 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Beatrix Zheng | -- | 1088 | 2022-11-14 01:44:23 | | | |
| 2 | Beatrix Zheng | + 14 word(s) | 1102 | 2022-11-14 03:28:20 | | | | |
| 3 | Beatrix Zheng | -2 word(s) | 1100 | 2022-11-15 06:55:35 | | |
Video Upload Options
Tyrocidine is a mixture of cyclic decapeptides produced by the bacteria Bacillus brevis found in soil. It can be composed of 4 different amino acid sequences, giving tyrocidine A–D. Tyrocidine is the major constituent of tyrothricin, which also contains gramicidin. Tyrocidine was the first commercially available antibiotic, but has been found to be toxic toward human blood and reproductive cells. The function of tyrocidine within its host B. brevis is thought to be regulation of sporulation. Tyrocidines A, B, and C are cyclic decapeptides. The biosynthesis of tyrocidine involves three enzymes. Parts of its sequence are identical to gramicidin S.
1. History
In 1939, the American microbiologist René Dubos discovered the soil microbe Bacillus brevis. He observed the ability of the microbe to decompose the capsule of pneumococcus bacterium, rendering it harmless. From the soil microbe B. brevis, he isolated tyrothricin, which had a high toxicity to a large range of bacteria. Tyrothricin was later found to be a mixture of the peptides gramicidin and tyrocidine. These were observed to have toxic effects in red blood cells and reproductive cells in humans, however, if applied externally as an ointment tyrocidine could also be used as a potent antimicrobial agent.[1] Dubos's discovery helped revive interest in research on penicillin.
2. Mechanism of Action
Tyrocidine has a unique mode of action in which it disrupts the cell membrane function, making it a favorable target for engineering derivatives.[2] Tyrocidine appears to perturb the lipid bilayer of a microbe’s inner membrane by permeating the lipid phase of the membrane. The exact affinity and location of tyrocidine within the phospholipid bilayer is not yet known.[3]
3. Biosynthesis
The biosynthesis of Tyrocidine is similar to Gramicidin S, and is achieved through the use of nonribosomal protein synthetases (NRPSs).[4] Its biosynthesis is via an enzymatic assembly consisting of 3 peptide synthetase proteins, TycA, TycB, and TycC, which contain 10 modules. The different tyrocidine analogues (A–D) are not produced by different enzymes, but rather by an enzyme system that is capable of incorporating different amino acids of structural similarity at specified sites. The amino acid sequence is determined by the organization of the enzyme and not by any RNA template.[5]

The tyrocine synthetases TycA, TycB, and TycC are encoded on the tyrocine operon. This consists of the three genes encoding for the three synthetases as well as three additional open reading frames (ORFs). These ORFs, labeled as TycD, TycE, and TycF are downstream of the three synthetase genes (see figure 1). TycD &TycE have the highest similarity to members of the ATP-binding cassette (ABC) transporter family which aid in the transport of substrates across a membrane. It has been suggested that the tandem transporters play a role in conferring resistance in the producer cell through tyrocidine secretion. TycF has been identified as a thioesterase (TE) and is similar to other TEs in bacterial operons used for encoding peptide synthetases. However, the precise function of these TEs remains unknown.[6] The size of the peptide synthetases corresponds to the amount of activation they carry out. TycA is the smallest and activates a single amino acid from one module, TycB is intermediate in size and activates 3 amino acids with 3 modules, and TycC is the largest and activates 6 amino acids with 6 modules (See figure 2).[6]
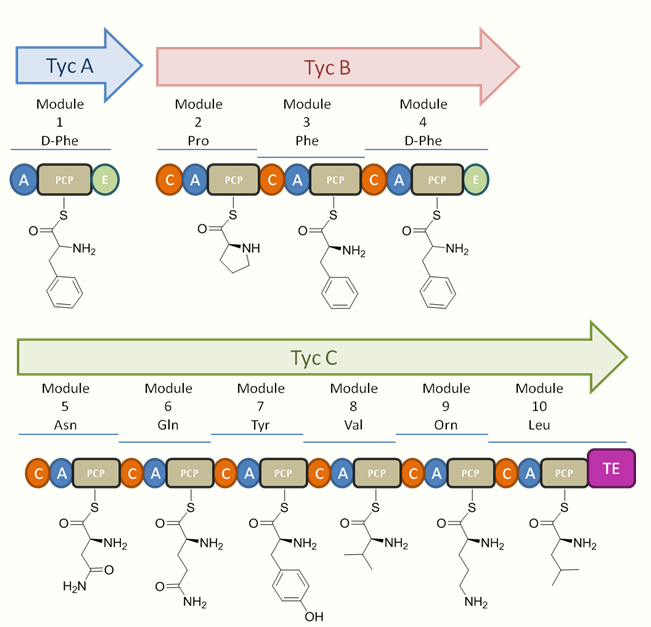
Each module performs all the catalytic reactions necessary to incorporate a single amino acid onto the peptide chain. This is accomplished through the subdomains for adenylation (A), peptityl carrier protein (PCP), condensation (C), and depending on the amino acid position, an epimerization (E). The adenylation subdomain is used in activating the specific amino acid. Each module uses one molecule of the selected substrate amino acid with one molecule of ATP to give an aminoacyl adenylate enzyme complex and pyrophosphate. The activated amino acid can then be transferred to the enzyme bound 4'-phosphopantetheine of the carrier protein with the expulsion of AMP from the system. The carrier protein uses the 4'-phosphopantetheine prosthetic group for loading of the growing peptide and their monomer precursors.[7] Elongation of the peptide chain is achieved through condensation of the upstream PCP onto an adjacent downstream PCP-bound monomer. In certain domains you will find modification subdomains, such as the E subdomain seen in domains 1 and 4 in tyrocidine, which will generate the D-configured amino acid. On the final module is the TE domain used as a catalyst for cyclization or product release. The release of the product from the carrier protein is achieved through acylation of the active site serine of TE in which the decapeptide is transferred from the thiol ether to the serine residue. Deacylation can then occur through intramolecular cyclization or through hydrolysis to give the cyclic or linear product respectively (See figure 3).
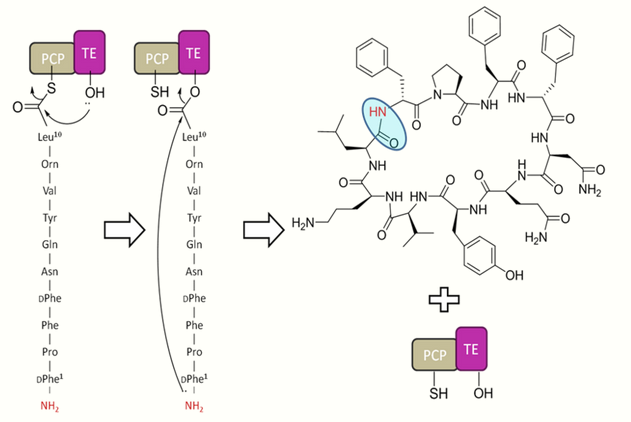
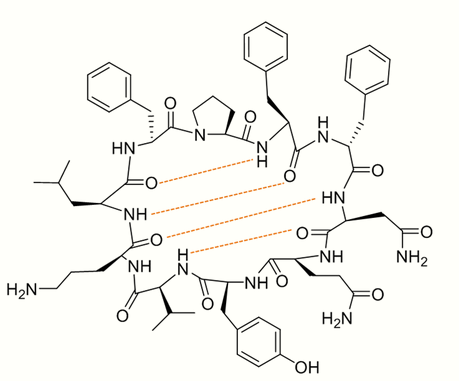
In the case of tyrocidine, ring closure has been shown to be highly favorable due to 4 H-bonds helping the decapeptide backbone to adopt a stable conformation (See figure 4).[2][7] This intramolecular cyclization occurs in a head-to-tail fashion involving the N-terminus of the D-Phe1 and the C-terminus of the L-Leu10 (See figure 3).[4][8]
4. Chemoenzymatic Strategies
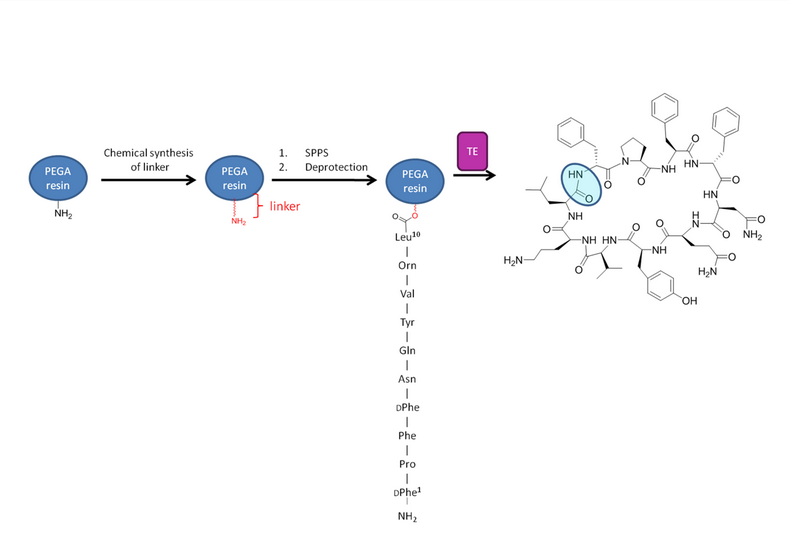
There is no general biochemical solution for macrocyclization of a peptide chain. Isolated tyrocidine (Tyc) TE domains can be used to cyclize chemically derived peptidyl-thioester substrates, providing a powerful route to new cyclic compounds. In order for this macrocyclization to occur, the peptide chain must be activated at its C-terminus with an N-acethylcysteamine (SNAC) leaving group.[4] An alanine scan through the 10 positions of tyrocidine shows that only the D-Phe and L-Orn are required for sufficient cyclization.
Tyc TE can also be used biomimetically in which it mimics the environment created by the TE domain with the substrate’s PCP through use of a synthetic tether linked to a polyethylene glycol (PEG) amide resin.[7] Use of this resin bound to a desired substrate with isolated TE can allow for catalytic release of the resin as well as macrocyclization of the substrate (See figure 5 [7]). Use of solid phase peptide synthesis (SPPS) allowed the incorporation of a diverse array of monomers into the peptide chain. Later studies used the high tolerance of Tyc TE in order to modify the peptide backbone post-synthetically. This also allowed for glycosylation of the tyrosine or serine residues to be incorporated.[4] Use of these methods has led to many promising new therapeutic agents.
References
- "Antibiotics". The Columbia Electronic Encyclopedia. 6th ed.. Columbia University Press. 2007. pp. online.
- "A chemical approach to generate molecular diversity based on the scaffold of cyclic decapeptide antibiotic tyrocidine A". J Comb Chem 5 (4): 353–5. 2003. doi:10.1021/cc0300255. PMID 12857101. https://dx.doi.org/10.1021%2Fcc0300255
- "The interaction of the antimicrobial peptide gramicidin S with lipid bilayer model and biological membranes". Biochim. Biophys. Acta 1462 (1–2): 201–21. December 1999. doi:10.1016/S0005-2736(99)00207-2. PMID 10590309. http://linkinghub.elsevier.com/retrieve/pii/S0005-2736(99)00207-2.
- "Macrocyclization strategies in polyketide and nonribosomal peptide biosynthesis". Nat Prod Rep 24 (4): 735–49. August 2007. doi:10.1039/b613652b. PMID 17653357. https://dx.doi.org/10.1039%2Fb613652b
- "Tyrocidine biosynthesis by three complementary fractions from Bacillus brevis (ATCC 8185)". Biochemistry 9 (25): 4839–45. December 1970. doi:10.1021/bi00827a002. PMID 4320358. https://dx.doi.org/10.1021%2Fbi00827a002
- "The tyrocidine biosynthesis operon of Bacillus brevis: complete nucleotide sequence and biochemical characterization of functional internal adenylation domains". J. Bacteriol. 179 (21): 6843–50. November 1997. PMID 9352938. PMC 179617. http://jb.asm.org/cgi/pmidlookup?view=long&pmid=9352938.
- "Biomimetic synthesis and optimization of cyclic peptide antibiotics". Nature 418 (6898): 658–61. August 2002. doi:10.1038/nature00907. PMID 12167866. https://dx.doi.org/10.1038%2Fnature00907
- "Peptide cyclization catalysed by the thioesterase domain of tyrocidine synthetase". Nature 407 (6801): 215–8. September 2000. doi:10.1038/35025116. PMID 11001063. https://dx.doi.org/10.1038%2F35025116

