M. Statistical optimization of cultural parameters for the optimized production of alginic acid using apple (Malus domestica) peels through solid-state fermentation. Biomass Convers. Biorefin. 2020.
- García, A.; Castillo, T.; Ramos, D.; Ahumada-Manuel, C.L.; Núñez, C.; Galindo, E.; Büchs, J.; Peña, C. Molecular weight and viscosifying power of alginates produced by mutant strains of Azotobacter vinelandii under microaerophilic conditions. Biotechnol. Rep. 2020, 26, e00436.
- Jiang, H.; Xue, S.J.; Li, Y.F.; Liu, G.L.; Chi, Z.M.; Hu, Z.; Chi, Z. Efficient transformation of sucrose into high pullulan concentrations by Aureobasidium melanogenum TN1-2 isolated from a natural honey. Food Chem. 2018, 257, 29–35.
- Singh, R.S.; Kaur, N. Biochemical and molecular characterization of a new pullulan producer Rhodosporidium paludigenum PUPY-06. J. Appl. Biol. Biotechnol. 2018, 6, 2–7.
- Chen, G.; Zhu, Y.; Zhang, G.; Liu, H.; Wei, Y.; Wang, P.; Xian, M.; Xiang, H.; Zhang, H. Optimization and characterization of pullulan production by a newly isolated high-yielding strain Aureobasidium melanogenum. Prep. Biochem. Biotechnol. 2019, 49, 557–566.
- Khan, H.; Kadam, A.; Dutt, D. Studies on bacterial cellulose produced by a novel strain of Lactobacillus genus. Carbohydr. Polym. 2020, 229, 115513.
- Greser, A.B.; Avcioglu, N.H. Optimization and physicochemical characterization of bacterial cellulose by Komagataeibacter nataicola and Komagataeibacter maltaceti strains isolated from grape, thorn apple and apple vinegars. Arch. Microbiol. 2022, 204, 465.
- Leonarski, E.; Cesca, K.; Pinto, C.C.; González, S.Y.; de Oliveira, D.; Poletto, P. Bacterial cellulose production from acerola industrial waste using isolated kombucha strain. Cellulose 2022, 29, 7613–7627.
- Skiba, E.A.; Gladysheva, E.K.; Golubev, D.S.; Budaeva, V.V.; Aleshina, L.A.; Sakovich, G.V. Self-standardization of quality of bacterial cellulose produced by Medusomyces gisevii in nutrient media derived from Miscanthus biomass. Carbohydr. Polym. 2021, 252, 117178.
- Liu, M.; Li, S.; Xie, Y.; Jia, S.; Hou, Y.; Zou, Y.; Zhong, C. Enhanced bacterial cellulose production by Gluconacetobacter xylinus via expression of Vitreoscilla hemoglobin and oxygen tension regulation. Appl. Microbiol. Biotechnol. 2018, 102, 1155–1165.
- Khatun, M.S.; Harrison, M.D.; Speight, R.E.; O’Hara, I.M.; Zhang, Z. Efficient production of fructo-oligosaccharides from sucrose and molasses by a novel Aureobasidium pullulan strain. Biochem. Eng. J. 2020, 163, 107747.
- Castro, C.C.; Nobre, C.; Duprez, M.E.; De Weireld, G.; Hantson, A.L. Screening and selection of potential carriers to immobilize Aureobasidium pullulans cells for fructo-oligosaccharides production. Biochem. Eng. J. 2017, 118, 82–90.
- Prakash, S.; Rajeswari, K.; Divya, P.; Ferlin, M.; Rajeshwari, C.T.; Vanavil, B. Optimization and production of curdlan gum using Bacillus cereus pr3 isolated from rhizosphere of leguminous plant. Prep. Biochem. Biotechnol. 2018, 48, 408–418.
- El-Sayed, M.H.; Arafat, H.H.; Elsehemy, I.A.; Basha, M. Optimization, purification and physicochemical characterization of curdlan produced by Paenibacillus sp. strain NBR-10. Biosci. Biotechnol. Res. Asia 2016, 13, 901–909.
- Anane, R.F.; Sun, H.; Zhao, L.; Wang, L.; Lin, C.; Mao, Z. Improved curdlan production with discarded bottom parts of Asparagus spear. Microb. Cell Fact. 2017, 16, 59.
- Wan, J.; Shao, Z.; Jiang, D.; Gao, H.; Yang, X. Curdlan production from cassava starch hydrolysates by Agrobacterium sp. DH-2. Bioprocess Biosyst. Eng. 2022, 45, 969–979.
- Gao, H.; Xie, F.; Zhang, W.; Tian, J.; Zou, C.; Jia, C.; Jin, M.; Huang, J.; Chang, Z.; Yang, X.; et al. Characterization and improvement of curdlan produced by a high-yield mutant of Agrobacterium sp. ATCC 31749 based on whole-genome analysis. Carbohydr. Polym. 2020, 245, 116486.
- Ortiz Martinez, C.; Pereira Ruiz, S.; Carvalho Fenelon, V.; Rodrigues de Morais, G.; Luciano Baesso, M.; Matioli, G. Characterization of curdlan produced by Agrobacterium sp. IFO 13140 cells immobilized in a loofa sponge matrix, and application of this biopolymer in the development of functional yogurt. J. Sci. Food Agric. 2016, 96, 2410–2417.
- Pedroso, G.B.; Silva, L.O.; Araujo, R.B.; Saldanha, L.F.; Denardi, L.; Martins, A.F. An innovative approach for the biotechnological production of succinoglycan from rice husks. Ind. Crops Prod. 2019, 137, 615–627.
- de Oliveira Delani, T.C.; Sampaio, A.R.; da Silva Palácios, R.; Sato, F.; Ruiz, S.P.; de Oliveira Petkowicz, C.L.; Reichembach, L.H.; Miyoshi, J.H.; Nascimento, M.G.; Matioli, G. Rheological and structural characterization of succinoglycan obtained by bioconversion of the agroindustrial residue deproteinized whey. Res. Sq. 2022.
- Gao, H.; Yang, L.; Tian, J.; Huang, L.; Huang, D.; Zhang, W.; Xie, F.; Niu, Y.; Jin, M.; Jia, C.; et al. Characterization and rheological properties analysis of the succinoglycan produced by a high-yield mutant of Rhizobium radiobacter ATCC 19358. Int. J. Biol. Macromol. 2021, 166, 61–70.
- Ruiz, S.P.; Martinez, C.O.; Noce, A.S.; Sampaio, A.R.; Baesso, M.L.; Matioli, G. Biosynthesis of succinoglycan by Agrobacterium radiobacter NBRC 12665 immobilized on loofa sponge and cultivated in sugar cane molasses. Structural and rheological characterization of biopolymer. J. Mol. Catal. B Enzym. 2015, 122, 15–28.

