Your browser does not fully support modern features. Please upgrade for a smoother experience.
Submitted Successfully!
 +1 credit
+1 credit
Thank you for your contribution! You can also upload a video entry or images related to this topic.
For video creation, please contact our Academic Video Service.
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Waqar Shafqat | -- | 2711 | 2022-11-21 22:32:58 | | | |
| 2 | Beatrix Zheng | + 1 word(s) | 2712 | 2022-11-22 03:13:19 | | | | |
| 3 | Beatrix Zheng | Meta information modification | 2712 | 2022-11-22 03:15:01 | | |
Video Upload Options
We provide professional Academic Video Service to translate complex research into visually appealing presentations. Would you like to try it?
Cite
If you have any further questions, please contact Encyclopedia Editorial Office.
Khalid, M.F.; Khan, R.I.; Jawaid, M.Z.; Shafqat, W.; Hussain, S.; Ahmed, T.; Rizwan, M.; Ercisli, S.; Pop, O.L.; Marc, R.A. Impact of Nanoparticles on Plants under Drought Stress. Encyclopedia. Available online: https://encyclopedia.pub/entry/35604 (accessed on 24 July 2026).
Khalid MF, Khan RI, Jawaid MZ, Shafqat W, Hussain S, Ahmed T, et al. Impact of Nanoparticles on Plants under Drought Stress. Encyclopedia. Available at: https://encyclopedia.pub/entry/35604. Accessed July 24, 2026.
Khalid, Muhammad Fasih, Rashid Iqbal Khan, Muhammad Zaid Jawaid, Waqar Shafqat, Sajjad Hussain, Talaat Ahmed, Muhammad Rizwan, Sezai Ercisli, Oana Lelia Pop, Romina Alina Marc. "Impact of Nanoparticles on Plants under Drought Stress" Encyclopedia, https://encyclopedia.pub/entry/35604 (accessed July 24, 2026).
Khalid, M.F., Khan, R.I., Jawaid, M.Z., Shafqat, W., Hussain, S., Ahmed, T., Rizwan, M., Ercisli, S., Pop, O.L., & Marc, R.A. (2022, November 21). Impact of Nanoparticles on Plants under Drought Stress. In Encyclopedia. https://encyclopedia.pub/entry/35604
Khalid, Muhammad Fasih, et al. "Impact of Nanoparticles on Plants under Drought Stress." Encyclopedia. Web. 21 November, 2022.
Copy Citation
Drought is a chronic abiotic stress affecting crop growth and development, accounting for approximately 70% of the potential loss of global crop yield and productivity. Drought hinders agriculture and forestry worldwide, due to very little rainfall or significant differences in moisture. The current trends of global warming are causing a major impact on the moisture levels of the soil and the environment, and are increasing the intensity of droughts. Plants are subjected to various stresses during their growth, and the morphology of plants is affected at all stages of development due to drought stress, with productivity losses expected to reach 30% globally by 2025.
nanoparticles
stress tolerance
physiology
molecular
drought
1. Introduction
A variety of factors influence agricultural productivity, including the climate. Agriculture is fundamental to human welfare, and many organizations and others are concerned about the effects of climate change on agriculture. As a result of increasing annual temperatures, changing patterns of rainfall, floods, and dwindling water reserves, major agriculture crops are affected by climate change. The agricultural sector provides income and employment to almost half of the labor force and supplies raw materials to industry in developing and less developed countries. Global hunger and food insecurity are continuously increasing due to the phenomenal increase in global population and stagnant agricultural performance [1]. Climate change causes many biotic and abiotic stresses to plants which affect plant growth and cause declines in yield [2]. Different strategies have been adopted to overcome these negative effects of climate change, i.e., the use of tolerant genotypes, application of different plant growth regulators, and the use of organic fertilizers. Currently, nanotechnology is substantially contributing to this sector. Nanotechnology studies the various structures of matter on the scale of a billionth of a meter. A nanoparticle (NP) is a small molecular aggregate with an interfacial layer surrounding a diameter of 1 to 100 nanometers. Several critical properties of matter are fundamentally impacted by this interfacial layer at the nanoscale [3][4]. As a result of their small size, NPs have some unusual properties compared with bulk materials. Nanoparticles refer to organic materials rather than individual molecules. The fact that NPs link bulk materials to atomic or molecular structures cause them to be of high scientific interest. The various NPs used for the treatment of plants to overcome environmental challenges are: titanium dioxide (TiO2), zinc (Zn), zinc oxide (ZnO), cesium (Ce), cobalt (Co), copper (Cu), copper oxide (CuO), selenium (Se) NPs, silver (Ag), silicon (Si), silicon oxide (SiO2), iron oxide (FeO), calcium (CaCO3), magnesium (Mg), magnesium oxide (MgO), manganese (Mn), and molybdenum (Mo) NPs; and aluminium oxide (AlO4) and carbon nanotubes (CNTs).
To cope with environmental stress, plants have developed a wide range of efficient and comprehensive molecular programs to rapidly sense stressors and adapt accordingly [5]. Plants can enhance this response through the interaction of NPs with plants. Nanotechnology promises to increase crop yield by improving plant tolerance mechanisms under abiotic stress conditions [6]. Several studies have shown that NPs play a vital role in improving the tolerance of plants to abiotic stresses by modulating various physiological, biochemical, and molecular processes (Figure 1).
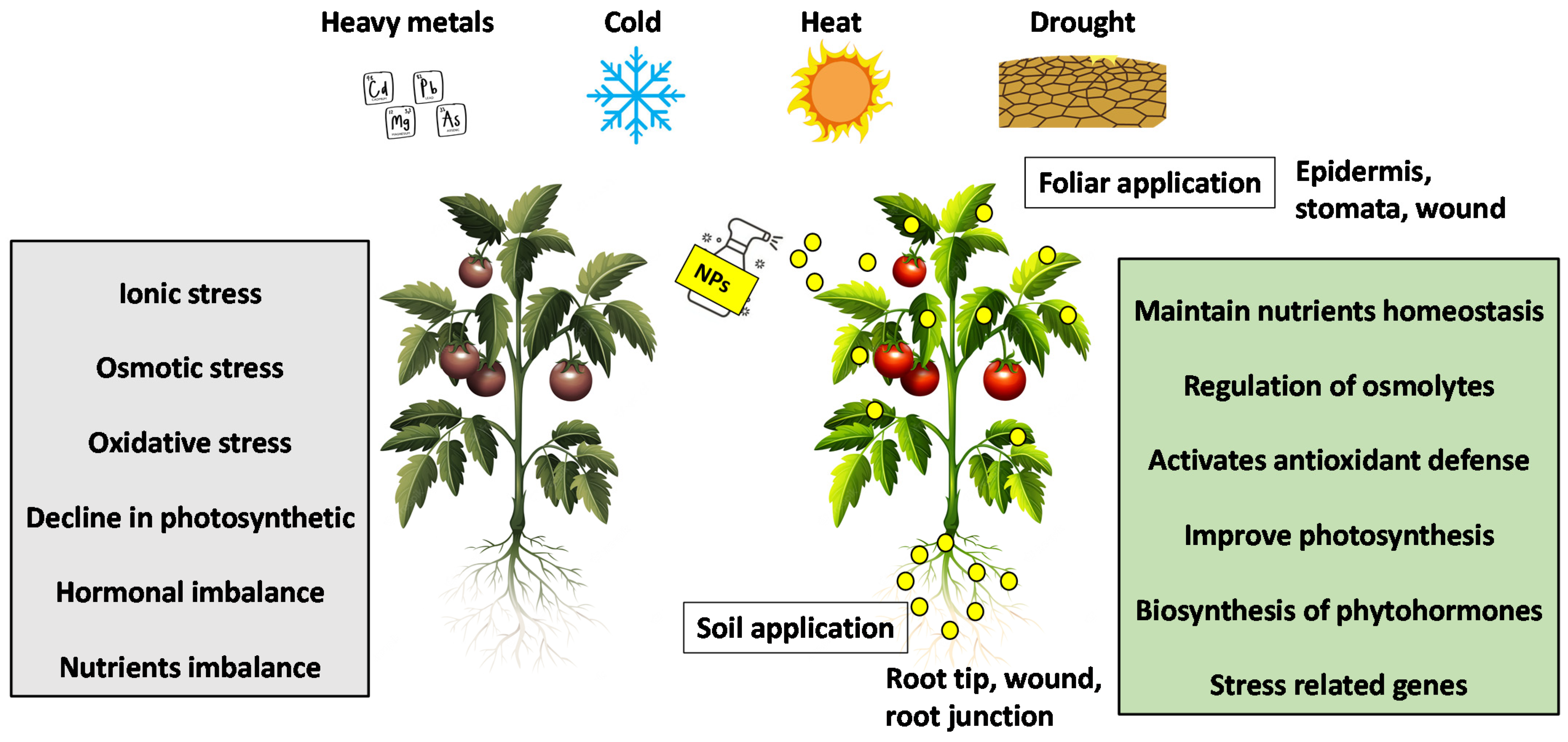
Figure 1. Mechanisms of NPs mitigate abiotic stresses in plants.
Crop growth and improvement can effectively be achieved in modern agriculture through nanotechnology. NPs can be used in the agricultural sector as nano-agrochemicals (nanobiocomposites, nanopesticides, nanofertilizers), agri-food production, nanobiosensors, agri-environment, organic agriculture, postharvest management, and plant genetic progress by NP-mediated gene transfer [7][8]. In recent years, the reliance on nanotechnology in different industries has been increasing due to its copious potential, sustainable, eco-friendly, and cost-effective applications. The use of nanopesticides and nanofertilizers has enhanced agricultural productivity, for example, urea-doped calcium phosphate nanofertilizers have helped commercial crops to obtain efficient nutrients from the soil, specifically urea; helped maintain crop growth and productivity; and helped to achieve sustainable agriculture [9][10][11]. Madusanka et al. [12] observed the slow release of nitrogen by using a urea-hydroxyapatite-montmorillonite nanohybrid composite. The use of hydroxyapatite nanoparticles significantly influenced the crop yield and germination attributes of tomato plants [13]. The range of applications of nanotechnology in the remediation of soil and water has increased food quality and production. Moreover, with nanotechnology being eco-friendly, its use has a significant benefit in reducing the harmful effects of chemicals used on crops, and the effects caused by agriculture on the environment [14]. NPs have been effective on seed and plant metabolisms by enhancing growth. The advantageous characteristics of NPs being small allows them to cross biological barriers in plants more efficiently and remediate plant stresses, such as salt stress and heat stress, and stress caused by heavy metals [15].
2. Drought Stress
Severe droughts are a major problem for agriculture in a changing climate, as water scarcity is predicted to become more common. Drought refers to the conditions where a plant’s water demand cannot be fully met, such as where the transpiration water level of the plant exceeds the water absorbed by the root system, insufficient precipitation, a drop in the groundwater level, or water retention by soil particles [16]. Plants reduce water loss through adjustments in morphological anatomy, physiology and biochemistry to maintain their water status as a result of drought [17][18]. Drought stress leads to a reduction in cell enlargement as compared with affecting cell division. It affects plant growth by altering the functioning of various physiological and biochemical processes, i.e., photosynthesis, respiration, enzymatic activity, and nutrient metabolism [19]. The response of plants to drought stress varies at different tissue levels, depending on the intensity and duration of the stress, as well as plant species and growth stage. Understanding how plants respond to drought is very important and an essential part of improving the tolerance of crops to stress.
Different molecular, biochemical, physiological, morphological and ecological traits and processes are disrupted under drought stress conditions [20][21]. A deficit of water has adverse effects on plant yield and quality. Growth stage, age, plant species, drought severity, and duration are key factors affecting plant response to drought [22]. Plants die off under prolonged drought conditions [23]. Water scarcity in plants increases the concentration of the solute in the cytosol and extracellular matrices as a result of the reduction in plant cells’ water potential and turgor, which leads to growth inhibition and reproductive failure. Wilting is caused by the accumulation of abscisic acid and compatible osmolytes [24][25]. Adverse influences are aggravated due to the overproduction of ROS and radical scavenging compounds such as ascorbate and glutathione [26][27]. Water stress in plants due to drought affects the stomatal functions and limits the gaseous exchange, decreasing the rate of transpiration and carbon assimilation [28]. In turn, the mechanisms of resistance of plants to drought vary. Therefore, plants can reduce resource utilization and regulate growth in response to adverse environmental conditions [29]. Signal transduction, a network at the molecular level, enhances these responses to drought stress [30]. Plant stomatal regulation by enhancing ion transport, transcription factor activity, and ABA signal transduction is also involved in the molecular mechanism of plant response [31]. In some changing environments, there is a need to enhance the resistance of plants against drought. To improve water use efficiency when the physical fitness of roots and leaves is insufficient to cope with certain drought molecular signals, plant enhancement may be conducted by including genes encoding regular proteins and signals by crosstalk, expressing many other genes according to different regulatory mechanisms [32]. To achieve future food demands, further advancement is required in enhancing drought tolerance in plants, and the adoption of economical and beneficial agricultural practices will be critical [33].
2.1. NPs Mitigate Drought Stress in Plants
NPs are known by their specific shape, tunable pore size, and high reactivity with enhanced surface area [34]. NPs are considered an effective and promising tool for regulating crop yield and overcoming current and future limitations of agricultural production by increasing the tolerance mechanisms in plants under abiotic stress conditions. The mitigating effect of NPs on drought stress is caused by inducing physiological and biochemical regulation, and regulating the expression of genes relating to drought response/tolerance. NPs enhance the photosynthetic activity of drought-induced plants, whereby the improvement of root growth, upregulation of aquaporins, altered intracellular water metabolism, accumulation of compatible solutes, and ionic homeostasis are the main mechanisms by which NPs alleviate osmotic stress caused by water deficiency. NPs reduce leaf water loss caused by the accumulation of ABA through stomatal closure, and ameliorate oxidative stress damage by reducing reactive oxygen species and activating antioxidant defense systems.
2.1.1. Physiological and Biochemical Aspect
Nanotechnology has the capability to enhance plant photosynthesis efficiency by altering the enzymatic activity involved in the C3 cycle, along with regulating photosynthetic pigments responsible for plant growth [35]. NPs have positive effects on plant germination and growth, however, their efficacy varies with their concentration and host plant. In sorghum plants under drought conditions, foliar spraying of nanowax increased seed yield in plants in comparison with spraying with water. TiO2 NPs have many strong effects on the morphological, biochemical, and physiological properties of crops [36]. During the growth phase of cucumber plants, exogenous application of NPs promoted rubisco activase activity, chlorophyll formation, and photosynthetic rate, which led to an increase in plant dry mass [37]. It was further noted that foliar application of NPs could increase the seed yield of soybean, due to enhanced photosynthesis [38].
The impact of nano-TiO2 varies with respect to changing environmental conditions, plant species, and different application doses. In this context, Mohammadi et al. [39] investigated the effects of nano-TiO2 concentrations on the biochemical and morphophysiological properties of dragonhead plants. The TiO2 increases the growth and essential oil in plants under water deficit stress. A formulation of nano-sized ZnO and CuO was used as a fertilizer. The results showed that at different NP doses, root growth was reduced, while contrarily, at other levels, Zn NPs expanded lateral root formation whereas Cu NPs induced proliferation and elongation of root hairs close to the roots of wheat seedlings under simulated drought stress [40]. These responses typically occur when the roots are colonized by a beneficial bacteria isolated from wheat roots grown in calcareous soils under dryland farming conditions.
It has been observed that ZnO and CuO NPs exhibited protection against drought stress in different plants [40]. This protection may be induced by the enhanced generation of lateral root hairs which resulted in proper water absorption. Enhanced cell wall lignification in mustard and Arabidopsis under CuO may alter water flow, thereby limiting cell wall elongation. The response of plants to drought stress is an increase in lignification. The disruption of water flow occurs due to the binding of copper ions to the pectin of the cell wall [41]. Some notable results were found in some studies, such as increased seed germination and antioxidant content after barley, soybean, and maize were treated with carbon nanotubes (CNTs) [42]. CNTs can induce root and shoot growth in wheat plants. Various major efforts have been conducted over the past few decades to reduce the effects of drought stress on plant quality and productivity. The researchers further suggest that fullerenol (FNPs) NPs with molecular formula C60(OH)24 may help alleviate the effects of drought stress and provide additional water supply between plant cells. Precisely, nanofullerenols (FNPs) can enter the root and leaf tissues of plants, where they can bind water molecules in various parts of the cell. This water absorbing FNP activity further suggests that FNPs may be useful for plants [43][44]. The results of this study by Borišev et al. [43] further demonstrated that foliar application of nanofullerenol could alter intracellular water metabolism in drought-stressed plants. Under drought stress, the content of the permeate product proline in plant roots and leaves was significantly increased. These results further suggest that FNPs could also function as a binder for intracellular water, thereby generating additional water reserves, and allowing them to adapt to drought stress. Ag NPs are the most used NPs in research experiments [45].
In plants, NPs target the cellular organelles and release various contents [8], thus modulating the activity of antioxidants enzymes, i.e., SOD, CAT, and POD [46]. This effect was exhibited by incremented SOD activity in plants under TiO2 NP application [47]. In agriculture, certain elements, along with oxides as NPs, have been used for incremental resistance against drought stress. Si NPs have been used extensively for ameliorating the negative impacts of various abiotic stresses including drought [48]. The improvement in growth, physio- and biochemical characteristics has been observed upon treatment with silica and ZnO NPs on different crops [34]. Similarly, Si NPs ameliorated drought stress on wheat plants [49]. Similarly, ZnO NPs reduced the negative impact of salinity and drought stress on plants [50]. It has been observed that excessive NP application led to a generation of oxidative stress, i.e., leading to cell cycle arrest, programmed cell death, protein regulation, and induction of antioxidant enzymes [51], whereas NP-treated plants exhibited significant reductions in MDA levels along with free radicals, i.e., H2O2 and O2−, under drought conditions. However, it was also observed that TiO2 application enhanced antioxidant enzyme activities, i.e., POD and CAT, whereas MDA levels were reduced due to the induction of the plant’s antioxidant system [26].
Under drought stress, the level of anthocyanin in plants exposed to CuO NPs continued to increase, and the level of proline was also shown to increase under drought stress. Wheat roots treated with CuO-treated NPs exhibited a greater accumulation of free radicals, consistent with plants responding to the challenge of NP-induced ROS bursts. Elevated ROS levels, further suggesting that drought stress triggers a consequence of elevated ABA, may lead to transcriptional changes that lead to tolerance. The amplification of various antioxidant enzymes (GR, SOD, GPX, APX, and CAT) in plants suggested that foliar application of fullerenol (FNPs) NPs with molecular formula C60(OH)24 might have some valuable effects on mitigating the oxidative effect of drought stress, which further depends on the concentration of NPs applied [42]. The exact mode of action and physiological mechanism of FNPs on plants needs to be further studied.
2.1.2. Molecular Aspect
Transcriptomic and proteomic approaches have deeply investigated the effects of NPs on different plant species at the molecular level. Morphological and physiological effects have been reported to largely depend on the dose used, as well as the type, size and shape of NPs [52][53]. Expression of the P5CS gene leads to increased plant tolerance to different environmental stress conditions, including biotic and abiotic stresses, since this gene encodes proline biosynthesis. MAPK2, a member of the MAP kinase gene family, plays an extremely important role in regulating phytohormones and antioxidant protection mechanisms in response to different stress environments [53] in combination with Ca21 and ROS. AREB/ABF are transcriptional regulators necessary for the regulation of the AREB gene encoding abscisic acid, and are critical in stimulating resistance to stressful environments such as drought and salt stress [53][54]. Downregulation of the ZFHD gene reduces the negative effects of salt and drought stress and is controlled by the abscisic acid biosynthesis pathway. On the other hand, downregulation of the TAS14 gene reduces osmotic pressure and enhances solute aggregation, including K1 and sugars, making plant species more resistant to drought and salt stress [53]. Application of Ag NPs (5 and 10 mg/L) to rape plants modulated the metabolic pathways of glucosinolate and phenolic related genes, which are also associated with biotic and abiotic stresses, and inhibited carotenoid genes [55]. Downregulation of the ZFHD gene reduces the negative effects of salt and drought stress and is controlled by the abscisic acid biosynthesis pathway. The use of Ag and Ag1 NPs on Arabidopsis plants resulted in overexpression of oxidative stress and metal response-related genes, and downregulation of ethylene and auxin-related genes [53]. Three of these genes overexpressed by Ag NPs are involved in the biosynthesis of thalianol, which is thought to contribute to a plant’s antioxidant protection mechanism. The response of different NPs against drought stress conditions is summarized in Table 1.
Table 1. Impact of NPs on plants under drought stress.
| NPs | Plant | Effect | Reference |
|---|---|---|---|
| ZnO | Triticum aestivum L. | Enhanced plant growth and mineral content in grains. | [56] |
| B NPs, SiO2 NPs and Zn NPs | Triticum aestivum L. | Enhanced protein contents and mitigates drought stress. | [57] |
| ZnO NPs | Zea mays L. | Enhanced yield and ameliorated antioxidative behavior. | [58] |
| Nano-Cu NPs | Zea mays L. | Upgraded the protective mechanism of maize under drought conditions. | [59] |
| Nano-Si NPs | Tanacetum parthenium L. | Improved water and phosphorus absorbing capabilities and general growth. | [60] |
| ZnO NPs | Solanum lycopersicum | Enhanced ascorbic acid and free phenols conc. along with the ameliorated activity of antioxidative enzymes. | [61] |
| Cu, Fe and Zn NPs | Glycine max (L.) Merrill | Upregulated expression of drought-sensitive genes. | [38] |
References
- Arora, N.K. Impact of climate change on agriculture production and its sustainable solutions. Environ. Sustain. 2019, 2, 95–96.
- Khalid, M.F.; Hussain, S.; Ahmad, S.; Ejaz, S.; Zakir, I.; Ali, M.A.; Ahmed, N.; Anjum, M.A. Impacts of Abiotic Stresses on Growth and Development of Plants. In Plant Tolerance to Environmental Stress; CRC Press: Boca Raton, FL, USA, 2019; pp. 1–8.
- Jeevanandam, J.; Barhoum, A.; Chan, Y.S.; Dufresne, A.; Danquah, M.K. Review on nanoparticles and nanostructured materials: History, sources, toxicity and regulations. Beilstein J. Nanotechnol. 2018, 9, 1050–1074.
- Baby, A.; Nazeerudeen, S.; Ranganath, S.; Samuel, R.S. Toxicological impacts of nanoparticles. In Emerging Trends of Nanotechnology in Environment and Sustainability; Springer: Cham, Switzerland, 2018; pp. 77–78.
- Van Wallendael, A.; Soltani, A.; Emery, N.C.; Peixoto, M.M.; Olsen, J.; Lowry, D.B. A Molecular View of Plant Local Adaptation: Incorporating Stress-Response Networks. Annu. Rev. Plant Biol. 2019, 70, 559–583.
- Arif, Y.; Singh, P.; Siddiqui, H.; Bajguz, A.; Hayat, S. Salinity induced physiological and biochemical changes in plants: An omic approach towards salt stress tolerance. Plant Physiol. Biochem. 2020, 156, 64–77.
- Heikal, Y.M.; El-Esawi, M.A.; El-Ballat, E.M.; Abdel-Aziz, H.M.M. Applications of nanoparticles for mitigating salinity and drought stress in plants: An overview on the physiological, biochemical and molecular genetic aspects. N. Z. J. Crop Hortic. Sci. 2022.
- Cunningham, F.J.; Goh, N.S.; Demirer, G.S.; Matos, J.L.; Landry, M.P. Nanoparticle-Mediated Delivery towards Advancing Plant Genetic Engineering. Trends Biotechnol. 2018, 36, 882–897.
- Feil, S.B.; Rodegher, G.; Gaiotti, F.; Alzate, M.Y.Z.; Carmona, F.J.; Masciocchi, N.; Cesco, S.; Pii, Y. Physiological and Molecular Investigation of Urea Uptake Dynamics in Cucumis sativus L. Plants Fertilized with Urea-Doped Amorphous Calcium Phosphate Nanoparticles. Front. Plant Sci. 2021, 12, 745581.
- Carmona, F.J.; Sasso, G.D.; Ramírez-Rodríguez, G.B.; Pii, Y.; Delgado-López, J.M.; Guagliardi, A.; Masciocchi, N. Urea-functionalized amorphous calcium phosphate nanofertilizers: Optimizing the synthetic strategy towards environmental sustainability and manufacturing costs. Sci. Rep. 2021, 11, 3419.
- Gaiotti, F.; Lucchetta, M.; Rodegher, G.; Lorenzoni, D.; Longo, E.; Boselli, E.; Cesco, S.; Belfiore, N.; Lovat, L.; Delgado-López, J.; et al. Urea-Doped Calcium Phosphate Nanoparticles as Sustainable Nitrogen Nanofertilizers for Viticulture: Implications on Yield and Quality of Pinot Gris Grapevines. Agronomy 2021, 11, 1026.
- Madusanka, N.; Sandaruwan, C.; Kottegoda, N.; Sirisena, D.; Munaweera, I.; De Alwis, A.; Karunaratne, V.; Amaratunga, G.A. Urea–hydroxyapatite-montmorillonite nanohybrid composites as slow release nitrogen compositions. Appl. Clay Sci. 2017, 150, 303–308.
- Marchiol, L.; Filippi, A.; Adamiano, A.; Degli Esposti, L.; Iafisco, M.; Mattiello, A.; Petrussa, E.; Braidot, E. Influence of Hydroxyapatite Nanoparticles on Germination and Plant Metabolism of Tomato (Solanum lycopersicum L.): Preliminary Evidence. Agronomy 2019, 9, 161.
- Ahmad, B.; Zaid, A.; Zulfiqar, F.; Bovand, F.; Dar, T.A. Nanotechnology: A novel and sustainable approach towards heavy metal stress alleviation in plants. Nanotechnol. Environ. Eng. 2022.
- Pereira, A.D.E.S.; Oliveira, H.C.; Fraceto, L.F.; Santaella, C. Nanotechnology Potential in Seed Priming for Sustainable Agriculture. Nanomaterials 2021, 11, 267.
- Salehi-Lisar, S.Y.; Bakhshayeshan-Agdam, H. Drought stress in plants: Causes, consequences, and tolerance. In Drought Stress Tolerance in Plants; Springer: Cham, Switzerland, 2016.
- Khalid, M.F.; Hussain, S.; Anjum, M.A.; Morillon, R.; Ahmad, S.; Ejaz, S.; Hussain, M.; Jaafar, H.Z.E.; Alrashood, S.T.; Ormenisan, A.N. Physiological and biochemical responses of Kinnow mandarin grafted on diploid and tetraploid Volkamer lemon rootstocks under different water-deficit regimes. PLoS ONE 2021, 16, e0247558.
- Chaudhry, S.; Sidhu, G.P.S. Climate change regulated abiotic stress mechanisms in plants: A comprehensive review. Plant Cell Rep. 2021, 41, 1–31.
- Jeyasri, R.; Muthuramalingam, P.; Satish, L.; Pandian, S.; Chen, J.-T.; Ahmar, S.; Wang, X.; Mora-Poblete, F.; Ramesh, M. An Overview of Abiotic Stress in Cereal Crops: Negative Impacts, Regulation, Biotechnology and Integrated Omics. Plants 2021, 10, 1472.
- Khalid, M.F.; Vincent, C.; Morillon, R.; Anjum, M.A.; Ahmad, S.; Hussain, S. Different strategies lead to a common outcome: Different water-deficit scenarios highlight physiological and biochemical strategies of water-deficit tolerance in diploid versus tetraploid Volkamer lemon. Tree Physiol. 2021, 41, 2359–2374.
- Ortiz, N.; Armada, E.; Duque, E.; Roldán, A.; Azcón, R. Contribution of arbuscular mycorrhizal fungi and/or bacteria to enhancing plant drought tolerance under natural soil conditions: Effectiveness of autochthonous or allochthonous strains. J. Plant Physiol. 2015, 174, 87–96.
- Gray, S.B.; Brady, S.M. Plant developmental responses to climate change. Dev. Biol. 2016, 419, 64–77.
- Takahashi, F.; Kuromori, T.; Sato, H.; Shinozaki, K. Regulatory gene networks in drought stress responses and resistance in plants. In Survival Strategies in Extreme Cold and Desiccation; Springer: Singapore, 2018; pp. 189–214.
- Ahmad, J.; Bashir, H.; Bagheri, R.; Baig, A.; Al-Huqail, A.; Ibrahim, M.M.; Qureshi, M.I. Drought and salinity induced changes in ecophysiology and proteomic profile of Parthenium hysterophorus. PLoS ONE 2017, 12, e0185118.
- Hussain, S.; Hussain, S.; Qadir, T.; Khaliq, A.; Ashraf, U.; Parveen, A.; Rafiq, M. Drought stress in plants: An overview on implications, tolerance mechanisms and agronomic mitigation strategies. Plant Sci. Today 2019, 6, 389–402.
- Ahmad, J.; Ali, A.A.; Baig, M.A.; Iqbal, M.; Haq, I.; Qureshi, M.I. Role of phytochelatins in cadmium stress tolerance in plants. In Cadmium Toxicity and Tolerance in Plants; Academic Press: Cambridge, UK, 2019; pp. 185–212.
- Hussain, A.; Ali, S.; Rizwan, M.; Rehman, M.Z.U.; Qayyum, M.F.; Wang, H.; Rinklebe, J. Responses of wheat (Triticum aestivum) plants grown in a Cd contaminated soil to the application of iron oxide nanoparticles. Ecotoxicol. Environ. Saf. 2019, 173, 156–164.
- Schulze, E.-D.; Beck, E.; Buchmann, N.; Clemens, S.; Müller-Hohenstein, K.; Scherer-Lorenzen, M. Water Deficiency (Drought). In Plant Ecology; Springer: Berlin, Heidelberg, 2019; pp. 165–202.
- Bielach, A.; Hrtyan, M.; Tognetti, V.B. Plants under Stress: Involvement of Auxin and Cytokinin. Int. J. Mol. Sci. 2017, 18, 1427.
- Zandalinas, S.I.; Fritschi, F.B.; Mittler, R. Signal transduction networks during stress combination. J. Exp. Bot. 2020, 71, 1734–1741.
- Prakash, V.; Singh, V.P.; Tripathi, D.K.; Sharma, S.; Corpas, F.J. Crosstalk between nitric oxide (NO) and abscisic acid (ABA) signalling molecules in higher plants. Environ. Exp. Bot. 2019, 161, 41–49.
- Shahid, M.J.; Ali, S.; Shabir, G.; Siddique, M.; Rizwan, M.; Seleiman, M.F.; Afzal, M. Comparing the performance of four macrophytes in bacterial assisted floating treatment wetlands for the removal of trace metals (Fe, Mn, Ni, Pb, and Cr) from polluted river water. Chemosphere 2020, 243, 125353.
- Diatta, A.A.; Thomason, W.E.; Abaye, O.; Thompson, T.L.; Battaglia, M.L.; Vaughan, L.J.; Lo, M.; Filho, J.F.D.C.L. Assessment of Nitrogen Fixation by Mungbean Genotypes in Different Soil Textures Using 15N Natural Abundance Method. J. Soil Sci. Plant Nutr. 2020, 20, 2230–2240.
- Seleiman, M.F.; Almutairi, K.F.; Alotaibi, M.; Shami, A.; Alhammad, B.A.; Battaglia, M.L. Nano-Fertilization as an Emerging Fertilization Technique: Why Can Modern Agriculture Benefit from Its Use? Plants 2020, 10, 2.
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and challenges for nanotechnology in the agritech revolution. Nat. Nanotechnol. 2019, 14, 517–522.
- Djanaguiraman, M.; Belliraj, N.; Bossmann, S.H.; Prasad, P.V. High-temperature stress alleviation by selenium nanoparticle treatment in grain sorghum. ACS Omega 2018, 3, 2479–2491.
- Ghani, M.I.; Saleem, S.; Rather, S.A.; Rehmani, M.S.; Alamri, S.; Rajput, V.D.; Kalaji, H.M.; Saleem, N.; Sial, T.A.; Liu, M. Foliar application of zinc oxide nanoparticles: An effective strategy to mitigate drought stress in cucumber seedling by modulating antioxidant defense system and osmolytes accumulation. Chemosphere 2022, 289, 133202.
- Linh, T.M.; Mai, N.C.; Hoe, P.T.; Lien, L.Q.; Ban, N.K.; Hien, L.T.T.; Chau, N.H.; Van, N.T. Metal-Based Nanoparticles Enhance Drought Tolerance in Soybean. J. Nanomater. 2020, 2020, 4056563.
- Mohammadi, H.; Esmailpour, M.; Gheranpaye, A. Effects of TiO2 nanoparticles and water deficit stress on morpho-physiological characteristics of dragonhead (Dracocephalum moldavica L.) plants. Acta Agric. Slov. 2016, 107, 385–396.
- Yang, K.Y.; Doxey, S.; McLean, J.E.; Britt, D.; Watson, A.; Al Qassy, D.; Anderson, A.J. Remodeling of root morphology by CuO and ZnO nanoparticles: Effects on drought tolerance for plants colonized by a beneficial pseudomonad. Botany 2017, 96, 175–186.
- Nair, P.M.G.; Chung, I.M. Study on the correlation between copper oxide nanoparticles induced growth suppression and enhanced lignification in Indian mustard (Brassica juncea L.). Ecotoxicol. Environ. Saf. 2015, 113, 302–313.
- Liu, F.Y.; Xiong, F.X.; Fan, Y.K.; Li, J.; Wang, H.Z.; Xing, G.M.; He, R. Facile and scalable fabrication engineering of fullerenol nanoparticles by improved alkaline-oxidation approach and its antioxidant potential in maize. J. Nanopart. Res. 2016, 18, 338.
- Borisev, M.; Borisev, I.; Zupunski, M.; Arsenov, D.; Pajevic, S.; Curcic, Z.; Djordjevic, A. Drought impact is alleviated in sugar beets (Beta vulgaris L.) by foliar application of fullerenol nanoparticles. PLoS ONE 2016, 11, e0166248.
- Verma, S.K.; Das, A.K.; Gantait, S.; Kumar, V.; Gurel, E. Applications of carbon nanomaterials in the plant system: A perspective view on the pros and cons. Sci. Total Environ. 2019, 667, 485–499.
- Ahmad, I.; Akhtar, M.S. Use of nanoparticles in alleviating salt stress. In Salt Stress, Microbes, and Plant Interactions: Causes and Solution; Springer: Berlin/Heidelberg, Germany, 2019; pp. 199–215.
- Siddiqi, K.S.; Husen, A. Engineered Gold Nanoparticles and Plant Adaptation Potential. Nanoscale Res. Lett. 2016, 11, 400.
- Ghasemlou, F.; Amiri, H.; Karamian, R.; Mirzaie-asl, A. Alleviation of the effects of on drought stress Verbascum nudicuale by methyl jasmonate and titanium dioxide nanoparticles. Plant Physiol. 2019, 9, 2911–2920.
- Siddiqui, H.; Ahmed, K.B.M.; Sami, F.; Hayat, S. Silicon Nanoparticles and Plants: Current Knowledge and Future Perspectives. In Sustainable Agriculture Reviews; Springer: Berlin/Heidelberg, Germany, 2020; Volume 41, pp. 129–142.
- Rizwan, M.; Ali, S.; Ali, B.; Adrees, M.; Arshad, M.; Hussain, A.; ur Rehman, M.Z.; Waris, A.A. Zinc and iron oxide nanoparticles improved the plant growth and reduced the oxidative stress and cadmium concentration in wheat. Chemosphere 2019, 214, 269–277.
- Khan, Z.S.; Rizwan, M.; Hafeez, M.; Ali, S.; Adrees, M.; Qayyum, M.F.; Khalid, S.; Rehman, M.Z.U.; Sarwar, M.A. Effects of silicon nanoparticles on growth and physiology of wheat in cadmium contaminated soil under different soil moisture levels. Environ. Sci. Pollut. Res. 2019, 27, 4958–4968.
- Jalil, S.U.; Ansari, M.I. Nanoparticles and abiotic stress tolerance in plants: Synthesis, action, and signaling mechanisms. In Plant Signaling Molecules; Woodhead Publishing: Sawston, UK, 2019; pp. 549–561.
- Pérez-De-Luque, A. Interaction of Nanomaterials with Plants: What Do We Need for Real Applications in Agriculture? Front. Environ. Sci. 2017, 5, 12.
- Pérez-Labrada, F.; Hernández-Hernández, H.; López-Pérez, M.C.; González-Morales, S.; BenavidesMendoza, A.; Juárez-Maldonado, A. Chapter 13—Nanoparticles in plants: Morphophysiological, biochemical, and molecular responses. In Plant Life under Changing Environment; Tripathi, D.K., Singh, V.P., Chauhan, D.K., Sharma, S., Prasad, S.M., Dubey, N.K., Ramawat, N., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 289–322.
- Yoshida, T.; Fujita, Y.; Maruyama, K.; Mogami, J.; Todaka, D.; Shinozaki, K.; Yamaguchi-Shinozaki, K.A. Four a rabidopsis AREB/ABF transcription factors function predominantly in gene expression downstream of SnRK2 kinases in abscisic acid signalling in response to osmotic stress. Plant Cell Environ. 2015, 38, 35–49.
- Thiruvengadam, M.; Gurunathan, S.; Chung, I.-M. Physiological, metabolic, and transcriptional effects of biologically synthesized silver nanoparticles in turnip (Brassica rapa ssp. rapa L.). Protoplasma 2015, 252, 1031–1046.
- Dimkpa, C.O.; Andrews, J.; Sanabria, J.; Bindraban, P.S.; Singh, U.; Elmer, W.H.; Gardea-Torresdey, J.L.; White, J.C. Interactive effects of drought, organic fertilizer, and zinc oxide nanoscale and bulk particles on wheat performance and grain nutrient accumulation. Sci. Total Environ. 2020, 722, 137808.
- Ahmadian, K.; Jalilian, J.; Pirzad, A. Nano-fertilizers improved drought tolerance in wheat under deficit irrigation. Agric. Water Manag. 2021, 244, 106544.
- Sun, L.; Song, F.; Guo, J.; Zhu, X.; Liu, S.; Liu, F.; Li, X. Nano-ZnO-Induced Drought Tolerance Is Associated with Melatonin Synthesis and Metabolism in Maize. Int. J. Mol. Sci. 2020, 21, 782.
- Van Nguyen, D.; Nguyen, H.M.; Le, N.T.; Nguyen, K.H.; Nguyen, H.T.; Le, H.M.; Nguyen, A.T.; Dinh, N.T.T.; Hoang, S.A.; Van Ha, C. Copper Nanoparticle Application Enhances Plant Growth and Grain Yield in Maize Under Drought Stress Conditions. J. Plant Growth Regul. 2022, 41, 364–375.
- Esmaili, S.; Tavallali, V.; Amiri, B. Nano-Silicon Complexes Enhance Growth, Yield, Water Relations and Mineral Composition in Tanacetum parthenium under Water Deficit Stress. Silicon 2020, 13, 2493–2508.
- El-Zohri, M.; Al-Wadaani, N.A.; Bafeel, S.O. Foliar Sprayed Green Zinc Oxide Nanoparticles Mitigate Drought-Induced Oxidative Stress in Tomato. Plants 2021, 10, 2400.
More
Information
Contributors
MDPI registered users' name will be linked to their SciProfiles pages. To register with us, please refer to https://encyclopedia.pub/register
:
View Times:
1.7K
Revisions:
3 times
(View History)
Update Date:
22 Nov 2022
Table of Contents


Notice
You are not a member of the advisory board for this topic. If you want to update advisory board member profile, please contact office@encyclopedia.pub.
OK
Confirm
Only members of the Encyclopedia advisory board for this topic are allowed to note entries. Would you like to become an advisory board member of the Encyclopedia?
Yes
No
${ textCharacter }/${ maxCharacter }
Submit
Cancel
Back
Comments
${ item }
|
${ item.createdUser.fullName }
${ item.createdAt }
${ item.vote }
${ item.reply }
Delete
${ reply.createdUser.fullName }
${ reply.createdAt }
${ reply.vote }
Delete
There is no reply to this comment~
${ item.replyTextCharacter }/${ item.replyMaxCharacter }
Submit
Cancel
More
No more~
There is no comment~
${ textCharacter }/${ maxCharacter }
Submit
Cancel
${ selectedItem.replyTextCharacter }/${ selectedItem.replyMaxCharacter }
Submit
Cancel
Confirm
Are you sure to Delete?
Yes
No