 +1 credit
+1 credit
| Version | Summary | Created by | Modification | Content Size | Created at | Operation |
|---|---|---|---|---|---|---|
| 1 | Pedro Sánchez Pellicer | -- | 5707 | 2022-11-03 11:55:16 | | | |
| 2 | Beatrix Zheng | + 3 word(s) | 5710 | 2022-11-04 04:22:31 | | | | |
| 3 | Beatrix Zheng | Meta information modification | 5710 | 2022-11-04 04:23:09 | | |
Video Upload Options
Alopecia areata is a multifactorial autoimmune-based disease with a complex pathogenesis. As in all autoimmune diseases, genetic predisposition is key. The collapse of the immune privilege of the hair follicle leading to scalp loss is a major pathogenic event in alopecia areata. The microbiota considered a bacterial ecosystem located in a specific area of the human body could somehow influence the pathogenesis of alopecia areata, as it occurs in other autoimmune diseases. Moreover, the Next Generation Sequencing of the 16S rRNA bacterial gene and the metagenomic methodology have provided an excellent characterization of the microbiota.
1. Hair Cycle Disruption
2. Hair Follicle Immune Privilege Collapse and Autoimmune Response
3. Role of Genetics in Alopecia Areata
4. Additional Influential Factors in the Development of Alopecia Areata
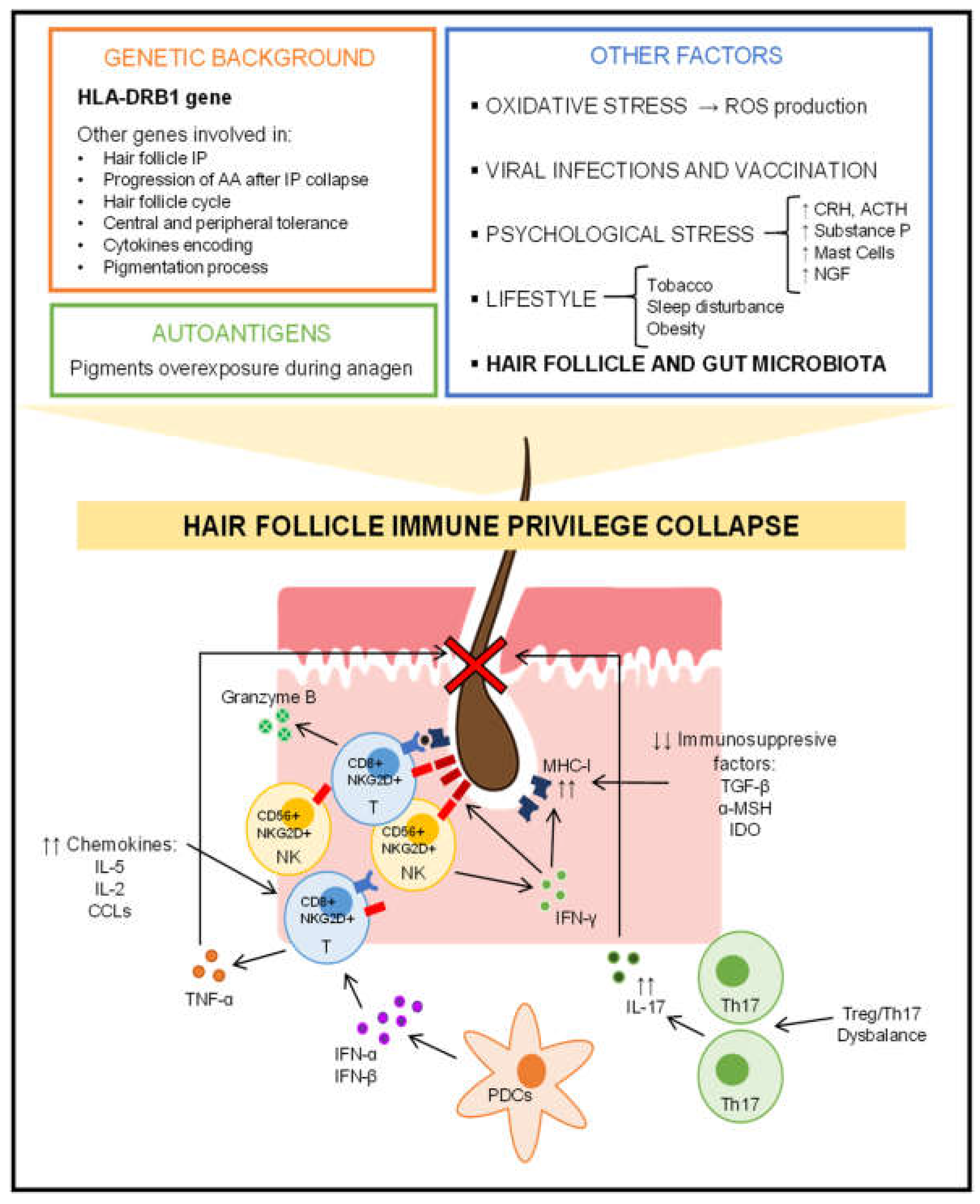
5. Alopecia Areata and Skin Microbiota
6. Alopecia Areata and Gut Microbiota
The human gastrointestinal tract harbors 100 trillion bacteria, most of which inhabit the large intestine, including about 1000 species [92]. These bacteria have cooperatively co-evolved with humans. This translates into a 1.3 to 1 ratio of bacteria to human cells, so there are more bacteria than human cells in human's organisms. The gut microbiota encodes more than 4 million genes (for approximately 25,000 genes contained in the human genome) and is involved in numerous metabolic reactions that influence the host’s physiology and metabolism to a substantial degree [93].
References
- Buffoli, B.; Rinaldi, F.; Labanca, M.; Sorbellini, E.; Trink, A.; Guanziroli, E.; Rezzani, R.; Rodella, L.F. The human hair: From anatomy to physiology. Int. J. Dermatol. 2014, 53, 331–341.
- Anzai, A.; Wang, E.H.C.; Lee, E.Y.; Aoki, V.; Christiano, A.M. Pathomechanisms of immune-mediated alopecia. Int. Immunol. 2019, 31, 439–447.
- Lin, X.; Zhu, L.; He, J. Morphogenesis, Growth Cycle and Molecular Regulation of Hair Follicles. Front. Cell. Dev. Biol. 2022, 10, 899095.
- Peckham, S.J.; Sloan, S.B.; Elston, D.M. Histologic features of alopecia areata other than peribulbar lymphocytic infiltrates. J. Am. Acad. Dermatol. 2011, 65, 615–620.
- Bernárdez, C.; Molina-Ruiz, A.M.; Requena, L. Histologic features of alopecias-part I: Nonscarring alopecias. Actas Dermosifiliogr. 2015, 106, 158–467.
- Watson, V.E.; Faniel, M.L.; Kamili, N.A.; Krueger, L.D.; Zhu, C. Immune-mediated alopecias and their mechanobiological aspects. Cells Dev. 2022, 170, 203793.
- Paus, R.; Ito, N.; Takigawa, M.; Ito, T. The hair follicle and immune privilege. J. Investig. Dermatol. Symp. Proc. 2003, 8, 188–194.
- Paus, R.; Bulfone-Paus, S.; Bertolini, M. Hair Follicle Immune Privilege Revisited: The Key to Alopecia Areata Management. J. Investig. Dermatol. Symp. Proc. 2018, 19, S12–S17.
- Suchonwanit, P.; Kositkuljorn, C.; Pomsoong, C. Alopecia Areata: An Autoimmune Disease of Multiple Players. Immunotargets Ther. 2021, 10, 299–312.
- Ito, T.; Ito, N.; Bettermann, A.; Tokura, Y.; Takigawa, M.; Paus, R. Collapse and restoration of MHC class-I-dependent immune privilege: Exploiting the human hair follicle as a model. Am. J. Pathol. 2004, 164, 623–634.
- Bertolini, M.; McElwee, K.; Gilhar, A.; Bulfone-Paus, S.; Paus, R. Hair follicle immune privilege and its collapse in alopecia areata. Exp. Dermatol. 2020, 29, 703–725.
- Rosenblum, M.D.; Yancey, K.B.; Olasz, E.B.; Truitt, R.L. CD200, a "no danger" signal for hair follicles. J. Dermatol. Sci. 2006, 41, 165–174.
- Kotwica-Mojzych, K.; Jodłowska-Jędrych, B.; Mojzych, M. CD200:CD200R Interactions and Their Importance in Immunoregulation. Int. J. Mol. Sci. 2021, 22, 1602.
- Trautman, S.; Thompson, M.; Roberts, J.; Thompson, C.T. Melanocytes: A possible autoimmune target in alopecia areata. J. Am. Acad. Dermatol. 2009, 61, 529–530.
- Bertolini, M.; Rossi, A.; Paus, R. Cover Image: Are melanocyte-associated peptides the elusive autoantigens in alopecia areata? Br. J. Dermatol. 2017, 176, 1106.
- Petukhova, L.; Duvic, M.; Hordinsky, M.; Norris, D.; Price, V.; Shimomura, Y.; Kim, H.; Singh, P.; Lee, A.; Chen, W.V.; et al. Genome-wide association study in alopecia areata implicates both innate and adaptive immunity. Nature 2010, 466, 113–117.
- Xing, L.; Dai, Z.; Jabbari, A.; Cerise, J.E.; Higgins, C.A.; Gong, W.; de Jong, A.; Harel, S.; DeStefano, G.M.; Rothman, L.; et al. Alopecia areata is driven by cytotoxic T lymphocytes and is reversed by JAK inhibition. Nat. Med. 2014, 20, 1043–1049.
- Simakou, T.; Butcher, J.P.; Reid, S.; Henriquez, F.L. Alopecia areata: A multifactorial autoimmune condition. J. Autoimmun. 2019, 98, 74–85.
- Ghoreishi, M.; Martinka, M.; Dutz, J.P. Type 1 interferon signature in the scalp lesions of alopecia areata. Br. J. Dermatol. 2010, 163, 57–62.
- Ito, T.; Ito, N.; Saatoff, M.; Hashizume, H.; Fukamizu, H.; Nickoloff, B.J.; Takigawa, M.; Paus, R. Maintenance of hair follicle immune privilege is linked to prevention of NK cell attack. J. Investig. Dermatol. 2008, 128, 1196–1206.
- Gilhar, A.; Keren, A.; Shemer, A.; d'Ovidio, R.; Ullmann, Y.; Paus, R. Autoimmune disease induction in a healthy human organ: A humanized mouse model of alopecia areata. J. Investig. Dermatol. 2013, 133, 844–847.
- Gilhar, A.; Kam, Y.; Assy, B.; Kalish, R.S. Alopecia areata induced in C3H/HeJ mice by interferon-gamma: Evidence for loss of immune privilege. J. Investig. Dermatol. 2005, 124, 288–289.
- Freyschmidt-Paul, P.; McElwee, K.J.; Hoffmann, R.; Sundberg, J.P.; Vitacolonna, M.; Kissling, S.; Zöller, M. Interferon-gamma-deficient mice are resistant to the development of alopecia areata. Br. J. Dermatol. 2006, 155, 515–521.
- Ito, T.; Suzuki, T.; Sakabe, J.I.; Funakoshi, A.; Fujiyama, T.; Tokura, Y. Plasmacytoid dendritic cells as a possible key player to initiate alopecia areata in the C3H/HeJ mouse. Allergol. Int. 2020, 69, 121–131.
- Saadeh, D.; Kurban, M.; Abbas, O. Update on the role of plasmacytoid dendritic cells in inflammatory/autoimmune skin diseases. Exp. Dermatol. 2016, 25, 415–421.
- Ito, T.; Tokura, Y. Alopecia areata triggered or exacerbated by swine flu virus infection. J. Dermatol. 2012, 39, 863–864.
- Tanemura, A.; Oiso, N.; Nakano, M.; Itoi, S.; Kawada, A.; Katayama, I. Alopecia areata: Infiltration of Th17 cells in the dermis, particularly around hair follicles. Dermatology 2013, 226, 333–336.
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730.
- Loh, S.H.; Moon, H.N.; Lew, B.L.; Sim, W.Y. Role of T helper 17 cells and T regulatory cells in alopecia areata: Comparison of lesion and serum cytokine between controls and patients. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1028–1033.
- Kasumagic-Halilovic, E.; Prohic, A.; Cavaljuga, S. Tumor necrosis factor-alpha in patients with alopecia areata. Indian. J. Dermatol. 2011, 56, 494–496.
- Abdel Halim, D.; Abu Zeid, O.M.; Rashed, L.; Saleh, M.A. Alteration of serum and tissue tumor necrosis factor alpha levels: A possible mechanism of action of oral pulse steroids in the treatment of alopecia areata. J. Cosmet. Dermatol. 2019, 18, 1128–1132.
- Bertolini, M.; Zilio, F.; Rossi, A.; Kleditzsch, P.; Emelianov, V.E.; Gilhar, A.; Keren, A.; Meyer, K.C.; Wang, E.; Funk, W.; et al. Abnormal interactions between perifollicular mast cells and CD8+ T-cells may contribute to the pathogenesis of alopecia areata. PLoS One 2014, 9, e94260.
- Blaumeiser, B.; van der Goot, I.; Fimmers, R.; Hanneken, S.; Ritzmann, S.; Seymons, K.; Betz, R.C.; Ruzicka, T.; Wienker, T.F.; De Weert, J.; et al. Familial aggregation of alopecia areata. J. Am. Acad. Dermatol. 2006, 54, 627–632.
- Petukhova, L.; Christiano, A.M. The genetic architecture of alopecia areata. J. Investig. Dermatol. Symp. Proc. 2013, 16, S16–S22.
- Ji, C.; Liu, S.; Zhu, K.; Luo, H.; Li, Q.; Zhang, Y.; Huang, S.; Chen, Q.; Cao, Y. HLA-DRB1 polymorphisms and alopecia areata disease risk: A systematic review and meta-analysis. Medicine 2018, 97, e11790.
- Arango, M.T.; Perricone, C.; Kivity, S.; Cipriano, E.; Ceccarelli, F.; Valesini, G.; Shoenfeld, Y. HLA-DRB1 the notorious gene in the mosaic of autoimmunity. Immunol. Res. 2017, 65, 82–98.
- Zuo, J.; Willcox, B.E.; Moss, P. ULBPs: Regulators of human lymphocyte stress recognition. Oncotarget 2017, 8, 106157–106158.
- Seok, H.; Jeon, H.S.; Park, H.J.; Kim, S.K.; Choi, J.H.; Lew, B.L.; Chung, J.H.; Sim, W.Y. Association of HSPA1B SNP rs6457452 with Alopecia Areata in the Korean population. Immunol. Investig. 2014, 43, 212–223.
- Betz, R.C.; Petukhova, L.; Ripke, S.; Huang, H.; Menelaou, A.; Redler, S.; Becker, T.; Heilmann, S.; Yamany, T.; Duvic, M.; et al. Genome-wide meta-analysis in alopecia areata resolves HLA associations and reveals two new susceptibility loci. Nat. Commun. 2015, 6, 5966.
- Rajabi, F.; Abdollahimajd, F.; Jabalameli, N.; Nassiri Kashani, M.; Firooz, A. The Immunogenetics of Alopecia areata. Adv. Exp. Med. Biol. 2022, 1367, 19–59.
- Wengraf, D.A.; McDonagh, A.J.; Lovewell, T.R.; Vasilopoulos, Y.; Macdonald-Hull, S.P.; Cork, M.J.; Messenger, A.G.; Tazi-Ahnini, R. Genetic analysis of autoimmune regulator haplotypes in alopecia areata. Tissue. Antigens. 2008, 71, 206–212.
- Conteduca, G.; Rossi, A.; Megiorni, F.; Parodi, A.; Ferrera, F.; Tardito, S.; Battaglia, F.; Kalli, F.; Negrini, S.; Pizzuti, A.; et al. Single nucleotide polymorphisms in the promoter regions of Foxp3 and ICOSLG genes are associated with Alopecia areata. Clin. Exp. Med. 2014, 14, 91–97.
- Lei, Z.X.; Chen, W.J.; Liang, J.Q.; Wang, Y.J.; Jin, L.; Xu, C.; Kang, X.J. The association between rs2476601 polymorphism in PTPN22 gene and risk of alopecia areata: A meta-analysis of case-control studies. Medicine 2019, 98, e15448.
- Kim, H.K.; Lee, H.; Lew, B.L.; Sim, W.Y.; Kim, Y.O.; Lee, S.W.; Lee, S.; Cho, I.K.; Kwon, J.T.; Kim, H.J. Association between TAP1 gene polymorphisms and alopecia areata in a Korean population. Genet Mol. Res. 2015, 14, 18820–18827.
- Seleit, I.; Bakry, O.A.; Gayed, E.A.E.; Gawad, A.E.D. Polymorphism of FAS and FAS Ligand Genes in Alopecia Areata: A Case-control Study in Egyptian Population. Indian J. Dermatol. 2018, 63, 220–226.
- AlFadhli, S.; Nanda, A. Genetic evidence for the involvement of NOTCH4 in rheumatoid arthritis and alopecia areata. Immunol. Lett. 2013, 150, 130–133.
- Kim, S.K.; Chung, J.H.; Park, H.J.; Kang, S.W.; Lim, D.J.; Byun, S.H.; Baek, D.G.; Ko, H.Y.; Lew, B.L.; Baik, H.H.; et al. Polymorphisms in the promoter regions of the CXCL1 and CXCL2 genes contribute to increased risk of alopecia areata in the Korean population. Genet Mol. Res. 2015, 14, 9667–9674.
- Fischer, J.; Degenhardt, F.; Hofmann, A.; Redler, S.; Basmanav, F.B.; Heilmann-Heimbach, S.; Hanneken, S.; Giehl, K.A.; Wolff, H.; Moebus, S.; et al. Genomewide analysis of copy number variants in alopecia areata in a Central European cohort reveals association with MCHR2. Exp. Dermatol. 2017, 26, 536–541.
- Petukhova, L.; Patel, A.V.; Rigo, R.K.; Bian, L.; Verbitsky, M.; Sanna-Cherchi, S.; Erjavec, S.O.; Abdelaziz, A.R.; Cerise, J.E.; Jabbari, A.; et al. Integrative analysis of rare copy number variants and gene expression data in alopecia areata implicates an aetiological role for autophagy. Exp. Dermatol. 2020, 29, 243–253.
- Abdel Fattah, N.S.; Ebrahim, A.A.; El Okda, E.S. Lipid peroxidation/antioxidant activity in patients with alopecia areata. J. Eur. Acad. Dermatol. Venereol. 2011, 25, 403–408.
- Akar, A.; Arca, E.; Erbil, H.; Akay, C.; Sayal, A.; Gür, A.R. Antioxidant enzymes and lipid peroxidation in the scalp of patients with alopecia areata. J. Dermatol. Sci. 2002, 29, 85–90.
- Acharya, P.; Mathur, M.C. Oxidative stress in alopecia areata: A systematic review and meta-analysis. Int. J. Dermatol. 2020, 59, 434–440.
- Sachdeva, S.; Khurana, A.; Goyal, P.; Sardana, K. Does oxidative stress correlate with disease activity and severity in alopecia areata? An analytical study. J. Cosmet. Dermatol. 2022, 21, 1629–1634.
- Fokam, D.; Hoskin, D. Instrumental role for reactive oxygen species in the inflammatory response. Front. Biosci. (Landmark Ed) 2020, 25, 1110–1119.
- Jain, U.; Saxena, K.; Chauhan, N. Helicobacter pylori induced reactive oxygen Species: A new and developing platform for detection. Helicobacter 2021, 26, e12796.
- Paus, R.; Arck, P. Neuroendocrine perspectives in alopecia areata: Does stress play a role? J. Investig. Dermatol. 2009, 129, 1324–1326.
- Ito, N.; Ito, T.; Kromminga, A.; Bettermann, A.; Takigawa, M.; Kees, F.; Straub, R.H.; Paus, R. Human hair follicles display a functional equivalent of the hypothalamic-pituitary-adrenal axis and synthesize cortisol. FASEB J. 2005, 19, 1332–1334.
- Arck, P.C.; Handjiski, B.; Peters, E.M.; Peter, A.S.; Hagen, E.; Fischer, A.; Klapp, B.F.; Paus, R. Stress inhibits hair growth in mice by induction of premature catagen development and deleterious perifollicular inflammatory events via neuropeptide substance P-dependent pathways. Am. J. Pathol. 2003, 162, 803–814.
- Azzawi, S.; Penzi, L.R.; Senna, M.M. Immune Privilege Collapse and Alopecia Development: Is Stress a Factor. Skin Appendage. Disord. 2018, 4, 236–244.
- Ito, N.; Sugawara, K.; Bodó, E.; Takigawa, M.; van Beek, N.; Ito, T.; Paus, R. Corticotropin-releasing hormone stimulates the in situ generation of mast cells from precursors in the human hair follicle mesenchyme. J. Investig. Dermatol. 2010, 130, 995–1004.
- Alexopoulos, A.; Chrousos, G.P. Stress-related skin disorders. Rev. Endocr. Metab. Disord. 2016, 17, 295–304.
- Smatti, M.K.; Cyprian, F.S.; Nasrallah, G.K.; Al Thani, A.A.; Almishal, R.O.; Yassine, H.M. Viruses and Autoimmunity: A Review on the Potential Interaction and Molecular Mechanisms. Viruses 2019, 11, 762.
- Fukuyama, M.; Ito, T.; Ohyama, M. Alopecia areata: Current understanding of the pathophysiology and update on therapeutic approaches, featuring the Japanese Dermatological Association guidelines. J. Dermatol. 2022, 49, 19–36.
- Nguyen, B.; Tosti, A. Alopecia in patients with COVID-19: A systematic review and meta-analysis. JAAD Int. 2022, 7, 67–77.
- May Lee, M.; Bertolani, M.; Pierobon, E.; Lotti, T.; Feliciani, C.; Satolli, F. Alopecia areata following COVID-19 vaccination: Vaccine-induced autoimmunity? Int. J. Dermatol. 2022, 61, 634–635.
- Minokawa, Y.; Sawada, Y.; Nakamura, M. Lifestyle Factors Involved in the Pathogenesis of Alopecia Areata. Int. J. Mol. Sci. 2022, 23, 1038.
- Dai, Y.X.; Yeh, F.Y.; Shen, Y.J.; Tai, Y.H.; Chou, Y.J.; Chang, Y.T.; Chen, T.J.; Li, C.P.; Wu, C.Y. Cigarette Smoking, Alcohol Consumption, and Risk of Alopecia Areata: A Population-Based Cohort Study in Taiwan. Am. J. Clin. Dermatol. 2020, 21, 901–911.
- Melnik, B.C.; John, S.M.; Chen, W.; Plewig, G. T helper 17 cell/regulatory T-cell imbalance in hidradenitis suppurativa/acne inversa: The link to hair follicle dissection, obesity, smoking and autoimmune comorbidities. Br. J. Dermatol. 2018, 179, 260–272.
- Dai, Y.X.; Tai, Y.H.; Chen, C.C.; Chang, Y.T.; Chen, T.J.; Chen, M.H. Bidirectional association between alopecia areata and sleep disorders: A population-based cohort study in Taiwan. Sleep Med. 2020, 75, 112–116.
- Inui, S.; Hamasaki, T.; Itami, S. Sleep quality in patients with alopecia areata: Questionnaire-based study. Int. J. Dermatol. 2014, 53, e39–e41.
- Besedovsky, L.; Lange, T.; Haack, M. The Sleep-Immune Crosstalk in Health and Disease. Physiol. Rev. 2019, 99, 1325–1380.
- Hagino, T.; Okazaki, S.; Serizawa, N.; Suzuki, K.; Kaga, M.; Otsuka, Y.; Mikami, E.; Hoashi, T.; Saeki, H.; Matsuda, H.; et al. Dietary Habits in Japanese Patients with Alopecia Areata. Clin. Cosmet. Investig. Dermatol. 2021, 14, 1579–1591.
- Nakamizo, S.; Honda, T.; Adachi, A.; Nagatake, T.; Kunisawa, J.; Kitoh, A.; Otsuka, A.; Dainichi, T.; Nomura, T.; Ginhoux, F.; et al. High fat diet exacerbates murine psoriatic dermatitis by increasing the number of IL-17-producing γδ T cells. Sci. Rep. 2017, 7, 14076.
- Taylor, E.B. The complex role of adipokines in obesity, inflammation, and autoimmunity. Clin. Sci. 2021, 135, 731–752.
- Nakatsuji, T.; Chiang, H.I.; Jiang, S.B.; Nagarajan, H.; Zengler, K.; Gallo, R.L. The microbiome extends to subepidermal compartments of normal skin. Nat. Commun. 2013, 4, 431.
- Watanabe, K.; Nishi, E.; Tashiro, Y.; Sakai, K. Mode and Structure of the Bacterial Community on Human Scalp Hair. Microbes. Environ. 2019, 34, 252–259.
- Matard, B.; Meylheuc, T.; Briandet, R.; Casin, I.; Assouly, P.; Cavelier-balloy, B.; Reygagne, P. First evidence of bacterial biofilms in the anaerobe part of scalp hair follicles: A pilot comparative study in folliculitis decalvans. J. Eur. Acad. Dermatol. Venereol. 2013, 27, 853–860.
- Lousada, M.B.; Lachnit, T.; Edelkamp, J.; Rouillé, T.; Ajdic, D.; Uchida, Y.; Di Nardo, A.; Bosch, T.C.G.; Paus, R. Exploring the human hair follicle microbiome. Br. J. Dermatol. 2021, 184, 802–815.
- Polak-Witka, K.; Rudnicka, L.; Blume-Peytavi, U.; Vogt, A. The role of the microbiome in scalp hair follicle biology and disease. Exp. Dermatol. 2020, 29, 286–294.
- Byrd, A.L.; Belkaid, Y.; Segre, J.A. The human skin microbiome. Nat. Rev. Microbiol. 2018, 16, 143–155.
- Constantinou, A.; Kanti, V.; Polak-Witka, K.; Blume-Peytavi, U.; Spyrou, G.M.; Vogt, A. The Potential Relevance of the Microbiome to Hair Physiology and Regeneration: The Emerging Role of Metagenomics. Biomedicines 2021, 9, 236.
- Reithmayer, K.; Meyer, K.C.; Kleditzsch, P.; Tiede, S.; Uppalapati, S.K.; Gläser, R.; Harder, J.; Schröder, J.M.; Paus, R. Human hair follicle epithelium has an antimicrobial defence system that includes the inducible antimicrobial peptide psoriasin (S100A7) and RNase 7. Br. J. Dermatol. 2009, 161, 78–89.
- Chen, X.; Niyonsaba, F.; Ushio, H.; Okuda, D.; Nagaoka, I.; Ikeda, S.; Okumura, K.; Ogawa, H. Synergistic effect of antibacterial agents human beta-defensins, cathelicidin LL-37 and lysozyme against Staphylococcus aureus and Escherichia coli. J. Dermatol. Sci. 2005, 40, 123–132.
- Wang, Z.; Mascarenhas, N.; Eckmann, L.; Miyamoto, Y.; Sun, X.; Kawakami, T.; Di Nardo, A. Skin microbiome promotes mast cell maturation by triggering stem cell factor production in keratinocytes. J. Allergy Clin. Immunol. 2017, 139, 1205–1216.e6.
- Sobiepanek, A.; Kuryk, Ł.; Garofalo, M.; Kumar, S.; Baran, J.; Musolf, P.; Siebenhaar, F.; Fluhr, J.W.; Kobiela, T.; Plasenzotti, R.; et al. The Multifaceted Roles of Mast Cells in Immune Homeostasis, Infections and Cancers. Int. J. Mol. Sci. 2022, 23, 2249.
- Kashem, S.W.; Haniffa, M.; Kaplan, D.H. Antigen-Presenting Cells in the Skin. Annu. Rev. Immunol. 2017, 35, 469–499.
- Hall, J.B.; Cong, Z.; Imamura-Kawasawa, Y.; Kidd, B.A.; Dudley, J.T.; Thiboutot, D.M.; Nelson, A.M. Isolation and Identification of the Follicular Microbiome: Implications for Acne Research. J. Investig. Dermatol. 2018, 138, 2033–2040.
- Lai, Y.; Di Nardo, A.; Nakatsuji, T.; Leichtle, A.; Yang, Y.; Cogen, A.L.; Wu, Z.R.; Hooper, L.V.; Schmidt, R.R.; von Aulock, S.; et al. Commensal bacteria regulate Toll-like receptor 3-dependent inflammation after skin injury. Nat. Med. 2009, 15, 1377–1382.
- Mohan, G.C.; Silverberg, J.I. Association of Vitiligo and Alopecia Areata With Atopic Dermatitis: A Systematic Review and Meta-analysis. JAMA Dermatol. 2015, 151, 522–528.
- Edslev, S.M.; Agner, T.; Andersen, P.S. Skin Microbiome in Atopic Dermatitis. Acta Derm. Venereol. 2020, 100, adv00164.
- Malik, K.; Guttman-Yassky, E. Cytokine Targeted Therapeutics for Alopecia Areata: Lessons from Atopic Dermatitis and Other Inflammatory Skin Diseases. J. Investig. Dermatol. Symp. Proc. 2018, 19, S62–S64.
- Gill, S.R.; Pop, M.; Deboy, R.T.; Eckburg, P.B.; Turnbaugh, P.J.; Samuel, B.S.; Gordon, J.I.; Relman, D.A.; Fraser-Liggett, C.M.; Nelson, K.E. Metagenomic analysis of the human distal gut microbiome. Science 2006, 312, 1355–1359.
- Gilbert, J.A.; Blaser, M.J.; Caporaso, J.G.; Jansson, J.K.; Lynch, S.V.; Knight, R. Current understanding of the human microbiome. Nat. Med. 2018, 24, 392–400.
- Rothschild, D.; Weissbrod, O.; Barkan, E.; Kurilshikov, A.; Korem, T.; Zeevi, D.; Costea, P.I.; Godneva, A.; Kalka, I.N.; Bar, N.; et al. Environment dominates over host genetics in shaping human gut microbiota. Nature 2018, 555, 210–215.
- Zmora, N.; Suez, J.; Elinav, E. You are what you eat: Diet, health and the gut microbiota. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 35–56.
- Lozupone, C.A.; Stombaugh, J.I.; Gordon, J.I.; Jansson, J.K.; Knight, R. Diversity, stability and resilience of the human gut microbiota. Nature 2012, 489, 220–230.
- Shanahan, F.; Ghosh, T.S.; O'Toole, P.W. The Healthy Microbiome-What Is the Definition of a Healthy Gut Microbiome? Gastroenterology 2021, 160, 483–494.
- Moya, A.; Ferrer, M. Functional Redundancy-Induced Stability of Gut Microbiota Subjected to Disturbance. Trends Microbiol. 2016, 24, 402–413.
- Takiishi, T.; Fenero, C.I.M.; Câmara, N.O.S. Intestinal barrier and gut microbiota: Shaping our immune responses throughout life. Tissue Barriers 2017, 5, e1373208.
- Jiao, Y.; Wu, L.; Huntington, N.D.; Zhang, X. Crosstalk Between Gut Microbiota and Innate Immunity and Its Implication in Autoimmune Diseases. Front. Immunol. 2020, 11, 282.
- Belkaid, Y.; Naik, S. Compartmentalized and systemic control of tissue immunity by commensals. Nat. Immunol. 2013, 14, 646–653.
- Sinha, S.; Lin, G.; Ferenczi, K. The skin microbiome and the gut-skin axis. Clin. Dermatol. 2021, 39, 829–839.
- Fu, Y.; Lee, C.H.; Chi, C.C. Association of Psoriasis With Inflammatory Bowel Disease: A Systematic Review and Meta-analysis. JAMA Dermatol. 2018, 154, 1417–1423.
- Ramírez-Boscá, A.; Navarro-López, V.; Martínez-Andrés, A.; Such, J.; Francés, R.; Horga de la Parte, J.; Asín-Llorca, M. Identification of Bacterial DNA in the Peripheral Blood of Patients With Active Psoriasis. JAMA Dermatol. 2015, 151, 670–671.
- Miyauchi, E.; Shimokawa, C.; Steimle, A.; Desai, M.S.; Ohno, H. The impact of the gut microbiome on extra-intestinal autoimmune diseases. Nat. Rev. Immunol. 2022. online ahead of print.
- Kinashi, Y.; Hase, K. Partners in Leaky Gut Syndrome: Intestinal Dysbiosis and Autoimmunity. Front. Immunol. 2021, 12, 673708.
- Clemente, J.C.; Manasson, J.; Scher, J.U. The role of the gut microbiome in systemic inflammatory disease. BMJ 2018, 360, j5145.
- Borde, A.; Åstrand, A. Alopecia areata and the gut-the link opens up for novel therapeutic interventions. Expert. Opin. Ther. Targets 2018, 22, 503–511.
- Lee, S.; Lee, H.; Lee, C.H.; Lee, W.S. Comorbidities in alopecia areata: A systematic review and meta-analysis. J. Am. Acad. Dermatol. 2019, 80, 466–477.e16.
- Gilhar, A.; Schrum, A.G.; Etzioni, A.; Waldmann, H.; Paus, R. Alopecia areata: Animal models illuminate autoimmune pathogenesis and novel immunotherapeutic strategies. Autoimmun. Rev. 2016, 15, 726–735.
- Sundberg, J.P.; Elson, C.O.; Bedigian, H.; Birkenmeier, E.H. Spontaneous, heritable colitis in a new substrain of C3H/HeJ mice. Gastroenterology 1994, 107, 1726–1735.
- Maghfour, J.; Olson, J.; Conic, R.R.Z.; Mesinkovska, N.A. The Association between Alopecia and Inflammatory Bowel Disease: A Systematic Review and Meta-Analysis. Dermatology 2021, 237, 658–672.
- Glassner, K.L.; Abraham, B.P.; Quigley, E.M.M. The microbiome and inflammatory bowel disease. J. Allergy Clin. Immunol. 2020, 145, 16–27.
- McElwee, K.J.; Niiyama, S.; Freyschmidt-Paul, P.; Wenzel, E.; Kissling, S.; Sundberg, J.P.; Hoffmann, R. Dietary soy oil content and soy-derived phytoestrogen genistein increase resistance to alopecia areata onset in C3H/HeJ mice. Exp. Dermatol. 2003, 12, 30–36.
- Nair, L.; Dai, Z.; Christiano, A.M. Gut microbiota plays a role in the development of alopecia areata. J. Investig. Dermatol. 2017, 137, S112.
- Raugh, A.; Allard, D.; Bettini, M. Nature vs. nurture: FOXP3, genetics, and tissue environment shape Treg function. Front. Immunol. 2022, 13, 911151.
- Sanchez Rodriguez, R.; Pauli, M.L.; Neuhaus, I.M.; Yu, S.S.; Arron, S.T.; Harris, H.W.; Yang, S.H.; Anthony, B.A.; Sverdrup, F.M.; Krow-Lucal, E.; et al. Memory regulatory T cells reside in human skin. J. Clin. Invest. 2014, 124, 1027–1036.
- Scharschmidt, T.C.; Vasquez, K.S.; Pauli, M.L.; Leitner, E.G.; Chu, K.; Truong, H.A.; Lowe, M.M.; Sanchez Rodriguez, R.; Ali, N.; Laszik, Z.G.; et al. Commensal Microbes and Hair Follicle Morphogenesis Coordinately Drive Treg Migration into Neonatal Skin. Cell Host Microbe 2017, 21, 467–477.e5.
- McElwee, K.J.; Freyschmidt-Paul, P.; Hoffmann, R.; Kissling, S.; Hummel, S.; Vitacolonna, M.; Zöller, M. Transfer of CD8(+) cells induces localized hair loss whereas CD4(+)/CD25(-) cells promote systemic alopecia areata and CD4(+)/CD25(+) cells blockade disease onset in the C3H/HeJ mouse model. J. Investig. Dermatol. 2005, 124, 947–957.
- Ali, N.; Zirak, B.; Rodriguez, R.S.; Pauli, M.L.; Truong, H.A.; Lai, K.; Ahn, R.; Corbin, K.; Lowe, M.M.; Scharschmidt, T.C. Regulatory T Cells in Skin Facilitate Epithelial Stem Cell Differentiation. Cell 2017, 169, 1119–1129.e11.
- Hamed, F.N.; Åstrand, A.; Bertolini, M.; Rossi, A.; Maleki-Dizaji, A.; Messenger, A.G.; McDonagh, A.J.G.; Tazi-Ahnini, R. Alopecia areata patients show deficiency of FOXP3+CD39+ T regulatory cells and clonotypic restriction of Treg TCRβ-chain, which highlights the immunopathological aspect of the disease. PLoS One 2019, 14, e0210308.
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From Dietary Fiber to Host Physiology: Short-Chain Fatty Acids as Key Bacterial Metabolites. Cell 2016, 165, 1332–1345.
- Smith, P.M.; Howitt, M.R.; Panikov, N.; Michaud, M.; Gallini, C.A.; Bohlooly-Y, M.; Glickman, J.N.; Garrett, W.S. The microbial metabolites, short-chain fatty acids, regulate colonic Treg cell homeostasis. Science 2013, 341, 569–573.
- Mu, Q.; Kirby, J.; Reilly, C.M.; Luo, X.M. Leaky Gut As a Danger Signal for Autoimmune Diseases. Front. Immunol. 2017, 8, 598.
- Hacınecipoğlu, F.; Gönül, M.; Özdemir, Ş.; Demir, Ö.F. Is there a link between alopecia areata and gut? J. Cosmet. Dermatol. 2022. online ahead of print.
- Rebello, D.; Wang, E.; Yen, E.; Lio, P.A.; Kelly, C.R. Hair Growth in Two Alopecia Patients after Fecal Microbiota Transplant. ACG Case Rep. J. 2017, 4, e107.
- Xie, W.R.; Yang, X.Y.; Xia, H.H.; Wu, L.H.; He, X.X. Hair regrowth following fecal microbiota transplantation in an elderly patient with alopecia areata: A case report and review of the literature. World J. Clin. Cases 2019, 7, 3074–3081.

