 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Sirius Huang | -- | 2872 | 2022-11-21 01:42:30 |
Video Upload Options
The megafaunal wolf (Canis cf. lupus) was a Late Pleistocene – early Holocene hypercarnivore similar in size to a large extant gray wolf. It had a shorter, broader palate with large carnassial teeth relative to its overall skull size. This adaptation allowed it to prey and scavenge on Pleistocene megafauna. Such an adaptation is an example of phenotypic plasticity. It was once distributed across the northern Holarctic.
1. Name
In 2007, the words "megafaunal" and "wolf" appeared separated in the title of a study.[1] In 2013, one of the study co-authors first used the term "megafaunal wolf" in a media release.[2]
2. Taxonomy
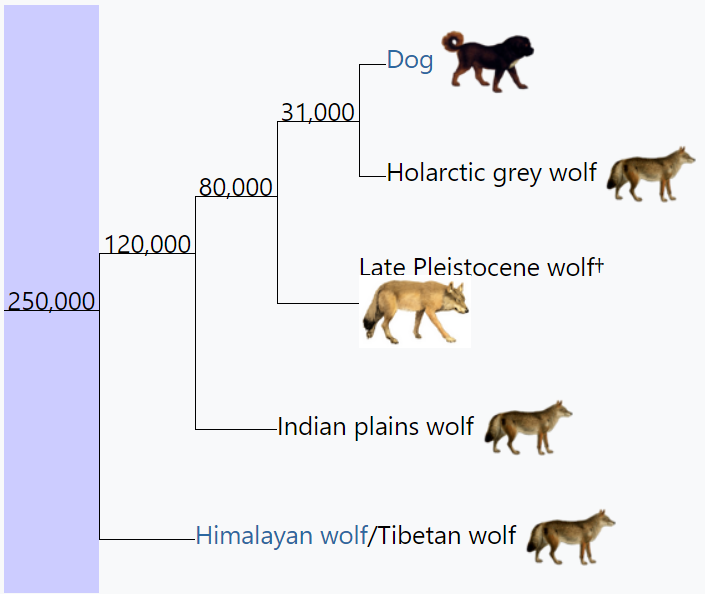
Phylogenetic tree with timing in years for Canis lupus[3]
The megafaunal wolf (Canis cf. lupus, where cf. in Latin means confer, uncertain) has not yet been taxonomically classified but based on genetic analysis is believed to be an ecomorph of Canis lupus.[1][4] The ancient wolf specimens from Europe have been classified as Canis lupus spelaeus Goldfuss, 1823 - the cave wolf.[5]
3. Differences
3.1. Populations
Ecological factors including habitat type, climate, prey specialization, and predatory competition will greatly influence gray wolf genetic population structure and cranio-dental plasticity.[6][7][8][9][10][11][12][13][14] Therefore, within the Pleistocene gray wolf population, the variations between local environments would have encouraged a range of wolf ecotypes that were genetically, morphologically, and ecologically distinct from one another.[14]
3.2. Types
In 2010, a study compared a 230 base pair sequence of the mitochondrial control region from 24 ancient wolf specimens from western Europe dated between 44,000–1,200 ybp with those of modern gray wolves. The sequences could be represented on a phylogenetic tree, which formed two haplogroups that were separated from each other by 5 mutations. Haplogroup 1 was monophyletic (indicating they share a common ancestor). Haplogroup 2 was basal to haplogroup 1, which indicated a more ancient lineage, and it was not monophyletic.[4][15] The ancient wolf samples from western Europe all belonged to haplogroup 2, indicating haplogroup 2 predominance in this region for over 40,000 years before and after the Last Glacial Maximum.[4]
4. Physical Description
4.1. Skull and Dentition
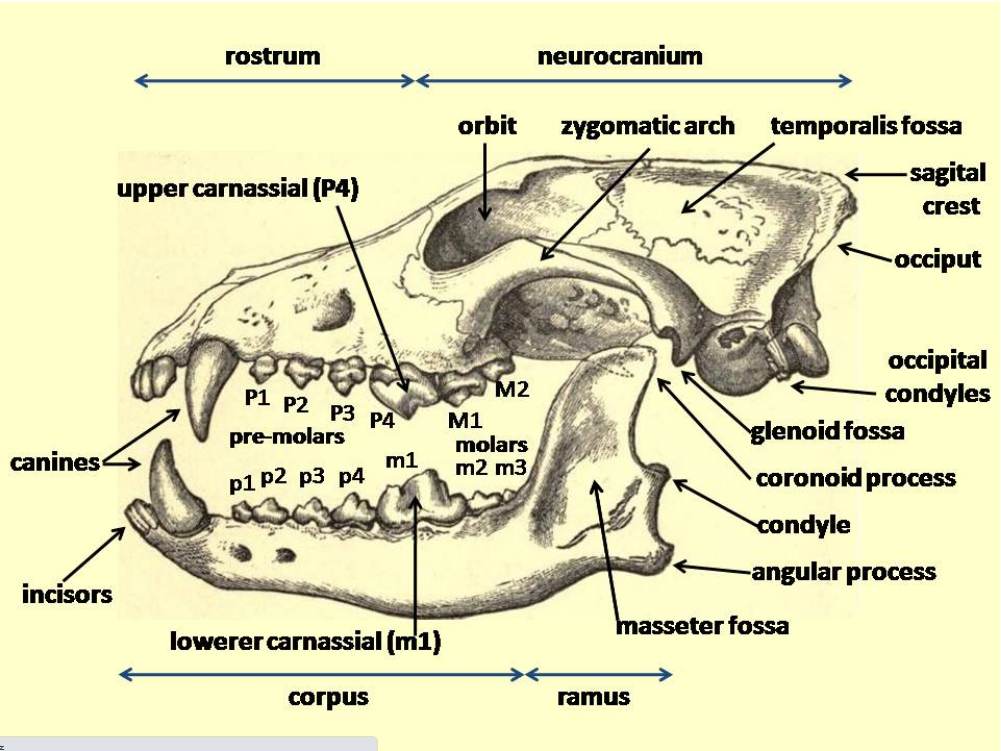
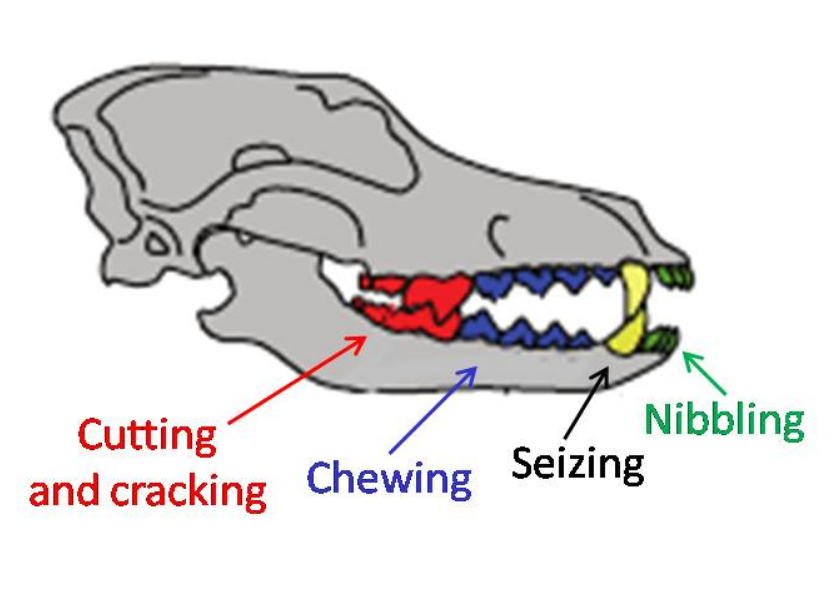
Megafaunal wolves were similar in physical size to other Pleistocene-era wolves and large extant gray wolves, but with stronger jaws and teeth. They tended to have short, broad palates with large carnassials relative to their overall skull size. These features suggest a wolf adapted for producing relatively large bite forces. The short, broad rostrum increased the mechanical advantage of a bite made with the canine teeth and strengthened the skull against torsional stresses caused by struggling prey. Relatively deep jaws are characteristic of habitual bone crackers, such as spotted hyenas, as well as canids that take prey as large as or larger than themselves. Overall, these features indicate that megafaunal wolves were more specialized than modern gray wolves in killing and consuming relatively large prey, and scavenging.[1]:1147
In comparison to other gray wolves, megafaunal wolf samples include many more individuals with moderately to heavily worn teeth, and significantly greater numbers of broken teeth. The distribution of fractures across the tooth row differs as well, with these wolves having much higher fracture frequencies of incisors, carnassials, and molars. A similar pattern was observed in spotted hyenas, suggesting that increased incisor and carnassial fracture reflects habitual bone consumption, because bones are gnawed with incisors and subsequently cracked with the cheek teeth.[1]:1148
In 2014, a study of the morphology of wolf (Canis lupus) remains from Europe dating from the Middle-Late Pleistocene and Holocene indicated that the size of the lower carnassial teeth did not fluctuate directly with changes in climate but possibly with the spread of cold megafauna, and therefore in the dietary regime. The lower carnassial length can be used to estimate carnivore body size.[16]
In 2015, a study looked at specimens of all of the carnivore species from Rancho La Brea, California, including remains of the large wolf Canis dirus that was also a megafaunal hypercarnivore. The evidence suggests that these carnivores were not food-stressed just before extinction, and that carcass utilization was less than among large carnivores today. The high incidence of tooth breakage likely resulted from the acquisition and consumption of larger prey.[17]
5. Diet
Isotopic bone collagen analysis of the specimens indicated that they ate horse, bison, woodland muskox, and mammoth — i.e., Pleistocene megafauna. This supports the conclusion that they were capable of killing and dismembering large prey. Compared with Pleistocene and extant gray wolves, the megafaunal wolf was hypercarnivorous, with a craniodental morphology more capable of capturing, dismembering, and consuming the bones of very large mega-herbivores. When their prey disappeared, this wolf did as well, resulting in a significant loss of phenotypic and genetic diversity within the species.[1]:1148
6. Habitat
Based on the morphological and genetic evidence, the megafaunal wolf's distribution was across the northern Holarctic.
6.1. Paleoecology

The last glacial period, commonly referred to as the "Ice Age", spanned 125,000[19] to 14,500[20] years ago and was the most recent glacial period within the current ice age which occurred during the last years of the Pleistocene era.[19] The Ice Age reached its peak during the last glacial maximum, when ice sheets commenced advancing from 33,000 years BP and reached their maximum positions 26,500 years BP. Deglaciation commenced in the Northern Hemisphere approximately 19,000 years BP, and in Antarctica approximately 14,500 years BP, which is consistent with evidence that this was the primary source for an abrupt rise in the sea level 14,500 years ago.[20]
A vast mammoth steppe stretched from Spain across Eurasia and over the Bering land bridge into Alaska and the Yukon, where it was stopped by the Wisconsin glaciation. This land bridge existed because more of the planet's water was locked up in glaciation than now, and therefore the sea levels were lower. When the sea levels began to rise, this bridge was inundated around 11,000 years BP.[21] During the last glacial maximum, the continent of Europe was much colder and drier than it is today, with polar desert in the north and the remainder steppe or tundra. Forest and woodland was almost non-existent, except for isolated pockets in the mountain ranges of southern Europe.[22]
The fossil evidence from many continents points to the extinction mainly of large animals at or near the end of the last glaciation. These animals have been termed Pleistocene megafauna. The most common definition of megafauna is an animal with an adult body weight of over 44 kg. Across Eurasia, the straight-tusked elephant became extinct between 100,000–50,000 years BP. The hippopotamus, interglacial rhinoceros (Stephanorhinus), cave bear (Ursus spelaeus), and heavy-bodied Asian antelope (Spirocerus) died out between 50,000-16,000 years BP. The spotted hyena, woolly rhinoceros, and mammoths died out between 16,000-11,500 years BP. The musk ox died out after 11,500 BP, as did the giant deer (Megaloceros), with the last pocket having survived until about 7,700 years BP in western Siberia.[23] A pocket of mammoths survived on Wrangel Island until 4,500 years BP.[24] As some species became extinct, so too did their predators. Among the top predators, the sabre-toothed cat (Homotherium) died out 28,000 years BP,[25] the Eurasian cave lion 11,900 years BP,[26] and the leopard in Europe died out 27,000 years BP.[27] The Late Pleistocene was characterized by a series of severe and rapid climate oscillations with regional temperature changes of up to 16° C, which has been correlated with megafaunal extinctions. There is no evidence of megafaunal extinctions at the height of the LGM, indicating that increasing cold and glaciation were not factors. Multiple events appear to also involve the rapid replacement of one species by one within the same genus, or one population by another within the same species, across a broad area.[28]
Modern humans' ancestors first appeared in East Africa 195,000 years ago.[29] Some migrated out of Africa 60,000 years ago, with one group reaching Central Asia 50,000 years ago.[30] From there they reached Europe, with human remains dated 43,000-45,000 years BP discovered in Italy,[31] Britain,[32] and in the European Russian Arctic 40,000 years ago.[33][34] Remains of mammoth that had been hunted by humans 45,000 YBP have been found at Yenisei Bay in the central Siberian Arctic.[35] Another group left Central Asia and reached the Yana River, Siberia, well above the Arctic circle 27,000 years ago.[36] Modern humans then made their way across the Bering land bridge and into North America between 20,000-11,000 years ago, after the Wisconsin glaciation had retreated but before the Bering land bridge became inundated by the sea.[37] These people then populated the Americas. In the Fertile crescent the first agriculture was developing 11,500 years ago.[38]
In this environment, the non-megafauna-specialist haplogroup 1 wolf became more numerous than the haplogroup 2 megafaunal wolf in the Old World and in the New World was unchallenged by the latter due to its prior extinction there.
6.2. Beringia
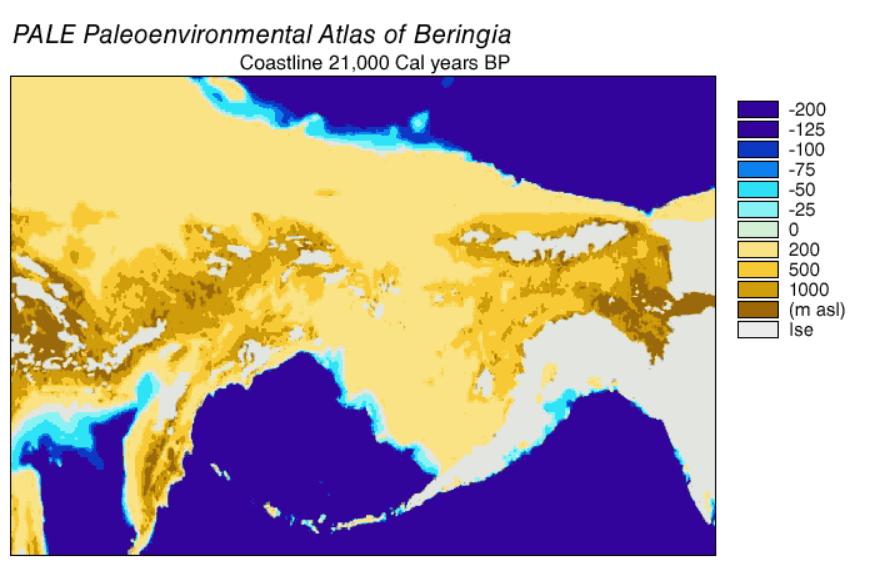
Beringia is a loosely defined region surrounding the Bering Strait, the Chukchi Sea, and the Bering Sea. It includes parts of Chukotka and Kamchatka in Russia , as well as Alaska in the United States . In historical contexts it also includes the Bering land bridge, an ancient land bridge roughly 1,600 kilometres (1,000 mi) wide (north to south) at its greatest extent, which connected Asia with North America at various times — all lying atop the existing North American plate, and east of the Siberian Chersky Range — during the Pleistocene ice ages. During ice ages, more water was stored as ice, the sea levels fell, and a land bridge was exposed.
East Beringia
In 2007, a study was undertaken on the skeletal material from 56 Pleistocene-period East Beringian wolves from permafrost deposits in Alaska. Uncalibrated radio carbon dating showed a continuous population from 45,500 years BP to 12,500 years BP, and one single wolf dated at 7,600 BP. This indicates that their population was in decline after 12,500 BP.[1] Megafauna was still available in this region until 10,500 BP, with the age of the more recent wolf specimen supported by the discovery of a remaining pocket of residual megafauna that still inhabited interior Alaska between 7,500–10,500 BP.[39]
The East Beringian wolf was identified as an ecomorph of the gray wolf (Canis lupis) with a skull morphology that was adapted for hunting and scavenging megafauna. None of the 16 mtDNA haplotypes recovered from a sample of 20 of the wolves was shared with any modern gray wolf, but similar haplotypes were found in Late Pleistocene Eurasian gray wolves. Six eastern-Beringian wolves had the same sequence found in two wolves from Ukraine dated 30,000 years BP and 28,000 years BP, and from Altai dated 33,000 years BP. Two eastern-Beringian wolves matched another haplotype with a wolf from the Czech Republic dated at 44,000 years BP. Its phylogeny indicates that, aside from the older-lineage Himalayan wolf and the Indian gray wolf, the Beringian wolf's unique haplotypes are basal to other gray wolves. Its genetic diversity was higher than that of its modern counterparts, implying that the wolf population of the Late Pleistocene was larger than the present population. Modern North American wolves are not their descendants, and this supports the existence of a separate origin for ancient and extant North American wolves.[1]
A more detailed analysis of the genetic material from three specimens were dated at 28,000 years BP, 21,000 years BP, and 20,800 years BP, respectively (with the samples deposited in GenBank with accession numbers KF661088, KF661089 and KF661090) and identified as Canis lupus.[40]
West Beringia
In 2009, a study was made on a skull fragment and right mandible of a wolf (Canis lupus) found near Lake Taimyr in the Taimyr Peninsula, Arctic Siberia, Russian Federation (the Lake Taimyr wolf). It is one of the northernmost records of Pleistocene carnivora in Eurasia. The skull was aged by radio carbon dating to 16,220 BP.[41]
The adult skull was small and assumed to be a female, as it did not differ in size to an extant female wolf skull from northern Siberia.[41] Another study of the Lake Taimyr wolf found that its comparatively small size and characters of the cheekteeth and skull raised the possibility that it might have been a domesticated or semi-domesticated animal.[42]
The increased skull width in comparison to extant wolves indicated pronounced development of the temporalis muscles. The specimens were compared to wolf (Canis lupus spelaeus) fossils found near Burnberg, Germany, and near the Paleolithic site of Kostenki 1 on the Don River near Voronezh, Russia. Both of the European fossil skulls demonstrated the same dentition as the fossil wolf from Taimyr. The skull and teeth arrangement suggest a considerable portion of carrion and bones in the diet. In the severe environmental conditions of the Late Pleistocene arctic zone of Eurasia, carrion had been one of the principal food sources for these animals. "Notably, the Pleistocene C. lupus from eastern Beringia, by the skull shape, tooth wear and isotopic data, is also reconstructed as a specialized hunter and scavenger of extinct North American megafauna."[41]
In 2019, the severed head of the world's first full-sized Pleistocene wolf was unearthed in the Abyisky district in the north of Yakutia. The wolf, whose rich mammoth-like fur and impressive fangs are still intact, was fully grown and aged from two to four years old when it died. ‘This is a unique discovery of the first ever remains of a fully grown Pleistocene wolf with its tissue preserved. We will be comparing it to modern-day wolves to understand how the species has evolved and to reconstruct its appearance,’ said an excited Albert Protopopov, from the Republic of Sakha Academy of Sciences.[43]
6.3. Europe
Canis lupus spelaeus
The European cave wolf (Canis lupus spelaeus) was first described by Georg August Goldfuss in 1823 based on a wolf pup skull found in the Zoolithen Cave located at Gailenreuth, Bavaria, Germany.[5] The wolf possibly belongs to a specialized Late Pleistocene wolf ecomorph. Its bone proportions are close to the Canadian Arctic-boreal mountain-adapted timber wolf and a little larger than those of the modern European wolf. It appears that in the early to middle Late Pleistocene this large wolf existed all over Europe, but was then replaced during the Last Glacial Maximum by a smaller wolf-type which then disappeared along with the reindeer fauna, finally replaced by the Holocene warm-period European wolf Canis lupus lupus. These wolves have not been well-studied, nor have they been well-defined by DNA.[44]
Canis lupus maximus
Canis lupus maximus (Boudadi-Maligne, 2012) was a subspecies larger than all other known fossil and extant wolves from Western Europe. The fossilized remains of this Late Pleistocene subspecies were found across a wide area of south-western France at: Jaurens cave, Nespouls, Corrèze dated 31,000 YBP; Maldidier cave, La Roque-Gageac, Dordogne dated 22,500 YBP; and Gral pit-fall, Sauliac-sur-Célé, Lot dated 16,000 YBP. The wolf's long bones are ten percent longer than those of extant European wolves and 20 percent longer than its probable ancestor, C.l. lunellensis. The teeth are robust, the posterior denticules on the lower premolars p2, p3, p4 and upper P2 and P3 are highly developed, and the diameter of the lower carnassial (m1) were larger than any known European wolf.[45]
Wolf body size in Europe has followed a steady increase from their first appearance up to the peak of the Last Glacial Maximum. The size of these wolves are thought to be an adaptation to a cold environment (Bergmann's rule) and plentiful game as their remains have been found in association with reindeer fossils.[45]
Italian wolves
A 2014 study found wolves in Late Pleistocene Italy were comparable in tooth morphology - and therefore in size - with C. l. maximus from France. These wolves were found near Avetrana, Taranto and near Buco del Frate, Brescia and Pocala cave in Friuli-Venezia Giulia.[46]
7. Relationship with the Domestic Dog

In 2013, a leading evolutionary biologist stated:[2]
We know also that there were distinct wolf populations existing ten of thousands of years ago. One such wolf, which we call the megafaunal wolf, preyed on large game such as horses, bison and perhaps very young mammoths. Isotope data show that they ate these species, and the dog may have been derived from a wolf similar to these ancient wolves in the late Pleistocene of Europe.—Robert K. Wayne, 2013
In 2015, a study looked at the mitogenome control region sequences of 13 ancient canid remains and one modern wolf from five sites across Arctic north-east Siberia. The 14 canids revealed nine haplotypes, three of which were on record and the others unique. Four of the Siberian canids dated 28,000 years before present (YBP), and one Canis variabilis dated 360,000 YBP. The phylogenetic relationship of the extracted sequences showed that the haplotype from specimen S805 (28,000 YBP) was one step away from another haplotype S902 (8,000 YBP) that represents the domestic dog and modern wolf lineages. Several ancient haplotypes were oriented around S805, including Canis c.f. variabilis (360,000 YBP), Belgium (36,000 YBP – the "Goyet dog") and Belgium (30,000 YBP), and Kostyonki, Voronezh Oblast, Russia (22,000 YBP). Given the position of the S805 haplotype, it may potentially represent a direct link from the putative progenitor (including Canis c.f. variabilis) to the domestic dog and modern wolf lineages.[47]
References
- Leonard, J. A.; Vilà, C; Fox-Dobbs, K; Koch, P. L.; Wayne, R. K.; Van Valkenburgh, B (2007). "Megafaunal extinctions and the disappearance of a specialized wolf ecomorph". Current Biology 17 (13): 1146–50. doi:10.1016/j.cub.2007.05.072. PMID 17583509. http://www.es.ucsc.edu/~pkoch/pdfs/Koch%20papers/2007/Leonard%20et%2007%20CurBio%2017-1146.pdf. Retrieved 2015-09-04.
- Wolpert, Stuart (November 14, 2013). "Dogs likely originated in Europe more than 18,000 years ago, UCLA biologists report". UCLA News Room. http://newsroom.ucla.edu/releases/dogs-likely-originated-in-europe-249325. Retrieved December 10, 2014.
- For a full set of supporting references refer to the note (a) in the phylotree at Evolution of the wolf
- Pilot, M. (2010). "Phylogeographic history of grey wolves in Europe". BMC Evolutionary Biology 10: 104. doi:10.1186/1471-2148-10-104. PMID 20409299. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2873414
- Goldfuß, G.A., 1823: Osteologische Beiträge zur Kenn- tniß verschiedener Säugethiere der Vorwelt. V. Ue - ber den Hölenwolf (Canis spelaeus).- Nov. Act. acad. Leopold., XI, 451-455. [Osteological contributions to different knowledge Beast of the ancients V. About the cave-wolf (Canis spelaeus).] https://books.google.com/books?id=7ub4f7z-IUYC&printsec=frontcover#v=onepage&q&f=false
- Leonard, Jennifer (2014). "Ecology drives evolution in grey wolves". Evolution Ecology Research 16: 461–473. http://www.evolutionary-ecology.com/open/v16n06/eear2913.pdf. Retrieved 2016-05-23.
- Musiani, Marco; Leonard, Jennifer A.; Cluff, H. Dean; Gates, C. Cormack; Mariani, Stefano; Paquet, Paul C.; Vilà, Carles; Wayne, Robert K. (2007). "Differentiation of tundra/taiga and boreal coniferous forest wolves: Genetics, coat colour and association with migratory caribou". Molecular Ecology 16 (19): 4149–70. doi:10.1111/j.1365-294X.2007.03458.x. PMID 17725575. https://semanticscholar.org/paper/2d88c88aa2a549dafd17f379ba471daeff8288ee.
- Carmichael, L. E.; Nagy, J. A.; Larter, N. C.; Strobeck, C. (2001). "Prey specialization may influence patterns of gene flow in wolves of the Canadian Northwest". Molecular Ecology 10 (12): 2787–98. doi:10.1046/j.0962-1083.2001.01408.x. PMID 11903892. https://dx.doi.org/10.1046%2Fj.0962-1083.2001.01408.x
- Carmichael, L.E., 2006. Ecological Genetics of Northern Wolves and Arctic Foxes. Ph.D. Dissertation. University of Alberta.
- Geffen, ELI; Anderson, Marti J.; Wayne, Robert K. (2004). "Climate and habitat barriers to dispersal in the highly mobile grey wolf". Molecular Ecology 13 (8): 2481–90. doi:10.1111/j.1365-294X.2004.02244.x. PMID 15245420. https://semanticscholar.org/paper/37558ff9144ec8bbfbb41bc820e14b482efd4d54.
- Pilot, Malgorzata; Jedrzejewski, Wlodzimierz; Branicki, Wojciech; Sidorovich, Vadim E.; Jedrzejewska, Bogumila; Stachura, Krystyna; Funk, Stephan M. (2006). "Ecological factors influence population genetic structure of European grey wolves". Molecular Ecology 15 (14): 4533–53. doi:10.1111/j.1365-294X.2006.03110.x. PMID 17107481. https://dx.doi.org/10.1111%2Fj.1365-294X.2006.03110.x
- Hofreiter, Michael; Barnes, Ian (2010). "Diversity lost: Are all Holarctic large mammal species just relict populations?". BMC Biology 8: 46. doi:10.1186/1741-7007-8-46. PMID 20409351. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2858106
- Flower, Lucy O.H.; Schreve, Danielle C. (2014). "An investigation of palaeodietary variability in European Pleistocene canids". Quaternary Science Reviews 96: 188–203. doi:10.1016/j.quascirev.2014.04.015. Bibcode: 2014QSRv...96..188F. https://dx.doi.org/10.1016%2Fj.quascirev.2014.04.015
- Perri, Angela (2016). "A wolf in dog's clothing: Initial dog domestication and Pleistocene wolf variation". Journal of Archaeological Science 68: 1–4. doi:10.1016/j.jas.2016.02.003. https://dx.doi.org/10.1016%2Fj.jas.2016.02.003
- Randi, Ettore (2011). "Genetics and conservation of wolves Canis lupus in Europe". Mammal Review 41 (2): 99–111. doi:10.1111/j.1365-2907.2010.00176.x. https://semanticscholar.org/paper/2a7ede0bf7457a1f37a6c6910ef547e0a9a7992c.
- Sansalone, Gabriele; Bertè, Davide Federico; Maiorino, Leonardo; Pandolfi, Luca (2015). "Evolutionary trends and stasis in carnassial teeth of European Pleistocene wolf Canis lupus (Mammalia, Canidae)". Quaternary Science Reviews 110: 36–48. doi:10.1016/j.quascirev.2014.12.009. https://dx.doi.org/10.1016%2Fj.quascirev.2014.12.009
- DeSantis, L.R.G. §, Schubert, B.W., *Schmitt-Linville, E., Ungar, P., *Donohue, S., *Haupt, R.J. In press. Dental microwear textures of carnivorans from the La Brea Tar Pits, California and potential extinction implications . Contributions in Science (a special volume entitled La Brea and Beyond: the Paleontology of Asphalt-Preserved Biotas, in commemoration of the 100th anniversary of the Natural History Museum of Los Angeles County's excavations at Rancho La Brea) http://www.tarpits.org/sites/default/files/pdfs/Rice%20et%20al%202015.%20LACM%20SS%2042.pdf#page=43
- Pavelková Řičánková, Věra; Robovský, Jan; Riegert, Jan (2014). "Ecological Structure of Recent and Last Glacial Mammalian Faunas in Northern Eurasia: The Case of Altai-Sayan Refugium". PLoS ONE 9 (1): e85056. doi:10.1371/journal.pone.0085056. PMID 24454791. Bibcode: 2014PLoSO...985056P. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=3890305
- Intergovernmental Panel on Climate Change (UN). "IPCC Fourth Assessment Report: Climate Change 2007 - Palaeoclimatic Perspective". The Nobel Foundation. https://www.ipcc.ch/publications_and_data/ar4/wg1/en/spmsspm-a-palaeoclimatic.html.
- Clark, P. U.; Dyke, A. S.; Shakun, J. D.; Carlson, A. E.; Clark, J.; Wohlfarth, B.; Mitrovica, J. X.; Hostetler, S. W. et al. (2009). "The Last Glacial Maximum". Science 325 (5941): 710–4. doi:10.1126/science.1172873. PMID 19661421. Bibcode: 2009Sci...325..710C. https://semanticscholar.org/paper/265b6f8e5e8afa1cc41e7a44ad7eb80aed7fa752.
- Elias, Scott A.; Short, Susan K.; Nelson, C. Hans; Birks, Hilary H. (1996). "Life and times of the Bering land bridge". Nature 382 (6586): 60–63. doi:10.1038/382060a0. Bibcode: 1996Natur.382...60E. https://dx.doi.org/10.1038%2F382060a0
- Jonathan Adams. "Europe during the last 150,000 years". Oak Ridge National Laboratory, Oak Ridge, USA. Archived from the original on 2005-11-26. https://web.archive.org/web/20051126084405/http://www.esd.ornl.gov/projects/qen/nercEUROPE.html.
- Stuart, Anthony John (1999). Late Pleistocene Megafaunal Extinctions, in Extinctions in Near Time. pp. 257–269. doi:10.1007/978-1-4757-5202-1_11. ISBN 978-1-4419-3315-7. https://dx.doi.org/10.1007%2F978-1-4757-5202-1_11
- Dale Guthrie, R. (2004). "Radiocarbon evidence of mid-Holocene mammoths stranded on an Alaskan Bering Sea island". Nature 429 (6993): 746–749. doi:10.1038/nature02612. PMID 15201907. Bibcode: 2004Natur.429..746D. https://dx.doi.org/10.1038%2Fnature02612
- Reumer, Jelle W. F.; Rook, Lorenzo; Van Der Borg, Klaas; Post, Klaas; Mol, Dick; De Vos, John (2003). "Late Pleistocene survival of the saber-toothed cat Homotheriumin northwestern Europe". Journal of Vertebrate Paleontology 23: 260–262. doi:10.1671/0272-4634(2003)23[260:LPSOTS2.0.CO;2]. https://dx.doi.org/10.1671%2F0272-4634%282003%2923%5B260%3ALPSOTS%5D2.0.CO%3B2
- Barnett, R.; Shapiro, B.; Barnes, I. A. N.; Ho, S. Y. W.; Burger, J.; Yamaguchi, N.; Higham, T. F. G.; Wheeler, H. T. et al. (2009). "Phylogeography of lions (Panthera leo ssp.) reveals three distinct taxa and a late Pleistocene reduction in genetic diversity". Molecular Ecology 18 (8): 1668–1677. doi:10.1111/j.1365-294X.2009.04134.x. PMID 19302360. https://semanticscholar.org/paper/086261772c124f43cb21755e5ef68becff2c5df5.
- Ghezzo, Elena; Rook, Lorenzo (2015). "The remarkable Panthera pardus (Felidae, Mammalia) record from Equi (Massa, Italy): Taphonomy, morphology, and paleoecology". Quaternary Science Reviews 110: 131–151. doi:10.1016/j.quascirev.2014.12.020. https://dx.doi.org/10.1016%2Fj.quascirev.2014.12.020
- Cooper, A.; Turney, C.; Hughen, K. A.; Brook, B. W.; McDonald, H. G.; Bradshaw, C. J. A. (2015). "Abrupt warming events drove Late Pleistocene Holarctic megafaunal turnover". Science 349 (6248): 602–6. doi:10.1126/science.aac4315. PMID 26250679. Bibcode: 2015Sci...349..602C. https://dx.doi.org/10.1126%2Fscience.aac4315
- White, T. D.; Asfaw, B.; Degusta, D.; Gilbert, H.; Richards, G. D.; Suwa, G.; Clark Howell, F. (2003). "Pleistocene Homo sapiens from Middle Awash, Ethiopia". Nature 423 (6941): 742–7. doi:10.1038/nature01669. PMID 12802332. Bibcode: 2003Natur.423..742W. https://dx.doi.org/10.1038%2Fnature01669
- "A Human Journey:Migration Routes". The genographic project. National Geographic Society. 2015. https://genographic.nationalgeographic.com/human-journey/. Retrieved 22 May 2017.
- Benazzi, S.; Douka, K.; Fornai, C.; Bauer, C. C.; Kullmer, O.; Svoboda, J. Í.; Pap, I.; Mallegni, F. et al. (2011). "Early dispersal of modern humans in Europe and implications for Neanderthal behaviour". Nature 479 (7374): 525–8. doi:10.1038/nature10617. PMID 22048311. Bibcode: 2011Natur.479..525B. https://dx.doi.org/10.1038%2Fnature10617
- Higham, T.; Compton, T.; Stringer, C.; Jacobi, R.; Shapiro, B.; Trinkaus, E.; Chandler, B.; Gröning, F. et al. (2011). "The earliest evidence for anatomically modern humans in northwestern Europe". Nature 479 (7374): 521–4. doi:10.1038/nature10484. PMID 22048314. Bibcode: 2011Natur.479..521H. https://dx.doi.org/10.1038%2Fnature10484
- Pavlov, Pavel; Svendsen, John Inge; Indrelid, Svein (2001). "Human presence in the European Arctic nearly 40,000 years ago". Nature 413 (6851): 64–7. doi:10.1038/35092552. PMID 11544525. Bibcode: 2001Natur.413...64P. https://dx.doi.org/10.1038%2F35092552
- "Mamontovaya Kurya:an enigmatic, nearly 40000 years old Paleolithic site in the Russian Arctic". https://notendur.hi.is/oi/AG-326%202006%20readings/Russian%20Arctic/Svendsen_MAMMOTH2003.pdf.
- Pitulko, V. V.; Tikhonov, A. N.; Pavlova, E. Y.; Nikolskiy, P. A.; Kuper, K. E.; Polozov, R. N. (2016). "Early human presence in the Arctic: Evidence from 45,000-year-old mammoth remains". Science 351 (6270): 260–3. doi:10.1126/science.aad0554. PMID 26816376. Bibcode: 2016Sci...351..260P. https://dx.doi.org/10.1126%2Fscience.aad0554
- Pitulko, V. V.; Nikolsky, P. A.; Girya, E. Y.; Basilyan, A. E.; Tumskoy, V. E.; Koulakov, S. A.; Astakhov, S. N.; Pavlova, E. Y. et al. (2004). "The Yana RHS Site: Humans in the Arctic Before the Last Glacial Maximum". Science 303 (5654): 52–6. doi:10.1126/science.1085219. PMID 14704419. Bibcode: 2004Sci...303...52P. https://dx.doi.org/10.1126%2Fscience.1085219
- Tamm, E.; Kivisild, T.; Reidla, M.; Metspalu, M.; Smith, D. G.; Mulligan, C. J.; Bravi, C. M.; Rickards, O. et al. (2007). Carter, Dee. ed. "Beringian Standstill and Spread of Native American Founders". PLoS ONE 2 (9): e829. doi:10.1371/journal.pone.0000829. PMID 17786201. Bibcode: 2007PLoSO...2..829T. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1952074
- Balter, M (4 July 2013). "Farming Was So Nice, It Was Invented at Least Twice". Science. http://news.sciencemag.org/archaeology/2013/07/farming-was-so-nice-it-was-invented-least-twice.
- Haile, J.; Froese, D. G.; MacPhee, R. D. E.; Roberts, R. G.; Arnold, L. J.; Reyes, A. V.; Rasmussen, M.; Nielsen, R. et al. (2009). "Ancient DNA reveals late survival of mammoth and horse in interior Alaska". Proceedings of the National Academy of Sciences 106 (52): 22352–7. doi:10.1073/pnas.0912510106. PMID 20018740. Bibcode: 2009PNAS..10622352H. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2795395
- Thalmann, O.; Shapiro, B.; Cui, P.; Schuenemann, V. J.; Sawyer, S. K.; Greenfield, D. L.; Germonpré, M. B.; Sablin, M. V. et al. (2013). "Complete Mitochondrial Genomes of Ancient Canids Suggest a European Origin of Domestic Dogs". Science 342 (6160): 871–74. doi:10.1126/science.1243650. PMID 24233726. Bibcode: 2013Sci...342..871T. https://semanticscholar.org/paper/29804bbd577d3f792d85550627491f8ad8dad244. refer Supplementary material Page 27 Table S1
- Baryshnikov, Gennady F.; Mol, Dick; Tikhonov, Alexei N (2009). "Finding of the Late Pleistocene carnivores in Taimyr Peninsula (Russia, Siberia) with paleoecological context". Russian Journal of Theriology 8 (2): 107–113. doi:10.15298/rusjtheriol.08.2.04. http://zmmu.msu.ru/rjt/articles/article.php?volume=8&issue=2&pages=107-113. Retrieved December 23, 2014.
- MacPhee, R. D. E.; Tikhonov, A. N.; Mol, D.; De Marliave, C.; Van Der Plicht, H.; Greenwood, A. D.; Flemming, C.; Agenbroad, L. (2002). "Radiocarbon Chronologies and Extinction Dynamics of the Late Quaternary Mammalian Megafauna of the Taimyr Peninsula, Russian Federation". Journal of Archaeological Science 29 (9): 1017–1042. doi:10.1006/jasc.2001.0802. https://pure.rug.nl/ws/files/9931204/2002JArchaeolSciMacPhee.pdf. Retrieved 2019-07-21. quoted from page 1033
- Still snarling after 40,000 years, a giant Pleistocene wolf discovered in Yakutia (Note: until tooth measurements are published, there is no evidence that this wolf was megafaunally adapted) https://siberiantimes.com/science/casestudy/news/still-snarling-after-40000-years-a-giant-pleistocene-wolf-discovered-in-yakutia/
- Diedrich, C. G. (2013). "Extinctions of Late Ice Age Cave Bears as a Result of Climate/Habitat Change and Large Carnivore Lion/Hyena/Wolf Predation Stress in Europe". ISRN Zoology 2013: 1–25. doi:10.1155/2013/138319. https://dx.doi.org/10.1155%2F2013%2F138319
- Boudadi-Maligne, Myriam (2012). "Une nouvelle sous-espèce de loup (Canis lupus maximus nov. Subsp.) dans le Pléistocène supérieur d'Europe occidentale [A new subspecies of wolf (Canis lupus maximus nov. subsp.) from the upper Pleistocene of Western Europe]". Comptes Rendus Palevol 11 (7): 475. doi:10.1016/j.crpv.2012.04.003. https://dx.doi.org/10.1016%2Fj.crpv.2012.04.003
- Berte, E.; Pandolfi, L. (2014). "Canis lupus (Mammalia, Canidae) from the Late Pleistocene deposit of Avetrana (Taranto, Southern Italy)". Rivista Italiana di Paleontoligia e Stratigrafia 120 (3): 367–379.
- Lee, E. (2015). "Ancient DNA analysis of the oldest canid species from the Siberian Arctic and genetic contribution to the domestic dog". PLoS ONE 10 (5): e0125759. doi:10.1371/journal.pone.0125759. PMID 26018528. Bibcode: 2015PLoSO..1025759L. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=4446326


