 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Rita Xu | -- | 2497 | 2022-11-10 01:35:10 |
Video Upload Options
Embryogenesis is the process by which the embryo forms and develops. In mammals, the term refers chiefly to early stages of prenatal development, whereas the terms fetus and fetal development describe later stages. Embryogenesis starts with the fertilization of the egg cell (ovum) by a sperm cell, (spermatozoon). Once fertilized, the ovum is referred to as a zygote, a single diploid cell. The zygote undergoes mitotic divisions with no significant growth (a process known as cleavage) and cellular differentiation, leading to development of a multicellular embryo. Although embryogenesis occurs in both animal and plant development, this article addresses the common features among different animals, with some emphasis on the embryonic development of vertebrates and mammals.
1. Fertilization and the Zygote
The egg cell is generally asymmetric, having an "animal pole" (future ectoderm and mesoderm) and a "vegetal pole" (future endoderm). It is covered with protective envelopes, with different layers. The first envelope – the one in contact with the membrane of the egg – is made of glycoproteins and is known as the vitelline membrane (zona pellucida in mammals). Different taxa show different cellular and acellular envelopes englobing the vitelline membrane.
Fertilization (also known as 'conception', 'fecundation' and 'syngamy') is the fusion of gametes to produce a new organism. In animals, the process involves a sperm fusing with an ovum, which eventually leads to the development of an embryo. Depending on the animal species, the process can occur within the body of the female in internal fertilisation, or outside in the case of external fertilisation. The fertilized egg cell is known as the zygote.
To prevent more than one sperm fertilizing the egg, polyspermy, fast block and slow block to polyspermy are used. Fast block, the membrane potential rapidly depolarizing and then returning to normal, happens immediately after an egg is fertilized by a single sperm. Slow block begins the first few seconds after fertilization and is when the release of calcium causes the cortical reaction, various enzymes releasing from cortical granules in the eggs plasma membrane, to expand and harden the outside membrane, preventing more sperm from entering.[1]
2. Cleavage and Morula
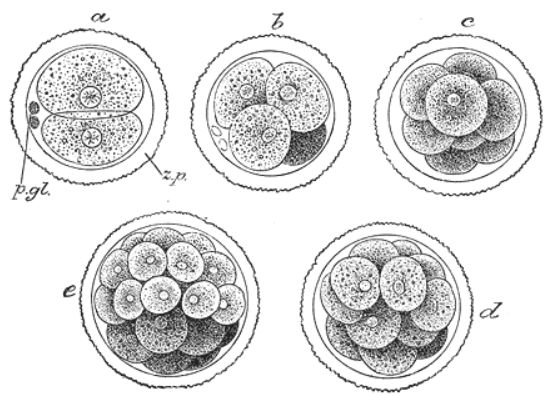
Cell division with no significant growth, producing a cluster of cells that is the same size as the original zygote, is called cleavage. At least four initial cell divisions occur, resulting in a dense ball of at least sixteen cells called the morula. The different cells derived from cleavage, up to the blastula stage, are called blastomeres. Depending mostly on the amount of yolk in the egg, the cleavage can be holoblastic (total) or meroblastic (partial).[2]
Holoblastic cleavage occurs in animals with little yolk in their eggs, such as humans and other mammals who receive nourishment as embryos from the mother, via the placenta or milk, such as might be secreted from a marsupium. On the other hand, meroblastic cleavage occurs in animals whose eggs have more yolk (i.e. birds and reptiles). Because cleavage is impeded in the vegetal pole, there is an uneven distribution and size of cells, being more numerous and smaller at the animal pole of the zygote.[2]
In holoblastic eggs the first cleavage always occurs along the vegetal-animal axis of the egg, and the second cleavage is perpendicular to the first. From here the spatial arrangement of blastomeres can follow various patterns, due to different planes of cleavage, in various organisms:
The end of cleavage is known as midblastula transition and coincides with the onset of zygotic transcription.
In amniotes, the cells of the morula are at first closely aggregated, but soon they become arranged into an outer or peripheral layer, the trophoblast, which does not contribute to the formation of the embryo proper, and an inner cell mass, from which the embryo is developed. Fluid collects between the trophoblast and the greater part of the inner cell-mass, and thus the morula is converted into a vesicle, called the blastodermic vesicle. The inner cell mass remains in contact, however, with the trophoblast at one pole of the ovum; this is named the embryonic pole, since it indicates the location where the future embryo will develop.[3]
3. Formation of the Blastula
After the 7th cleavage has produced 128 cells, the embryo is called a blastula.[2] The blastula is usually a spherical layer of cells (the blastoderm) surrounding a fluid-filled or yolk-filled cavity (the blastocoel)
Mammals at this stage form a structure called the blastocyst,[1] characterized by an inner cell mass that is distinct from the surrounding blastula. The blastocyst must not be confused with the blastula; even though they are similar in structure, their cells have different fates.
Before gastrulation, the cells of the trophoblast become differentiated into two strata: The outer stratum forms a syncytium (i.e., a layer of protoplasm studded with nuclei, but showing no evidence of subdivision into cells), termed the syncytiotrophoblast, while the inner layer, the cytotrophoblast or "Layer of Langhans", consists of well-defined cells. As already stated, the cells of the trophoblast do not contribute to the formation of the embryo proper; they form the ectoderm of the chorion and play an important part in the development of the placenta. On the deep surface of the inner cell mass, a layer of flattened cells, called the endoderm, is differentiated and quickly assumes the form of a small sac, called the yolk sac. Spaces appear between the remaining cells of the mass and, by the enlargement and coalescence of these spaces, a cavity called the amniotic cavity is gradually developed. The floor of this cavity is formed by the embryonic disk, which is composed of a layer of prismatic cells, the embryonic ectoderm, derived from the inner cell mass and lying in apposition with the endoderm.[3]
Formation of the Germ Layers
The embryonic disk becomes oval and then pear-shaped, the wider end being directed forward. Near the narrow, posterior end, an opaque streak, called the primitive streak, makes its appearance and extends along the middle of the disk for about one-half of its length; at the anterior end of the streak there is a knob-like thickening termed the primitive node or knot, (known as Hensen's knot in birds). A shallow groove, the primitive groove, appears on the surface of the streak, and the anterior end of this groove communicates by means of an aperture, the blastopore, with the yolk sac. The primitive streak is produced by a thickening of the axial part of the ectoderm, the cells of which multiply, grow downward, and blend with those of the subjacent endoderm. From the sides of the primitive streak a third layer of cells, the mesoderm, extends laterally between the ectoderm and endoderm; the caudal end of the primitive streak forms the cloacal membrane. The blastoderm now consists of three layers, named from without inward: ectoderm, mesoderm, and endoderm; each has distinctive characteristics and gives rise to certain tissues of the body. For many mammals, it is sometime during formation of the germ layers that implantation of the embryo in the uterus of the mother occurs.[3]
4. Formation of the Gastrula
During gastrulation cells migrate to the interior of the blastula, subsequently forming two (in diploblastic animals) or three (triploblastic) germ layers. The embryo during this process is called a gastrula. The germ layers are referred to as the ectoderm, mesoderm and endoderm. In diploblastic animals only the ectoderm and the endoderm are present.[2]
- Among different animals, different combinations of the following processes occur to place the cells in the interior of the embryo:
- Epiboly – expansion of one cell sheet over other cells[2]
- Ingression – migration of individual cells into the embryo (cells move with pseudopods)[3]
- Invagination – infolding of cell sheet into embryo, forming the mouth, anus, and archenteron.[2]
- Delamination – splitting or migration of one sheet into two sheets
- Involution – inturning of cell sheet over the basal surface of an outer layer
- Polar proliferation – Cells at the polar ends of the blastula/gastrula proliferate, mostly at the animal pole.
- Other major changes during gastrulation:
- Heavy RNA transcription using embryonic genes; up to this point the RNAs used were maternal (stored in the unfertilized egg).
- Cells start major differentiation processes, losing their totipotentiality.
In most animals, a blastopore is formed at the point where cells are entering the embryo. Two major groups of animals can be distinguished according to the blastopore's fate. In deuterostomes the anus forms from the blastopore, while in protostomes it develops into the mouth. See Embryological origins of the mouth and anus for more information.
4.1. Formation of the Early Nervous System – Neural Groove, Tube and Notochord
In front of the primitive streak, two longitudinal ridges, caused by a folding up of the ectoderm, make their appearance, one on either side of the middle line formed by the streak. These are named the neural folds; they commence some little distance behind the anterior end of the embryonic disk, where they are continuous with each other, and from there gradually extend backward, one on either side of the anterior end of the primitive streak. Between these folds is a shallow median groove, the neural groove. The groove gradually deepens as the neural folds become elevated, and ultimately the folds meet and coalesce in the middle line and convert the groove into a closed tube, the neural tube or canal, the ectodermal wall of which forms the rudiment of the nervous system. After the coalescence of the neural folds over the anterior end of the primitive streak, the blastopore no longer opens on the surface but into the closed canal of the neural tube, and thus a transitory communication, the neurenteric canal, is established between the neural tube and the primitive digestive tube. The coalescence of the neural folds occurs first in the region of the hind brain, and from there extends forward and backward; toward the end of the third week, the front opening (anterior neuropore) of the tube finally closes at the anterior end of the future brain, and forms a recess that is in contact, for a time, with the overlying ectoderm; the hinder part of the neural groove presents for a time a rhomboidal shape, and to this expanded portion the term sinus rhomboidalis has been applied. Before the neural groove is closed, a ridge of ectodermal cells appears along the prominent margin of each neural fold; this is termed the neural crest or ganglion ridge, and from it the spinal and cranial nerve ganglia and the ganglia of the sympathetic nervous system are developed.[4] By the upward growth of the mesoderm, the neural tube is ultimately separated from the overlying ectoderm.[5]
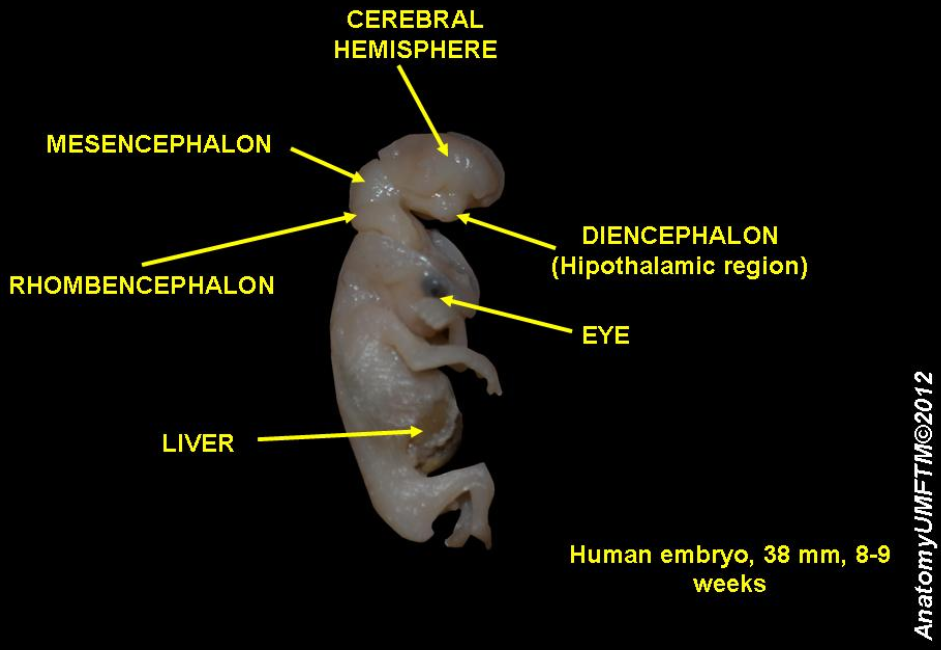
The cephalic end of the neural groove exhibits several dilatations that, when the tube is shut, assume the form of three vesicles; these constitute the three primary cerebral vesicles, and correspond, respectively, to the future 'fore-brain' (prosencephalon), 'midbrain' (mesencephalon), and 'hind-brain' (rhombencephalon) (Fig. 18). The walls of the vesicles are developed into the nervous tissue and neuroglia of the brain, and their cavities are modified to form its ventricles. The remainder of the tube forms the spinal cord (medulla spinalis); from its ectodermal wall the nervous and neuroglial elements of the spinal cord are developed, while the cavity persists as the central canal.[5]
4.2. Formation of the Early Septum
The extension of the mesoderm takes place throughout the whole of the embryonic and extra-embryonic areas of the ovum, except in certain regions. One of these is seen immediately in front of the neural tube. Here the mesoderm extends forward in the form of two crescentic masses, which meet in the middle line so as to enclose behind them an area that is devoid of mesoderm. Over this area, the ectoderm and endoderm come into direct contact with each other and constitute a thin membrane, the buccopharyngeal membrane, which forms a septum between the primitive mouth and pharynx.[3]
4.3. Early Formation of the Heart and Other Primitive Structures
In front of the buccopharyngeal area, where the lateral crescents of mesoderm fuse in the middle line, the pericardium is afterward developed, and this region is therefore designated the pericardial area. A second region where the mesoderm is absent, at least for a time, is that immediately in front of the pericardial area. This is termed the proamniotic area, and is the region where the proamnion is developed; in humans, however, it appears that a proamnion is never formed. A third region is at the hind end of the embryo, where the ectoderm and endoderm come into apposition and form the cloacal membrane.[3]
5. Somitogenesis
Somitogenesis is the process by which somites (primitive segments) are produced. These segmented tissue blocks differentiate into skeletal muscle, vertebrae, and dermis of all vertebrates.
Somitogenesis begins with the formation of somitomeres (whorls of concentric mesoderm) marking the future somites in the presomitic mesoderm (unsegmented paraxial). The presomitic mesoderm gives rise to successive pairs of somites, identical in appearance that differentiate into the same cell types but the structures formed by the cells vary depending upon the anteroposterior (e.g., the thoracic vertebrae have ribs, the lumbar vertebrae do not). Somites have unique positional values along this axis and it is thought that these are specified by the Hox homeotic genes.
Toward the end of the second week after fertilization, transverse segmentation of the paraxial mesoderm begins, and it is converted into a series of well-defined, more or less cubical masses, also known as the somites, which occupy the entire length of the trunk on either side of the middle line from the occipital region of the head. Each segment contains a central cavity (known as a myocoel), which, however, is soon filled with angular and spindle-shape cells. The somites lie immediately under the ectoderm on the lateral aspect of the neural tube and notochord, and are connected to the lateral mesoderm by the intermediate cell mass. Those of the trunk may be arranged in the following groups, viz.: cervical 8, thoracic 12, lumbar 5, sacral 5, and coccygeal from 5 to 8. Those of the occipital region of the head are usually described as being four in number. In mammals, somites of the head can be recognized only in the occipital region, but a study of the lower vertebrates leads to the belief that they are present also in the anterior part of the head and that, altogether, nine segments are represented in the cephalic region.[6]
6. Organogenesis
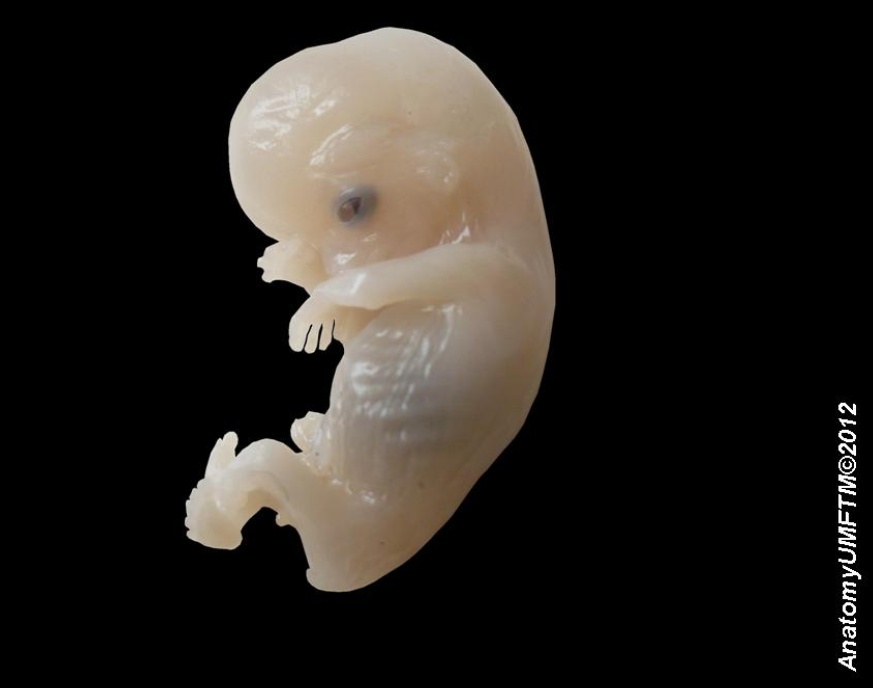
At some point after the different germ layers are defined, organogenesis begins. The first stage in vertebrates is called neurulation, where the neural plate folds forming the neural tube (see above).[2] Other common organs or structures that arise at this time include the heart and somites (also above), but from now on embryogenesis follows no common pattern among the different taxa of the animal kingdom.
In most animals organogenesis, along with morphogenesis, results in a larva. The hatching of the larva, which must then undergo metamorphosis, marks the end of embryonic development.
References
- Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002) (in en). Fertilization. Archived from the original on 2017-05-14. https://web.archive.org/web/20170514114426/https://www.ncbi.nlm.nih.gov/books/NBK26843/.
- What is a cell? 2004. A Science Primer: A Basic Introduction to the Science Underlying NCBI Resources. NCBI; and Campbell, Neil A.; Reece, Jane B.; Biology Benjamin Cummings, Pearson Education 2002. http://www.ncbi.nih.gov/About/primer/genetics_cell.html
- "Yahoo". Archived from the original on 2009-12-22. https://web.archive.org/web/20091222222223/http://education.yahoo.com/reference/gray/subjects/subject/6.
- E., Charles (2012-04-20) (in en). Embryogenesis. InTech. doi:10.5772/36871. ISBN 9789535104667. http://www.intechopen.com/books/embryogenesis/human-embryogenesis.
- "The Neural Groove and Tube". Archived from the original on 2007-08-22. https://web.archive.org/web/20070822135819/http://education.yahoo.com/reference/gray/subjects/subject/7.
- "The Primitive Segments". Archived from the original on 2007-09-11. https://web.archive.org/web/20070911054326/http://education.yahoo.com/reference/gray/subjects/subject/9.

