 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Leonardo Luis Fruttero | + 3220 word(s) | 3220 | 2020-11-25 03:48:56 | | | |
| 2 | Vivi Li | -19 word(s) | 3201 | 2020-12-14 05:02:04 | | | | |
| 3 | Vivi Li | + 1 word(s) | 3202 | 2020-12-15 07:07:05 | | | | |
| 4 | Vivi Li | + 2 word(s) | 3203 | 2020-12-15 07:08:24 | | |
Video Upload Options
Biopesticides are a type of pesticides derived from natural materials including plants and bacteria. In this entry, we reviewed the structural and biological features of Jaburetox and Soyuretox, two plant urease-derived recombinant peptides that present entomotoxic and fungitoxic effects without harming beneficial species or the environment. We also discussed other bioactive peptides in the context of their action mechanism.
1. Jaburetox and Soyuretox: Historical Aspects and Potential as Biopesticides
1.1. Transgenic Plants Expressing Biocide Polypeptides and Plant Defense
Hunger continues to afflict mainly the poorer countries around the world. In 2016, 10.7% of the world population were chronically undernourished (www.worldhunger.org). According to data provided by the World Bank (www.databank.worldbank.org), over the last 15 years, the word population has increased with an annual growth rate of ~1% (1.075 in 2019), from 6.59 (2006) to 7.67 (2019) billions inhabitants. With the increase in life expectancy, from 69.2 (2006) to 72.5 (2018) years, particularly in the richer countries, even considering a decrease of growth rate, estimates are that the world population could reach 9.7 billion (www.population.un.org) in 2050. By then, food demand will be 60% higher (www.webforum.org). As agricultural land is finite (it increased only from 47.18 in 2006 to 48.43 in 2016 millions square kilometers according to the World Bank), and is expected to shrink due to urbanization, climate change and soil degradation, increases in food production will require an even more efficient agriculture. The output of agriculture is hampered, however, by losses in the field or after harvesting, by a variety of insect pests, nematodes, fungi and diseases induced by bacteria or viruses [1][2]. Herbivores alone feeding on foliage, sap and root can decrease more than 20% of net plant productivity and food losses to insects are expected to even grow in a scenario of global warming [3].
To efficiently control insect pests in agriculture, combining different strategies is frequently required, including the use of resistant crop varieties. When there is no natural plant genotypes genetically resistant to insect pests, development of genetically modified (GM) resistant plants is an option. A milestone in the development of insect-resistant crops was established in the late 1980′s, by genetically engineering a tobacco plant to express an entomotoxic protein from the bacterium Bacillus thuringiensis (Bt) [4][5]. In 2018, the list of genetically modified plants that were commercialized had 26 species, either tolerant to herbicides or with increased resistance to insects (most expressing Bt toxins), grown or imported in 75 countries (www.gmoanswers.com). Bt toxins (or Cry proteins) have some restrictions, such as the low toxicity against sap-sucking insects [6][7]. Moreover, an increasing level of resistance of insects against Cry insecticidal proteins has been reported [8]. Fortunately, there is a number of plant entomotoxic proteins that can be used instead or in synergy with the Bt technology to control insect pests in new generations of transgenic plants yet to be developed [9].
This is the case of Jaburetox and Soyuretox, recombinant peptides derived from Jack Bean (Canavalia ensiformis) and Soybean (Glycine max) ureases, respectively (doi:10.1016/j.peptides.2007.08.009, doi:10.3390/ijms20215401). These peptides are intrinsically disordered proteins (IDP’s), proteins that do not have a stable 3D structure but can present relevant biological activities (doi:10.1146/annurev-biochem-072711-164947, doi:10.1007/s10930-009-9201-4). Jaburetox and Soyuretox have entomotoxic effects that can contribute to the rational control of insect pests without harming other animals (doi:10.1016/j.peptides.2007.08.009, doi:10.1016/j.bbagen.2013.11.010). Studies evaluating transgenic crops that express the peptides have shown that these plants were less susceptible to insects and nematodes indicating an enormous potential for IDP's on pest control (https://doi.org/10.1016/j.biori.2019.12.002).
1.2. Plant Proteins and Peptides with Insecticidal and Fungitoxic Properties
Insect pests and phytopathogenic fungi are detrimental to several crops and cause significant economic losses in agriculture worldwide. To cope with herbivory and fungal diseases, plants have evolved sophisticated defense mechanisms. Plant tissues accumulate, constitutively or after induction, various classes of defensive compounds that confer resistance against herbivores and infection by fungi, bacteria, viruses, as well as nematodes. The most known plant proteins involved in defense mechanisms against insect pests include lectins [10], ribosome-inactivating proteins of types 1 and 2 [11], inhibitors of proteolytic enzymes and glycohydrolases [12], modified forms of storage proteins [13][14], among others. Several plant peptides display antifungal properties such as defensins [15], lipid transport proteins [16], chitinases [17], lectins [18], thionins [19], cyclopeptide alkaloids [20] and other less common types. For a general review on these topics please refer to Dang and Van Damme [21] and Grossi-De-Sá et al. [9].
Ureases represent another group of plant proteins with insecticidal and antifungal properties which widen the proposed physiological roles of these enzymes [22][23][24][25]. Ureases (urea amidohydrolase; EC 3.5.1.5) are well conserved and nickel-dependent enzymes that catalyze urea hydrolysis into ammonia and carbon dioxide, synthesized by plants, fungi and bacteria [26][27][28][29]. Canatoxin is a less abundant urease isoform isolated from Canavalia ensiformis (jack bean) seeds [30][31]. Structurally similar to the seed’s major urease, both proteins display insecticidal and antifungal properties independent of their ureolytic activity [23][26][27]. Soybean (Glycine max) and pigeon pea (Cajanus cajan) ureases were also shown to display insecticidal [32][33][34] and antifungal [32][35][36] properties. Noteworthy, ureases are insecticidal against hemipteran pests (such as the stink bug Nezara viridula and the cotton stainer bug Dysdercus peruvianus), which were not susceptible to the entomotoxic activity of Cry toxins from B. thuringiensis [6][7]. Since these proteins are abundant in many edible vegetables, particularly in legumes, they can be generally regarded as biosafe [37].
1.3. Ureases and Derived Peptides as Sources of Insecticidal and Fungitoxic (Poly)Peptides
The insecticidal [38] and fungitoxic [39] effects of canatoxin were described before its characterization as an isoform of jack bean urease (JBU) [31]. In the first study of canatoxin’s insecticidal effect, it became clear that only insects relying on cathepsin-like digestive enzymes (such as the cowpea weevil Callosobruchus maculatus and the kissing bug Rhodnius prolixus) are sensitive to the toxin, while insects with digestion based on trypsin-like enzymes (such as the tobacco hornworn Manduca sexta or the fruitfly Drosophila melanogaster) show no susceptibility. The hypothesis of a proteolytic activation of the toxin was then proposed [38]. The hydrolysis of canatoxin with C. maculatus digestive enzymes yielded a 10 kDa entomotoxic peptide named pepcanatox [40]. Our group has demonstrated through inhibition of cathepsin-like enzymes, that the enzymatic activity of cathepsin B (cysteine proteinase) and cathepsin D (aspartic proteinases) is necessary for the release of toxic fragments of Canatoxin [38][40]. Cathepsin B is a cysteine proteinase that can act as an exopeptidase or endopeptidase at acidic pH [41]. Cleavage by cathepsin B has a preference for basic and hydrophobic amino acids [42]. Meanwhile, cathepsin D cleavage occurs at acidic pH and has a preference for hydrophobic residues [43]. Subsequently, the major proteolytic activities of midgut homogenates of D. peruvianus nymphs were shown in vitro to catalyze the release of pepcanatox from JBU [44]. Cysteine, aspartic and metalloproteinases are present in both homogenates. Fluorogenic substrates containing JBU partial sequences flanking the N-terminal or the C-terminal portion of the entomotoxic peptide were efficiently cleaved by the D. peruvianus nymph midgut homogenates. Different classes of enzymes in the homogenates cleaved both substrates suggesting that in vivo the release of the entomotoxic peptide results from the concerted action of at least two different proteinases [44].
Jaburetox-2Ec, a recombinant peptide with 93 amino acids (~11 kDa) equivalent to pepcanatox, was produced heterologously in Escherichia coli from the corresponding sequence of the JBU isoform JBURE-II [45]. Here the term “peptide” is used solely to emphasize the fact that it is a fragment of a much larger protein regardless of its molecular mass. Later on, the peptide called simply Jaburetox was developed, with the same urease-derived sequence and the 6 His tail found in Jaburetox-2Ec (Figure 1), but lacking the V5 epitope present in the latter [46]. Both peptides, Jaburetox-2Ec and Jaburetox, display equivalent insecticidal activity, evidencing that the epitope V5 is not implied in their entomotoxicity [46].
Since one of the most well studied mechanisms of defense against insect pest is digestive enzyme inhibition, this possibility was explored by our group for ureases and derived peptides. As described by Carlini et al. [38] and Ferreira da-Silva et al. [40], canatoxin showed no inhibitory effect on the proteolytic (cathepsin B or D-like) or α-amylase activities. Moreover, the peptides derived from Canatoxin’s digestion with cathepsin-like enzymes, including pepcanatox, did not display either proteolytic or amylase inhibitory properties [47]. Although Jaburetox itself was not tested, taking into account its virtually identical sequence when compared to pepcanatox, it is safe to assume that Jaburetox has no inhibitory effects upon digestive enzymes.
Jaburetox is lethal to several insects susceptible to canatoxin (the cotton stainer bug D. peruvianus, the kissing bugs R. prolixus and Triatoma infestans) and also kills insects that are resistant to intact ureases, such as lepidopterans (fall armyworm Spodoptora frugiperda, cotton bollworm Helicoverpa armigera) and dipterans (Aedes aegypti) [45][48], because the hydrolysis of the protein to release the peptide is no longer required.
Concerning the antifungal property of ureases and derived peptides, the most abundant jack bean isoform, JBU, was shown to display antifungal properties against a panel of 16 phytopathogenic filamentous fungi species of 11 genera, blocking spore germination and/or mycelial growth, and inhibiting multiplication of yeasts at submicromolar concentrations [35][49]. Jaburetox also displayed antifungal properties against filamentous fungi and yeasts [49].
Antifungal effects were observed in vitro also for isoforms of soybean urease (SBU) [35][50]. The participation of ureases in plant defense against fungal diseases was demonstrated in urease-null soybean plants obtained by gene silencing [51]. Later, the peptide called Soyuretox, homologous to Jaburetox (Figure 1), but derived from the ubiquitous isoform of the soybean urease, was heterologously expressed in E. coli, characterized structurally and its entomotoxic and antifungal effects were demonstrated [52].
Jaburetox showed no acute toxicity to mice and rats [45] and was found not toxic in a risk assessment study [53] while Soyuretox was not toxic to zebrafish embryos [52]. These data suggest that these peptides may be safe alternatives to attain resistance to insect herbivory and/or fungal disease in transgenic plants.
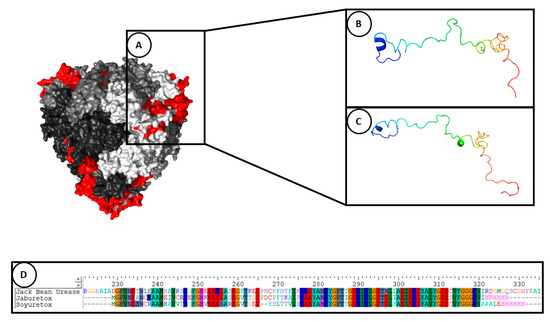
Figure 1. Structural representation of entomotoxic peptides and Jack Bean Urease. (A) Graphical representation of the location of the peptides (red) in the protein structure of Jack Bean Urease (pdb: 3LA4). Each monomer of the JBU hexamer is represented with a different shade of grey. (B) Jaburetox, (C) Soyuretox and (D) comparison of the primary sequences.
2. Structural Aspects of Jaburetox and Soyuretox and their impact in the biological activity
A series of studies were conducted in order to characterize the structural properties of Jaburetox and Soyuretox, including the use of different techniques such as Molecular Dynamics simulation, Light Scattering, Circular Dichroism (CD) and Nuclear Magnetic Resonance (NMR). NMR reveals that Jaburetox is an IDP, showing low signal dispersion in the proton dimension. The SSP (Secondary Structure Propensity) analysis of chemical shifts generated by NMR predicted that the peptide is widely disordered with a small tendency to form α-structures. Jaburetox exhibits a large hydrodynamic radius, suggestive of a disordered polypeptide, through dynamic light scattering analysis (doi.org/10.1111/febs.13201). CD demonstrated that Jaburetox increased its secondary structure content when in contact with SDS-micelles and large unilamellar vesicles (doi.org/10.1016/j.colsurfb.2017.08.053). The HSQC NMR spectrum for Soyuretox also showed low signal dispersion in the proton dimension, revealing its IDP behavior, assuming a more ordered structure of the molecule when in the presence of SDS micelles (10 mM). CD showed disordered behavior for both peptide at pH 6.5 although Soyuretox acquired some secondary structure at pH 8 (doi.org/10.3390/ijms20215401). Despite this structural difference, both peptides showed similar entomotoxic and fungitoxic effects.
3. Structural Aspects of Other Intrinsically Disordered Bioactive Polypeptides
Even though approximately 30% of eukaryote proteins have disordered regions composed of fifty or more amino acid residues [54], there are relatively few reports of antimicrobial and insecticidal IDPs in the literature. Without aiming to be exhaustive, we briefly discuss here some examples and establish comparisons to Jaburetox and Soyuretox when relevant.
Plants are an inexhaustible source of bioactive molecules, including those that are part of their highly evolved defense mechanisms [22]. One important example are the cyclotides, naturally occurring macrocyclic peptides found in several families of plants. They present a unique head-to-tail cyclized backbone, stabilized by three disulfide bonds forming a cystine knot. This arrangement makes cyclotides exceptionally stable against chemical, thermal and biological degradation. These macromolecules are able to cross cellular membranes and control intracellular protein-protein interactions, enabling them to act upon different targets [55]. Cyclotides present diverse host-defense roles including insecticidal activity and it is believed that this property is derived from their ability to bind to membranes and form pores [56]. Kalata B1 is the most studied cyclotide, derived from the African plant Oldelandia affinis [57][58]. Daly et al. [59] reported that the N-terminal pro-domain of the kalata B1 precursor is intrinsically unstructured. This terminal region induces the self-association of the precursor to form a dimeric structure, which can, in turn, be determinant for the role of the N-terminal as a vacuolar-targeting signal. According to the authors, the disorder in the terminal region could be linked either to the fact that it is a functional segment with higher mobility or because it partially folds upon binding to a target, as could be also the case for Jaburetox and Soyuretox [52][60] Thus, the pore-forming capacity seems to be part of the toxic mechanism of both, urease-derived peptides and cyclotides. However, the intrinsically disordered nature of the cyclotides does not seem to be related to their insecticidal effect but rather to a role in signaling. The relevance of IDPs in signal transduction in plants is better documented and, in this case, it is believed that the intrinsically disordered nature is necessary to confer the low affinity and high specificity needed to perform the required interactions [61].
Concerning antifungal IDPs, histatins are a family of small, histidine-rich, cationic proteins present in mammalian saliva that constitute the first line of defense against oral candidiasis caused by C. albicans and to other pathogenic fungi. Histatin 5, an intrinsically disordered model protein, is the major histatin component of the unstimulated parotid secretion and the most potent antifungal protein of all the histatin family [62][63]. Histatin-5 has antifungal activity against C. albicans at 15 µM [64], a similar fungitoxic concentration for Jaburetox and Soyuretox [49][52]. The physiological concentration of histatin-5 in human saliva is 15 to 50 µM, while the concentration of protein required to kill half of maximum number of cells (ED50) is 1.4 µM. There is an extensive debate regarding the mode of action of this protein, with evidences pointing against pore formation or membrane lysis. The targets of histatin-5 appear to be intracellular and, once taken up by cells, it affects mitochondrial functions causing oxidative stress and ultimately killing the cells by ion imbalance and volume dysregulation induced by osmotic stress [65]. In addition, this peptide is related to depletion of intracellular ATP content and also oxidative damage due to ROS formation in intracellular organelles [66]. The production of oxidative molecules by histatin-5 is a common aspect with the mode of actions of Jaburetox and Soyuretox, which induced ROS generation both in insect hemocytes [67] and in C. albicans cells [52]. Since the fungitoxic mode of action of histatin-5 is not completely understood, it is not clear how the intrinsically disordered nature of the protein participates in the process. Nevertheless, histatin-5 mechanism of action against C. albicans is similar to what is known so far for Jaburetox and Soyuretox, including membrane interaction and permeabilization, and ROS formation. There is also evidence that Jaburetox is taken up by hemocytes [68], thus suggesting intracellular targets.
Hornerin is an IDP of 254 kDa that belongs to the S100-fused-type family. This protein is believed to be one of the main reasons why healthy human skin is remarkably resistant towards the infection by Pseudomonas aeruginosa, an environmental opportunistic pathogen widespread in water and soil [69]. Recently, fragments of hornerin were characterized as potent microbicidal agents and that this feature is maintained, independent of the amino acid sequence, provided they are linear cationic peptides containing a high percentage of disorder-promoting amino acids and a low percentage of order-promoting ones. The authors reported that the antimicrobial capacity of these cationic intrinsically disordered antimicrobial peptides (CIDAMPs) depends on their chain length, net charge, lipidation and environmental conditions [70]. The CIDAMPs have an intracellular mode of action, as hornerin transverses bacterial membranes by an energy-dependent mechanism and accumulates in the cytoplasm. The molecular targets of CIDAMPs seems to be different sites of the protein synthesis machinery [71]. The described features of CIDAMPS and the other IDP active peptides are summarized in Table 1.
Table 1. Other intrinsically disordered proteins with biological activities.
| IDP | Source | Biological Activity | Disorder Region | Reference |
|---|---|---|---|---|
| Kalata B1 | African plant Oldelandia affinis | Signaling, ability to bind and to form pores in membranes | N-terminal pro-domain | [59] |
| Histatin 5 | Mammalian saliva | Antifungal activity | No defined structure in solution | [65][66] |
| Hornerin | Human skin | Antibacterial activity | Almost all the protein is unstructured, except the N-terminus. Cationic peptides generated from hornerin present antimicrobial activity | [70] |
4. Biotechnological Applications and Perspectives
The use of GM crops resistant to pests such as fungi, nematodes and insects is an appealing strategy considering the current need of efficiently increasing the yield of the agricultural production with less impact in the environment and health [72]. Since their discovery in the 80′s, the use of transgenic crops has been dominated by the Bt technology. Nevertheless, some insect species are not susceptible to them and its intensive application has led to the development of resistance [7][73]. Considering the fact that IDPs do not need to fold in a proper way to be biologically active, this feature can potentially be an advantage regarding their expression in transgenic plants. In the case of Jaburetox, its disordered structure gives the peptide the capacity to withstand a vast range of temperatures and pH without losing its biological activity [60], a desirable feature for a biotechnological tool. Moreover, the conformational flexibility of Jaburetox and Soyuretox allows them to interact with several binding partners with different subcellular distributions, leading ultimately to diverse targets. This feature gives them the ability of avoiding or at least delaying the generation of resistance.
In this context, three types of transgenic crops expressing urease-derived peptides have been developed with promising results [74][75][76]. Soybean plants overexpressing Soyuretox were challenged with the root-knot nematode Meloidogyne javanica, a major agricultural pest in several countries [76]. As a result, the average reproductive factor of the nematode was significantly reduced. On the other hand, Didoné [74] reported that maize (Zea mays) expressing Jaburetox fed to the important polyphagous pest S. frugiperda led not only to a 39% lethality of larvae, but also to other sub-lethal statistically significant effects, such as body weight reduction, decreased ingestion and remarkably, fertility decline. In addition, Ceccon [75] demonstrated that Jaburetox-expressing tobacco plants also produced high mortality and a pronounced reduction in the leaf consumption by the lepidopteran H. armigera. These authors started the development of gene stacking strategy, with plants expressing simultaneously Jaburetox and a double-stranded RNA complementary to the rieske gene, which has the advantage of diminishing the possibility of resistance development events [74]. The use of plants overexpressing Soyuretox or Jaburetox would, in principle, be a way to avoid harming beneficial or innocuous insects, since only those species that fed on the plants would be affected. Nevertheless, since off-target effects could be an issue related to the broad insecticidal activity of Jaburetox/Soyuretox, transgenic crops can be improved using tissue-specific or damage-induced promoters [77][78].
These latest studies [74][75][76] are a proof of concept that IDPs in general, and the urease-derived peptides in particular, are very attractive pesticides that can be engineered for use as an effective and environmental-friendly strategy, alone or in combination with other IDPs or toxic molecules. The multidisciplinary research approaches employed by our group and collaborators improved significantly the understanding of the structural and biological aspects of these IDPs and encourage us to pursue a full comprehension of their mechanism of action that would, ultimately, facilitate their application in the field.
References
- Mitchell, C.; Brennan, R.M.; Graham, J.; Karley, A.J. Plant defense against herbivorous pests: Exploiting resistance and tolerance traits for sustainable crop protection. Front. Plant Sci. 2016, 7, 1132.
- Oerke, E.C. Crop losses to pests. J. Agric. Sci. 2006, 144, 31–43.
- Gregory, P.J.; Johnson, S.N.; Newton, A.C.; Ingram, J.S.I. Integrating pests and pathogens into the climate change/food security debate. J. Exp. Bot. 2009, 60, 2827–2838.
- Ahmad, P.; Ashraf, M.; Younis, M.; Hu, X.; Kumar, A.; Akram, N.A.; Al-Qurainy, F. Role of transgenic plants in agriculture and biopharming. Biotechnol. Adv. 2012, 30, 524–540.
- Palma, L.; Muñoz, D.; Berry, C.; Murillo, J.; Caballero, P.; Caballero, P. Bacillus thuringiensis toxins: An overview of their biocidal activity. Toxins 2014, 6, 3296–3325.
- Chougule, N.P.; Bonning, B.C. Toxins for transgenic resistance to hemipteran pests. Toxins 2012, 4, 405–429.
- Heckel, D.G. How do toxins from Bacillus thuringiensis kill insects? An evolutionary perspective. Arch. Insect Biochem. Physiol. 2020, 104, e21673.
- Wei, J.; Zhang, Y.; An, S. The progress in insect cross-resistance among Bacillus thuringiensis toxins. Arch. Insect Biochem. Physiol. 2019, 102, e21547.
- Grossi-de-Sá, M.F.; Pelegrini, P.B.; Vasconcelos, I.M.; Carlini, C.R.; Silva, M.S. Entomotoxic Plant Proteins: Potential Molecules to Develop Genetically Modified Plants Resistant to Insect-Pests. In Plant Toxins; Carlini, C.R., Ligabue-Braun, R., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2017; pp. 415–447. ISBN 978-94-007-6464-4.
- Tsaneva, M.; Van Damme, E.J.M. 130 years of Plant Lectin Research. Glycoconj. J. 2020, 37, 533–551.
- Bolognesi, A.; Bortolotti, M.; Maiello, S.; Battelli, M.G.; Polito, L. Ribosome-inactivating proteins from plants: A historical overview. Molecules 2016, 21, 1627.
- Zhu-Salzman, K.; Zeng, R. Insect Response to Plant Defensive Protease Inhibitors. Annu. Rev. Entomol. 2015, 60, 233–252.
- Souza Cândido, E.; Pinto, M.F.S.; Pelegrini, P.B.; Lima, T.B.; Silva, O.N.; Pogue, R.; Grossi-de-Sá, M.F.; Franco, O.L. Plant storage proteins with antimicrobial activity: Novel insights into plant defense mechanisms. FASEB J. 2011, 25, 3290–3305.
- Sales, M.P.; Gerhardt, I.R.; Grossi-de-Sa, M.F.; Xavier-Filho, J. Do legume storage proteins play a role in defending seeds against Bruchids? Plant Physiol. 2000, 124, 515–522.
- Vriens, K.; Cammue, B.P.A.; Thevissen, K. Antifungal plant defensins: Mechanisms of action and production. Molecules 2014, 19, 12280–12303.
- Carvalho, A.d.O.; Gomes, V.M. Role of plant lipid transfer proteins in plant cell physiology—A concise review. Peptides 2007, 28, 1144–1153.
- Grover, A. Plant Chitinases: Genetic Diversity and Physiological Roles. CRC. Crit. Rev. Plant Sci. 2012, 31, 57–73.
- Breitenbach Barroso Coelho, L.C.; Marcelino dos Santos Silva, P.; Felix de Oliveira, W.; de Moura, M.C.; Viana Pontual, E.; Soares Gomes, F.; Guedes Paiva, P.M.; Napoleão, T.H.; dos Santos Correia, M.T. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252.
- Stec, B. Plant thionins—The structural perspective. Cell. Mol. Life Sci. 2006, 63, 1370–1385.
- Khan, H.; Mubarak, M.S.; Amin, S. Antifungal Potential of Alkaloids as an Emerging Therapeutic Target. Curr. Drug Targets 2016, 18.
- Dang, L.; Van Damme, E.J.M. Toxic proteins in plants. Phytochemistry 2015, 117, 51–64.
- Carlini, C.R.; Grossi-De-Sá, M.F. Plant toxic proteins with insecticidal properties. A review on their potentialities as bioinsecticides. Toxicon 2002, 40, 1515–1539.
- Carlini, C.R.; Ligabue-Braun, R. Ureases as multifunctional toxic proteins: A review. Toxicon 2016, 110, 90–109.
- Carlini, C.R.; Polacco, J.C. Toxic properties of urease. Crop Sci. 2008, 48, 1665–1672.
- Stanisçuaski, F.; Carlini, C.R. Plant ureases and related peptides: Understanding their entomotoxic properties. Toxins 2012, 4, 55–67.
- Kappaun, K.; Piovesan, A.R.; Carlini, C.R.; Ligabue-Braun, R. Ureases: Historical aspects, catalytic, and non-catalytic properties—A review. J. Adv. Res. 2018, 13, 3–17.
- Ligabue-Braun, R.; Carlini, C.R. Moonlighting Toxins: Ureases and Beyond. In Plant Toxins; Springer Netherlands: Dordrecht, The Netherlands, 2015; pp. 1–21.
- Mazzei, L.; Musiani, F.; Ciurli, S. Urease. In RSC Metallobiology series “The Biological Chemistry of Nickel”; The Royal Society of Chemistry: London, UK, 2017; ISBN 1555812139.
- Mobley, H.L.T.; Island, M.D.; Hausinger, R.P. Molecular biology of microbial ureases. Microbiol. Rev. 1995, 59, 451–480.
- Carlini, C.R.; Guimarães, J.A. Isolation and characterization of a toxic protein from Canavalia ensiformis (jack bean) seeds, distinct from concanavalin A. Toxicon 1981, 19, 667–675.
- Follmer, C.; Barcellos, G.B.S.; Zingali, R.B.; Machado, O.L.T.; Alves, E.W.; Barja-Fidalgo, C.; Guimarães, J.A.; Carlini, C.R. Canatoxin, a toxic protein from jack beans (Canavalia ensiformis), is a variant form of urease (EC 3.5.1.5): Biological effects of urease independent of its ureolytic activity. Biochem. J. 2001, 360, 217–224.
- Balasubramanian, A.; Durairajpandian, V.; Elumalai, S.; Mathivanan, N.; Munirajan, A.K.; Ponnuraj, K. Structural and functional studies on urease from pigeon pea (Cajanus cajan). Int. J. Biol. Macromol. 2013, 58, 301–309.
- Follmer, C.; Real-Guerra, R.; Wasserman, G.E.; Olivera-Severo, D.; Carlini, C.R. Jackbean, soybean and Bacillus pasteurii ureases: Biological effects unrelated to ureolytic activity. Eur. J. Biochem. 2004, 271, 1357–1363.
- Real-Guerra, R.; Staniscuaski, F.; Carlini, C.R. Soybean Urease: Over a Hundred Years of Knowledge. In A Comprehensive Survey of International Soybean Research—Genetics, Physiology, Agronomy and Nitrogen Relationships; Board, J.E., Ed.; InTech: London, UK, 2013.
- Becker-Ritt, A.B.; Martinelli, A.H.S.; Mitidieri, S.; Feder, V.; Wassermann, G.E.; Santi, L.; Vainstein, M.H.; Oliveira, J.T.A.; Fiuza, L.M.; Pasquali, G.; et al. Antifungal activity of plant and bacterial ureases. Toxicon 2007, 50, 971–983.
- Martinelli, A.H.S.; Lopes, F.C.; Broll, V.; Defferrari, M.S.; Ligabue-Braun, R.; Kappaun, K.; Tichota, D.M.; Fruttero, L.L.; Moyetta, N.R.; Demartini, D.R.; et al. Soybean ubiquitous urease with purification facilitator: An addition to the moonlighting studies toolbox. Process Biochem. 2017, 53, 245–258.
- Follmer, C. Insights into the role and structure of plant ureases. Phytochemistry 2008, 69, 18–28.
- Carlini, C.R.; Oliveira, A.E.; Azambuja, P.; Xavier-Filho, J.; Wells, M.A. Biological effects of canatoxin in different insect models: Evidence for a proteolytic activation of the toxin by insect cathepsinlike enzymes. J. Econ. Entomol. 1997, 90, 340–348.
- Oliveira, A.E.A.; Gomes, V.M.; Sales, M.P.; Fernandes, K.V.S.; Carlini, C.R.; Xavier-Filho, J. The toxicity of jack bean [Canavalia ensiformis (L.) DC.] canatoxin to plant pathogenic fungi. Rev. Bras. Biol. 1999, 59, 59–62.
- Ferreira-DaSilva, C.T.; Gombarovits, M.E.C.; Masuda, H.; Oliveira, C.M.; Carlini, C.R. Proteolytic activation of canatoxin, a plant toxic protein, by insect cathepsin-like enzymes. Arch. Insect Biochem. Physiol. 2000, 44, 162–171.
- Polgár, L.; Csoma, C. Dissociation of ionizing groups in the binding cleft inversely controls the endo- and exopeptidase activities of cathepsin B. J. Biol. Chem. 1987, 262, 14448–14453.
- Turk, D.; Gunčar, G.; Podobnik, M.; Turk, B. Revised definition of substrate binding sites of papain-like cysteine proteases. Biol. Chem. 1998, 379, 137–147.
- Sun, H.; Lou, X.; Shan, Q.; Zhang, J.; Zhu, X.; Zhang, J.; Wang, Y.; Xie, Y.; Xu, N.; Liu, S. Proteolytic Characteristics of Cathepsin D Related to the Recognition and Cleavage of Its Target Proteins. PLoS ONE 2013, 8.
- Piovesan, A.R.; Stanisçuaski, F.; Marco-Salvadori, J.; Real-Guerra, R.; Defferrari, M.S.; Carlini, C.R. Stage-specific gut proteinases of the cotton stainer bug Dysdercus peruvianus: Role in the release of entomotoxic peptides from Canavalia ensiformis urease. Insect Biochem. Mol. Biol. 2008, 38, 1023–1032.
- Mulinari, F.; Stanisçuaski, F.; Bertholdo-Vargas, L.R.; Postal, M.; Oliveira-Neto, O.B.; Rigden, D.J.; Grossi-de-Sá, M.F.; Carlini, C.R. Jaburetox-2Ec: An insecticidal peptide derived from an isoform of urease from the plant Canavalia ensiformis. Peptides 2007, 28, 2042–2050.
- Martinelli, A.H.S.; Kappaun, K.; Ligabue-Braun, R.; Defferrari, M.S.; Piovesan, A.R.; Stanisçuaski, F.; Demartini, D.R.; Dal Belo, C.A.; Almeida, C.G.M.; Follmer, C.; et al. Structure-function studies on jaburetox, a recombinant insecticidal peptide derived from jack bean (Canavalia ensiformis) urease. Biochim. Biophys. Acta Gen. Subj. 2014, 1840, 935–944.
- Gombarovits, M. Peptídeos entomotóxicos gerados a partir da CNTX: Obtenção, isolamento, propriedades biológicas e caracterização físico-química. Ph.D. Thesis, Universidade Federal do Rio de Janeiro, Rio de Janeiro, Brazil, 1999.
- Becker-Ritt, A.B.; Portugal, C.S.; Carlini, C.R. Jaburetox: Update on a urease-derived peptide. J. Venom. Anim. Toxins Incl. Trop. Dis. 2017, 23, 1–8.
- Postal, M.; Martinelli, A.H.S.; Becker-Ritt, A.B.; Ligabue-Braun, R.; Demartini, D.R.; Ribeiro, S.F.F.; Pasquali, G.; Gomes, V.M.; Carlini, C.R. Antifungal properties of Canavalia ensiformis urease and derived peptides. Peptides 2012, 38, 22–32.
- Broll, V.; Martinelli, A.H.S.; Lopes, F.C.; Fruttero, L.L.; Zambelli, B.; Salladini, E.; Dobrovolska, O.; Ciurli, S.; Carlini, C.R. Structural analysis of the interaction between Jaburetox, an intrinsically disordered protein, and membrane models. Colloids Surfaces B Biointerfaces 2017, 159, 849–860.
- Wiebke-Strohm, B.; Pasquali, G.; Margis-Pinheiro, M.; Bencke, M.; Bücker-Neto, L.; Becker-Ritt, A.B.; Martinelli, A.H.S.; Rechenmacher, C.; Polacco, J.C.; Stolf, R.; et al. Ubiquitous urease affects soybean susceptibility to fungi. Plant Mol. Biol. 2012, 79, 75–87.
- Kappaun, K.; Martinelli, A.H.S.; Broll, V.; Zambelli, B.; Lopes, F.C.; Ligabue-Braun, R.; Fruttero, L.L.; Moyetta, N.R.; Bonan, C.D.; Carlini, C.R.; et al. Soyuretox, an intrinsically disordered polypeptide derived from soybean (Glycine max) ubiquitous urease with potential use as a biopesticide. Int. J. Mol. Sci. 2019, 20, 5401.
- Sá, C.A.; Vieira, L.R.; Pereira Almeida Filho, L.C.; Real-Guerra, R.; Lopes, F.C.; Souza, T.M.; Vasconcelos, I.M.; Staniscuaski, F.; Carlini, C.R.; Urano Carvalho, A.F.; et al. Risk assessment of the antifungal and insecticidal peptide Jaburetox and its parental protein the Jack bean (Canavalia ensiformis) urease. Food Chem. Toxicol. 2020, 136, 110977.
- Dunker, A.K.; Lawson, J.D.; Brown, C.J.; Williams, R.M.; Romero, P.; Oh, J.S.; Oldfield, C.J.; Campen, A.M.; Ratliff, C.M.; Hipps, K.W.; et al. Intrinsically disordered protein. J. Mol. Graph. Model. 2001, 19, 26–59.
- Narayani, M.; Babu, R.; Chadha, A.; Srivastava, S. Production of bioactive cyclotides: A comprehensive overview. Phytochem. Rev. 2020, 19, 787–825.
- Huang, Y.H.; Colgrave, M.L.; Daly, N.L.; Keleshian, A.; Martinac, B.; Craik, D.J. The biological activity of the prototypic cyclotide Kalata B1 is modulated by the formation of multimeric pores. J. Biol. Chem. 2009, 284, 20699–20707.
- Saether, O.; Craik, D.J.; Campbell, I.D.; Sletten, K.; Juul, J.; Norman, D.G. Elucidation of the Primary and Three-Dimensional Structure of the Uterotonic Polypeptide Kalata B1. Biochemistry 1995, 34, 4147–4158.
- Plan, M.R.R.; Göransson, U.; Clark, R.J.; Daly, N.L.; Colgrave, M.L.; Craik, D.J. The cyclotide fingerprint in Oldenlandia affinis: Elucidation of chemically modified, linear and novel macrocyclic peptides. ChemBioChem 2007, 8, 1001–1011.
- Daly, N.L.; Gunasekera, S.; Clark, R.J.; Lin, F.; Wade, J.D.; Anderson, M.A.; Craik, D.J. The N-terminal pro-domain of the kalata B1 cyclotide precursor is intrinsically unstructured. Biopolymers 2016, 106, 825–833.
- Lopes, F.C.; Dobrovolska, O.; Real-Guerra, R.; Broll, V.; Zambelli, B.; Musiani, F.; Uversky, V.N.; Carlini, C.R.; Ciurli, S. Pliable natural biocide: Jaburetox is an intrinsically disordered insecticidal and fungicidal polypeptide derived from jack bean urease. FEBS J. 2015, 282, 1043–1064.
- Marín, M.; Ott, T. Intrinsic disorder in plant proteins and phytopathogenic bacterial effectors. Chem. Rev. 2014, 114, 6912–6932.
- Oppenheim, F.G.; Xu, T.; McMillian, F.M.; Levitz, S.M.; Diamond, R.D.; Offner, G.D.; Troxler, R.F. Histatins, a novel family of histidine-rich proteins in human parotid secretion. Isolation, characterization, primary structure, and fungistatic effects on Candida albicans. J. Biol. Chem. 1988, 263, 7472–7477.
- Jephthah, S.; Staby, L.; Kragelund, B.B.; Skepö, M. Temperature Dependence of Intrinsically Disordered Proteins in Simulations: What are We Missing? J. Chem. Theory Comput. 2019, 15, 2672–2683.
- Tsai, H.; Raj, P.A.; Bobek, L.A. Candidacidal activity of recombinant human salivary histatin-5 and variants. Infect. Immun. 1996, 64, 5000–5007.
- Puri, S.; Edgerton, M. How does it kill?: Understanding the candidacidal mechanism of salivary histatin 5. Eukaryot. Cell 2014, 13, 958–964.
- Melino, S.; Santone, C.; Di Nardo, P.; Sarkar, B. Histatins: Salivary peptides with copper (II)-and zinc (II)-binding motifs: Perspectives for biomedical applications. FEBS J. 2014, 281, 657–672.
- Coste Grahl, M.V.; Perin, A.P.A.; Lopes, F.C.; Porto, B.N.; Uberti, A.F.; Canavoso, L.E.; Stanisçuaski, F.; Fruttero, L.L. The role of extracellular nucleic acids in the immune system modulation of Rhodnius prolixus (Hemiptera: Reduviidae). Pestic. Biochem. Physiol. 2020, 167, 104591.
- Moyetta, N.R.; Broll, V.; Perin, A.P.A.; Uberti, A.F.; Coste Grahl, M.V.; Staniscuaski, F.; Carlini, C.R.; Fruttero, L.L. Jaburetox-induced toxic effects on the hemocytes of Rhodnius prolixus (Hemiptera: Reduviidae). Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 200, 17–26.
- Wu, Z.; Meyer-Hoffert, U.; Reithmayer, K.; Paus, R.; Hansmann, B.; He, Y.; Bartels, J.; Gläser, R.; Harder, J.; Schröder, J.M. Highly complex peptide aggregates of the S100 fused-type protein hornerin are present in human skin. J. Investig. Dermatol. 2009, 129, 1446–1458.
- Latendorf, T.; Gerstel, U.; Wu, Z.; Bartels, J.; Becker, A.; Tholey, A.; Schröder, J.M. Cationic Intrinsically Disordered Antimicrobial Peptides (CIDAMPs) Represent a New Paradigm of Innate Defense with a Potential for Novel Anti-Infectives. Sci. Rep. 2019, 9, 1–15.
- Gerstel, U.; Latendorf, T.; Bartels, J.; Becker, A.; Tholey, A.; Schröder, J.M. Hornerin contains a Linked Series of Ribosome-Targeting Peptide Antibiotics. Sci. Rep. 2018, 8, 1–15.
- Tyczewska, A.; Woźniak, E.; Gracz, J.; Kuczyński, J.; Twardowski, T. Towards Food Security: Current State and Future Prospects of Agrobiotechnology. Trends Biotechnol. 2018, 36, 1219–1229.
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–521.
- Didoné, D.A. Development of Jaburetox-expressing maize plants for resistance to pest lepidopterans. Ph.D. Thesis, Universidade de Passo Fundo, Passo Fundo, Brazil, 2018.
- Ceccon, C.C. Transgenic tobacco plants expressing hairpin and jaburetox RNA as strategies for Helicoverpa armigera control. Ph.D. Thesis, Universidade de Passo Fundo, Passo Fundo, Brazil, 2019.
- Rechenmacher, C.; Wiebke-Strohm, B.; de Oliveira-Busatto, L.A.; Weber, R.L.M.; Corso, M.C.M.; Lopes-Caitar, V.S.; Silva, S.M.H.; Dias, W.P.; Marcelino-Guimarães, F.C.; Carlini, C.R.; et al. Endogenous soybean peptide overexpression: An alternative to protect plants against root-knot nematodes. Biotechnol. Res. Innov. 2019, 3, 10–18.
- Biłas, R.; Szafran, K.; Hnatuszko-Konka, K.; Kononowicz, A.K. Cis-regulatory elements used to control gene expression in plants. Plant Cell. Tissue Organ Cult. 2016, 127, 269–287.
- Muthusamy, S.K.; Sivalingam, P.N.; Sridhar, J.; Singh, D.; Haldhar, S.M. Biotic stress inducible promoters in crop plants-a review. J. Agric. Ecol. 2017, 4, 14–24.

